一个新疆杂草黑麦居群的细胞学分析
2016-11-30车永和申雅婷杨燕萍李旭青徐荣蕊刘伟华李秀全杨欣明
车永和,申雅婷,杨燕萍,李旭青,宋 楠,徐荣蕊,刘伟华,李秀全,杨欣明
(1.河北科技师范学院生命科技学院,河北秦皇岛 066600;2.中国农业科学院作物科学研究所,北京 100081)
一个新疆杂草黑麦居群的细胞学分析
车永和1,申雅婷1,杨燕萍1,李旭青1,宋 楠1,徐荣蕊1,刘伟华2,李秀全2,杨欣明2
(1.河北科技师范学院生命科技学院,河北秦皇岛 066600;2.中国农业科学院作物科学研究所,北京 100081)
新疆杂草黑麦(Secalecerealesubsp.segetaleZhuk.)是我国发现的一种稀有野生黑麦资源,具有极强的抗病、抗逆性和优异的丰产性状。为系统了解该特殊种质资源的生物学特征,并为这一珍贵种质资源应用于远缘杂交育种、牧草种质开发及其生物多样性保护提供基础的细胞学依据,选取新疆杂草黑麦的一个居群89R38,观察分析其花粉母细胞的减数分裂行为规律及其根尖细胞的核型特点。结果表明,89R38的绝大多数花粉母细胞减数分裂中染色体的行为正常,在终变期同源染色体配对可形成7个二价体,而在少数花粉母细胞减数分裂中(2.06%)观察到落后染色体、染色体桥等异常行为;成熟花粉粒的醋酸洋红染色反应检测其花粉粒的育性为99.08%;核型公式为2n=2x=14=12m+2sm,核型为1A型。
新疆杂草黑麦;花粉母细胞;减数分裂;染色体行为;核型分析
黑麦属(SecaleL.)植物是最早也是最成功地用于小麦改良的近缘植物之一,具有许多普通小麦所不及的优良性状,另外作为粮食和饲料作物在全球也有较为广泛的种植。由于自然或人为因素破坏,一些野生型、杂草型黑麦属物种目前已经逐渐消失,这无疑是对宝贵基因资源的巨大浪费。我国发现的一种稀有野生黑麦类型,初步命名为Secalecerealesubsp.segetaleZhuk.,其突出优点是抗白粉病和锈病,具有很强的抗旱、抗寒能力,另一个突出优点是分蘖力强,有的材料有效分蘖平均达30个[1-3]。具有这些特性的此类黑麦对饲用黑麦和旱地小麦育种,尤其针对目前生产中突出的小麦抗病、抗旱、抗寒和丰产遗传改良将会起到积极作用。目前此类遗传资源更多地是被埋藏在种质资源库中,除个别研究报道外[4-7],尚缺少系统评价和研究。
细胞学准确直观体现物种染色体特征特性,为物种研究提供基本生物学特征。植物的染色体数目与核型分析是对染色体的各种特征进行定性和定量描述的一种基本方法。减数分裂时期的染色体配对行为能准确直观反应染色体结构变异。与单纯的形态分类相比,染色体数据可以解决常规形态分类难以解决的问题[8-9]。本试验对新疆杂草黑麦中选取的一个居群的减数分裂时期染色体行为、成熟花粉粒育性及其染色体核型进行了观察分析,以期为这一珍贵种质资源应用于远缘杂交育种、牧草种质开发及其生物多样性保护提供基础的细胞学依据。
1 材料与方法
供试材料为中国农业科学院作物科学研究所收集的新疆杂草黑麦居群,现种植和保存于河北科技师范学院。随机选取该类种质中的一个居群89R38,于4月下旬或5月初,待黑麦的旗叶露出2~3 cm时,在每天早上的8:00-9:00对黑麦幼穗进行采集。花粉母细胞固定和根尖染色体制备参照代 明等[7]的方法,在卡诺液中固定26 h,转入70%乙醇中于4 ℃保存备用。采用细胞压片法制备染色体标本,并在显微镜下观察拍照。核型类型参照Stebbins[10]的标准。核型不对称系数(As.K,%)根据Arano[11]的方法进行计算,具体计算公式为:
长臂总长/全组染色体总长×100%。
2 结果与分析
2.1 花粉母细胞减数分裂观察结果
对随机选取的黑麦居群89R38的200个减数分裂时期的花粉母细胞进行了观察,结果(图1)发现,97.94%的花粉母细胞减数分裂染色体的行为正常,于染色体计数的最佳时期中期I出现了7个环状二价体或6个环状二价体和1个棒状二价体2种情况,但以7个环状二价体为绝大多数,占95%以上;2.06%的花粉母细胞减数分裂行为出现异常,于减数分裂过程中出现染色体桥、落后染色体及异常四分体。
2.2 花粉粒育性的观察结果
通过随机取样的方法,共观察统计了1 200个成熟花粉粒的醋酸洋红染色反应结果(图2),其中不被染色或染色浅的、外形皱瘪的花粉粒有11个,可育花粉粒占99.08%,花粉败育率为0.92%。
2.3 体细胞核型分析结果
通过对50个新疆杂草黑麦居群89R38根尖细胞中期染色体数目的统计,未发现染色体数目变异,确定其染色体数目为14(图3)。核型分析参数取自5个分散良好、着丝点清晰的中期细胞染色体的平均值,新疆杂草黑麦居群89R38的第7对同源染色体的臂比值在1.71~3.00范围内,染色体为近中部着丝点染色体(sm型),其余6对同源染色体的臂比值都在1.00~1.70范围内,染色体为中部着丝点染色体(m型),在居群89R38的染色体中没有发现具随体的染色体。根据以上数据,可以确定新疆杂草黑麦居群89R38的染色体核型公式为2n=2x=14=12m+2sm。
由表1可知,居群89R38染色体相对长度范围为13.27%~18.11%,根据Stebbins[10]提出的“不对称核型分析”分类标准,居群89R38染色体中最长和最短的染色体的比值为1.36,没有发现臂比大于2∶1的染色体,属于1A型。根据Arano[10]的计算方法得出,居群89R38染色体的不对称系数为56.10%,对称程度较高,是较为对称类型。
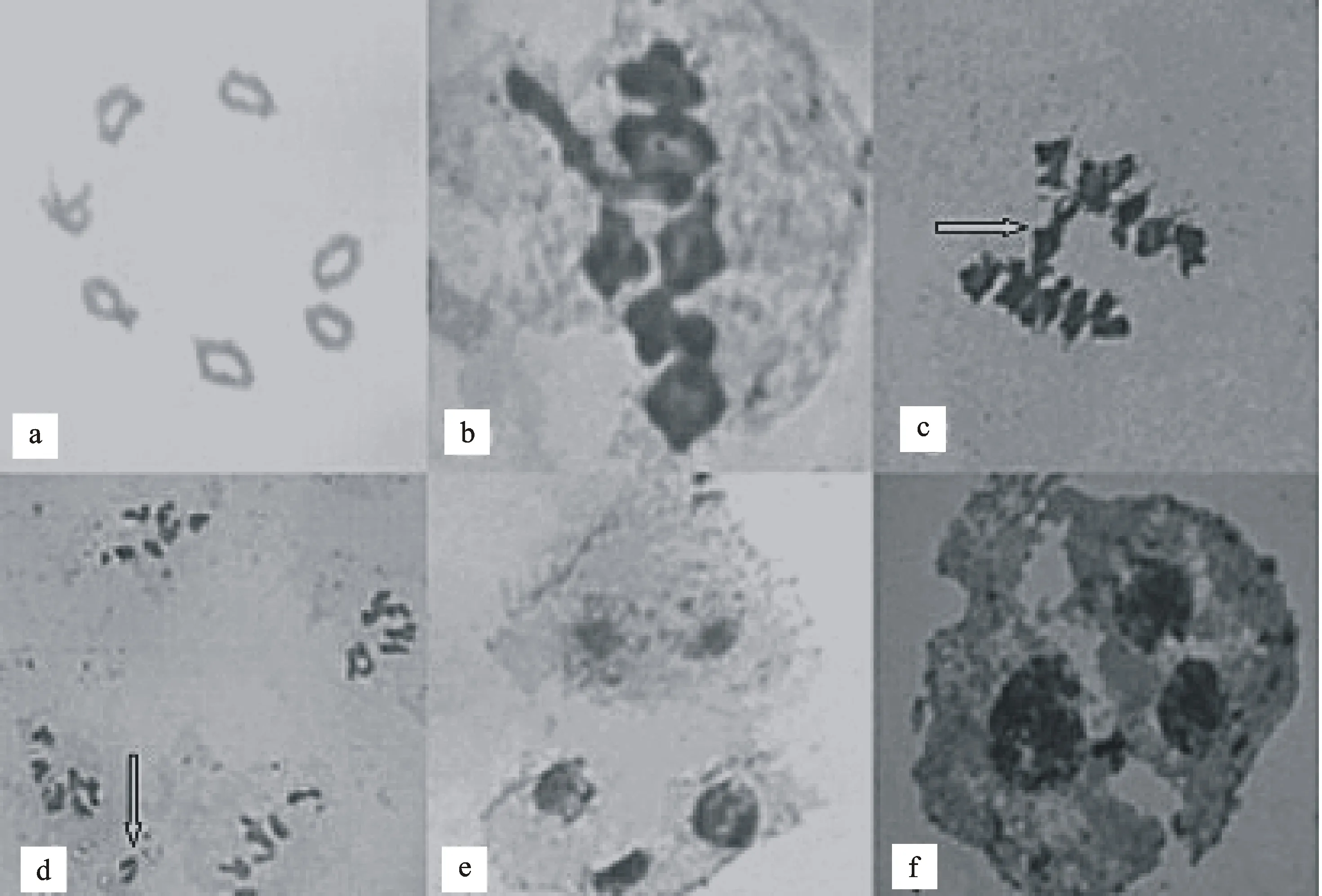
a:中期具7个环状二价体;b:中期具6个环状二价体和1个棒状二价体;c:后期Ⅰ染色体桥(箭头所示);d:后期Ⅱ落后染色体(箭头所示);e和f:异常四分体。
a:Seven ringⅡ in metaphase; b:Six ring Ⅱ+ 1 rodⅡ in metaphase; c:Chromosome bridge in anaphase I indicated by arrow; d:Laggard chromosomes in anaphase Ⅱ indicated by arrow; e and f: Abnormal tetrad.
图1 新疆杂草黑麦居群89R38花粉母细胞减数分裂
Fig.1 Meiosis of pollen mother cells inS.cerealssubsp.segetalepopulation 89R38

箭头所示败育花粉粒。 The arrows stand for infertile pollen.

核型公式Karyotypeformula染色体相对长度范围Chromosomerelativelengthrange/%最长染色体与最短染色体长度之比Lengthratioofthelongestchromosometotheshortestone臂比>2的染色体数No.ofchromosomewitharmratio(L/S)>2臂指数Armindex核型不对称系数As.K/%核型类型Karyotypetype2n=2x=14=12m+2sm13.27~18.111.3600.4456.101A
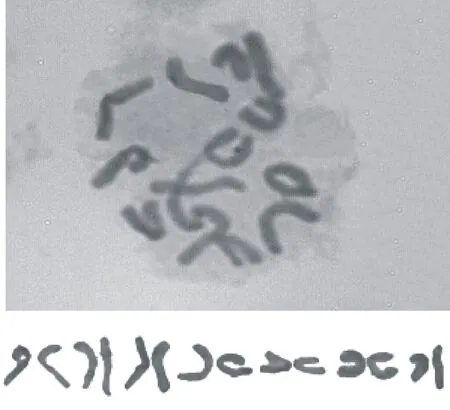
图3 新疆杂草黑麦居群89R38根尖细胞染色体核型分析
3 讨 论
本研究发现新疆杂草黑麦89R38的花粉母细胞减数分裂过程中有少量的(2.06%)花粉母细胞减数分裂行为异常,主要表现为染色体落后和染色体桥等。成熟花粉粒的醋酸洋红染色反应检测花粉粒育性为99.08%。落后染色体和染色体桥造成染色体的不均匀分配,形成遗传组成不平衡的配子,产生不同遗传组成的花粉粒,这是导致花粉败育的重要原因之一。Ricci等[12]对俯仰臂形草[Brachiariadecumbens(Poaceae)]进行研究,发现二倍体材料减数分裂异常率从0.82%到7.93%不等,四倍体材料减数分裂异常率从18.41%到65.83%不等。Silva等[13]如同本研究所观察到的非正常四分体也在Mesosetumchaseae(Poaceae)的减数分裂过程中观察到形成三个细胞的异常四分体。Felismino等[14]在六倍体和四倍体Urochloaadspersa(Trin.) R. D. Webster物种中分别观察到5.3%和 6.2%的非正常细胞,而在另外一个种中由于落后染色体、染色体粘连等导致了16.9%非正常细胞。异常减数分裂会形成不平衡配子影响花粉活力,Pagliarini等[15]在珊状臂形草[Brachiariabrizantha(Poaceae)]中观察到异常的减数分裂从0.36%至95.76%不等,所有材料中只有四倍体异常减数分裂细胞低于40%而适合用于种内和种间杂交。在进化上稳定的植物通过花粉败育淘汰染色体变异的配子,有利于维持物种的稳定性。新疆杂草黑麦居群89R38少数花粉细胞在减数分裂过程中虽有染色体落后、染色体桥等异常现象出现导致一定量的的花粉败育,但整体花粉可育性高。
代 明等[7]对新疆杂草黑麦的核型研究中检测到1A和2A两种类型的核型,而本研究中新疆杂草黑麦居群89R38的核型为1A型,且两研究染色体数据特征间存在一定差异。同样,杨欣明等[3]和姚景侠等[9]的研究结论也表明新疆杂草黑麦居群间染色体形态上存在差异。这些研究表明,新疆杂草黑麦在染色体水平具有丰富的遗传多样性。染色体作为遗传物质的主要载体,其结构或数量上的任何变化都会引起相应的遗传改变,从而更加丰富物种遗传多样性。本研究结果检测和验证了新疆杂草黑麦细胞学特征,对黑麦属基因资源的搜集、保存、研究和利用提供了相关依据。
[1]董玉琛,杨欣明,周荣华,等.黑麦的种类及遗传研究进展[J].新疆农业科学,1994(5):205.
DONG Y S,YANG X M,ZHOU R H,etal.Research Progress on the species and inheritance of rye [J].XinjiangAgricultureSciences,1994(5):205.
[2]陆 峻,戚家华,杨松杰.新疆杂草黑麦的分布、分类地位及来源[J].新疆农业科学,1994(1):24.
LU J,QI J H,YANG S J.Distribution,taxonomic status and origin of weedy rye in Xinjiang [J].XinjiangAgricultureSciences,1994(1):24.
[3]杨欣明,董玉琛,周荣华,等.新疆杂草黑麦抗病性鉴定和细胞学研究[J].新疆农业科学,1994(3):117.
YANG X M,DONG Y S,ZHOU R H,etal.Resistance identification and cytology analysis of weedy rye in Xinjiang [J].XinjiangAgricultureSciences,1994(3):117.
[4]车永和,杨欣明,杨燕萍,等.新疆杂草黑麦居群遗传多样性研究[J].麦类作物学报,2008,28(5):755.
CHE Y H,YANG X M,YANG Y P,etal.Genetic diversity ofSecalecerealesubsp.segetalepopulations in Xinjiang [J].JournalofTriticeaeCrops,2008,28(5):755.
[5]车永和,杨欣明,杨燕萍,等.新疆杂草黑麦居群形态学性状调查[J].湖北农业科学,2008,47(8):880.
CHE Y H,YANG X M,YANG Y P,etal.Morphological investigation ofSecalecerealesubsp.segetalepopulations in Xinjiang [J].HubeiAgriculturalSciences,2008,47(8):880.
[6]CHE Y H,DAI M,YANG Y P,etal.Genetic diversity of gliadin inSecalecerealesubsp.segetalefrom Xinjiang,China [J].GeneticResourcesandCropEvolution,2016,63(7):1173.
[7]代 明,李 飞,陈 嫚,等.新疆杂草黑麦染色体核型分析[J].麦类作物学报,2013,33(3):440.
DAI M,LI F,CHEN M,etal.Karyotypes analysis ofSecalecerealesubsp.segetale[J].JournalofTriticeaeCrops,2013,33(3):440.
[8]李懋学,陈瑞阳.关于植物核型分析的标准化问题[J].武汉植物学研究,1985,3(4):297.
LI C X,CHEN R Y.A suggestion on the standardization of kyrotype analysis in plant [J].JournalofWuhanBotanicalResearch,1985,3(4):297.
[9]姚景侠,程本旭. “冰麦”的核型分析和C-带带型分析[J].作物品种资源,1985(4):19.
YAO J X,CHENG B X.Karyotype and C-band pattern analysis of 'ice wheat' [J].ChinaSeeds,1985(4):19.
[10]STEBBINS G L.Chromosomal Evolution in Higher Plants [M].London: Edward Aronld,1971:88.
[11]ARANO H.Cytological studies in subfamily carduoideae (compositae) of Japan Ⅸ [J].BotanicalMagazine,1963,76:32.
[12]RICCI G C,De Souza-Kaneshima A M,Felismino M F,etal.Chromosome numbers and meiotic analysis in the pre-breeding ofBrachiariadecumbens(Poaceae) [J].JournalofGenetetic,2011,90(2):289.
[13]SILVA L A C,PAGLIARINI M S,SANTOS S A,etal.Chromosome number,microsporogenesis,microgametogenesis,and pollen viability in the Brazilian native grassMesosetumchaseae(Poaceae) [J].GeneticsandMolecularResearch,2012,11(4): 4100.
[14]FELISMINO M F,MAIOR R L S,DAMASCENO JR G A,etal.Meiotic behavior and chromosome number ofUrochloaadspersa(Trin.) R.D.Webster from the Brazilian Chaco [J].GeneticsandMolecularResearch,2015,14(3):7455.
[15]PAGLIARINI M S,VALLE C B,SANTOS E M,etal.Microsporogenesis inBrachiariabrizantha(Poaceae) as a selection tool for breeding [J].GeneticsandMolecularResearch,2012,11(2):1309.
Cytological Studies of a Population ofSecalecerealesubsp.segetaleZhuk.
CHE Yonghe1, SHEN Yating1, YANG Yanping1, LI Xuqing1, SONG Nan1,XU Rongrui1, LIU Weihua2, LI Xiuquan2, YANG Xinming2
(1.School of Life Science and Technology, Hebei College of Normal and Sci-Tech, Qinghuangdao, Hebei 066600,China; 2.Institute of Crop Sciences, Chinese Academy of Agriculture Science, Beijing 100081, China)
A weedy rye was found in Xinjiang province of China. The studies described its classification as a subspecies ofSecalecerealeL. and namedSecalecerealesubsp.segetaleZhuk. Strong stress tolerance and disease resistance, and the desirable genes exhibiting good yield-related traits ofS.cerealesubsp.segetalesuggested it could be a good germplasm resource for wheat improvement and forage breeding. The cytological studies ofS.cerealesubsp.segetalepopulation 89R38 were conducted on the root tip karyotype and the pollen mother cell meiosis behavior for better understanding this special species. The observation results showed that chromosomes of most pollen mother cells were paired as seven bivalents. The laggard chromosomes, chromosome bridge and other abnormal behavior were observed in a minority of meiosis of pollen mother cells, taking up to 2.06%. The percentage of pollen fertility was calculated as 99.08% by stained with aceto-carmine. The karyotype formula of population 89R38 was 2n=2x=14=12m+2sm, as a 1A karyotype.
S.cerealesubsp.segetale; Pollen mother cells; Chromosome behaviour; Karyotypes analysis
时间:2016-10-08
2016-04-13
2016-08-24
河北省自然科学基金项目(C2015407056);河北科技师范学院科学研究基金项目
E-mail:cheyonghe2002@163.com(车永和);yangxinming@caas.cn(杨欣明,与第一作者同等贡献)
S512.5;S326
A
1009-1041(2016)10-1323-05
网络出版地址:http://www.cnki.net/kcms/detail/61.1359.S.20161008.0932.016.html
