喀斯特地区同一花序白三叶形态多样性研究
2016-11-29吴永洁王元素
吴永洁,王元素,李 莉
(1.贵州师范大学生命科学学院,中国 贵阳 550001;2.贵州省草原监理站,中国 贵阳 550001)
喀斯特地区同一花序白三叶形态多样性研究
吴永洁1,王元素2,李 莉1*
(1.贵州师范大学生命科学学院,中国 贵阳 550001;2.贵州省草原监理站,中国 贵阳 550001)
通过观测19份野生白三叶形态指标,比较同一花序及不同海拔高度的贵州野生白三叶形态多样性.结果表明:同一花序及不同海拔高度的贵州野生白三叶形态都存在较大差异;各指标总变异系数从大到小为中叶面积、地上生物量、匍匐茎长度、生长点数、株高、中叶长、中叶宽、生长习性、有无“V”斑;不同海拔区域白三叶形态形状居群间存在不同程度的差异.进行聚类统计显示分类与海拔高度没有明显的关系.
喀斯特;贵州;白三叶;同一花序;形态多样性
基因漂移、遗传漂变、遗传瓶颈、自然选择和地方适应性等因素,可以通过在不同时空尺度上的动态变化来影响并改变生物适应性和多样化的进程[1].目前,花粉污染、基因漂移等问题已经在实验中得到普遍证实,而大部分研究只是针对一些基础性工作,如花粉传播因素、基因漂移距离、杂交亲和性等[2-5].空间分离群体之间的隔离和基因流动的程度决定了遗传差异、生殖隔离和物种形成的潜在能力.遗传漂变和自然选择都会使群体之间的差异增加,而基因流的作用会使群体之间的差异减小[5].研究同一花序水平上植物形态多样性,可以减少以上因素的影响,使其表型性状与其生态适应性的研究更趋于一致.
白三叶(TrifoliumrepensL.)是多年生草本优质牧草,属异化授粉植物,广泛分布于世界各地,在农牧业中地位重要.贵州是中国发育最典型的喀斯特地貌之一,不仅地势起伏大、切割强、相对高度常达到300~700 m,且喀斯特广泛发育,地貌类型较复杂.其次,贵州具有高原峡谷地貌结构特征,导致水土资源分布不平衡,形成了多种特殊的生态位.白三叶在长期的演化过程中,不断向着适应其生境的方向进化.本研究收集贵州省境内19份白三叶种质为试验材料,比较其不同海拔高度居群间和同一花序水平上的多样性,为下一步选育优质高效白三叶新品种提供指导.
1 材料和方法
1.1 贵州地理与气候
试验在贵州贵阳市贵州牧草推广站温室大棚内进行,种子收集于贵州省不同海拔区域.贵州全省大部分地区气候温和,四季分明.全省平均气温的最高值出现在7月,平均22~25 ℃;最低值出现在1月,平均为4~6 ℃;年平均气温为14~19 ℃.全年极端最高气温为34.0~36.0 ℃,极端最低气温为-6.0~-9.0 ℃.常年雨量充沛,时空分布不均.全省各地多年平均年降水量大部分地区为1 100~1 300 mm,最大值接近1 600 mm,最小值约为850 mm.一年中的大多数雨量集中在夏季.全省大部分地区年日照1 200~1 600 h,气候特点是垂直方向差异较大,立体气候明显.
1.2 材料收集
2013年5~8月在全省范围内采集野生白三叶种子,有效样本19份,见表1.
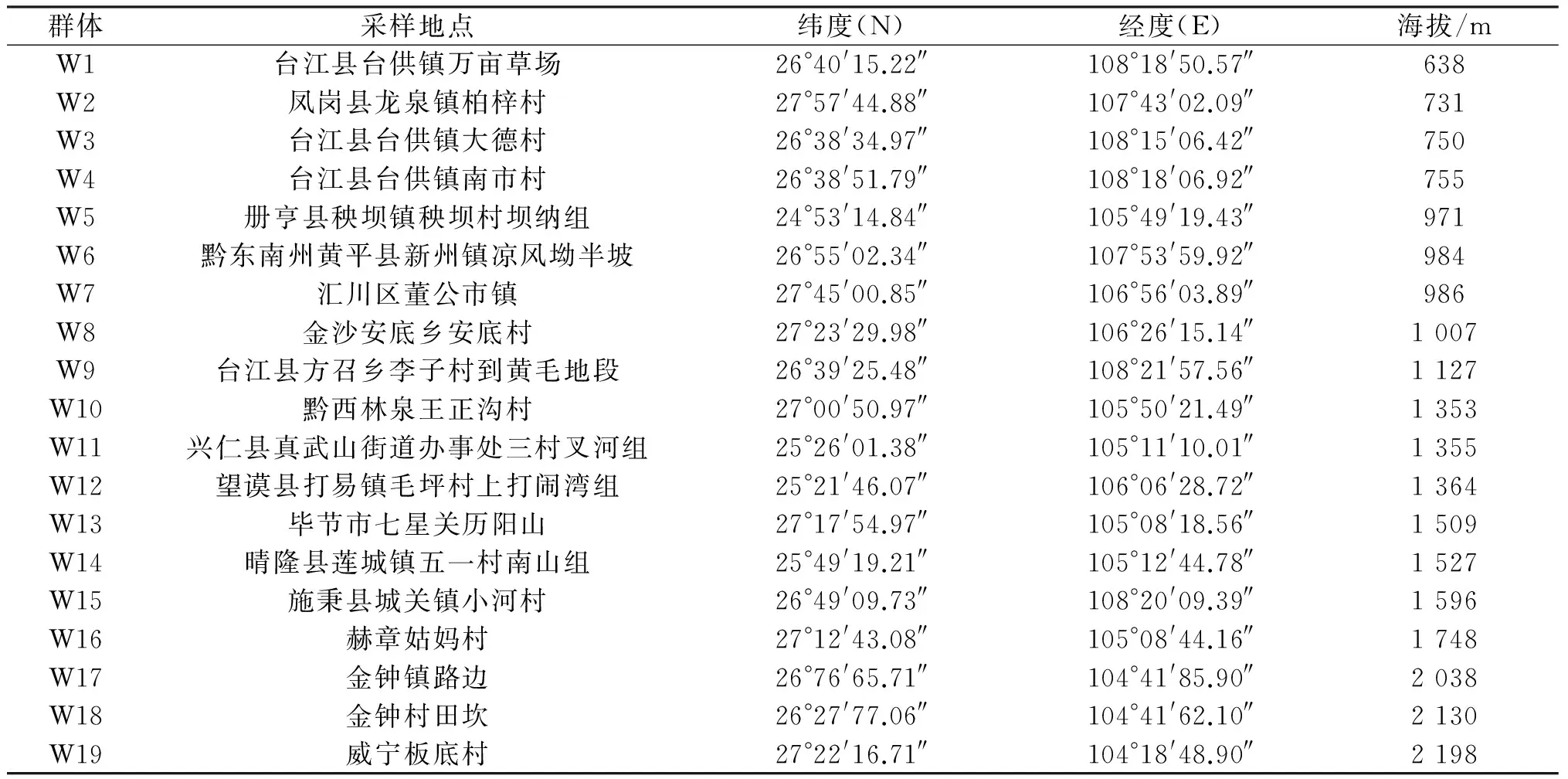
表1 试验材料
1.3 栽培与观测
1.3.1 温室栽培 2014年9月20日在贵州省牧草推广站温室大棚播种.从19份材料中各选取一个果实饱满的花序,将来自同一花序的种子播入有32个小格的育苗培养盘,孔穴大小60 mm×60 mm.每小格播2粒种子,覆土1 cm,浇水.培养基质为m(石英砂)∶m(细土)∶m(腐殖质)=3∶4∶3混合而成,高压灭菌冷却后装盘.待长出小苗后留选长势较好的一株苗用于观察研究.
1.3.2 施肥和管理 每隔3个月施复合肥1次,每次施用量0.03 kg/m2,苗期施尿素1次,使用量折合0.012 kg/m2,及时除杂草,适时浇水.
1.3.3 测量指标 2015年5月1日,参照《牧草种质资源描述规范和数据标准》[6]及前人研究方法观测V型斑纹、叶面积、株高、中叶宽、中叶长、匍匐茎长度、生长点个数及地上生物量等指标.为便于分析,将非量化性状特征进行标准化赋值.海拔:1=1 000 m以下,2=1 000~1 500 m,3=1 500~2 000 m,4=2 000 m以上;有无V型斑:有斑=1,无斑=2;生长习性:1=直立生长,2=匍匐分枝生长,3=匍匐不分枝生长.
1.4 数据分析
用Excel和SPSS 16.0对试验数据进行统计,分析不同海拔高度区域白三叶种内、同一花序水平上各个性状的差异程度.各形态变异性以变异系数CV表示,CV(%)=100×S/M(S为标准差,M为平均值).用Person相关系数分析各个性状之间的相关性,并对各个指标标准化后用算术平均法(UPGMA)聚类分析.
2 结果与分析
2.1 白三叶形态学分析
2.1.1 同一花序白三叶形态学性状差异分析 同一花序水平上和不同种群间白三叶形态存在较大差异(表2).总变异系数大于300%的有7个居群,其中W16总变异系数最大,高达429.74%,其次是W12,W1,W9,W6,W3,W10;最小的是W7,总变异系数为236.9%.各指标总变异系数从大到小为中叶面积、地上生物量、匍匐茎长度、生长点数、株高、中叶长、中叶宽、生长习性、有无“V”斑.株高变异系数最大的是W10和W18,分别为37.93%和37.51%,W2最小,只有12.63%.地上生物量变异系数最大的是W3和W1,分别为98.48%和81.67%,W14和W7的最小,为38.31%和38.55%.匍匐茎长度的变异范围为30.95%~65.75%,变异系数最大和最小分别是W7和W3.中叶宽的变异范围主要为10.89%~32.11%,只有居群W8的变异系数高达60.61%.中叶长的变异范围为13.33%~32.77%,变异系数最大的为W16,最小为W7.中叶面积的总变异系数比其他指标大,其中W16,W12,W3和W6的变异系数均大于100%,最小的是W11,只有30.33%.生长习性的变异范围为0~35.33%,其中W14,W18和W19的变异系数均为0,即这几个居群都是匍匐分枝生长.有无“V”形斑中只有W1,W5,W11,W13和W19发生了变异,其余居群全都有“V”斑.生长点个数最大变异是W1,为64.09%,其次是W16,W9,W12和W10,最小的是W2,仅为0.75%.
2.1.2 不同海拔高度白三叶形态学数量性状分析 不同海拔区域白三叶形态形状居群间存在显著差异,见表3.总体上,随着海拔的提升,白三叶植株越来越高,生长点数越来越多,地上生物量也越来越大;而匍匐茎长度、中叶宽、中叶长和中叶面积则随着海拔的提升而增大,高于2 000 m的区域则有所减小.在低于1 000 m的区域,匍匐茎长度的变异系数最大,高达102.87%,其次是地上生物量、中叶面积,最小的是中叶宽,仅为25.49%.在1 000~1 500 m区域,变异范围为21.43%~80.46%,最大的为匍匐茎长度,其次是地上生物量、生长点个数、中叶面积,最小的是中叶宽.在1 500~2 000 m区域,变异范围是21.29%~49.62%,其中变异最大的为地上生物量,其次是中叶面积,中叶宽变异最小.在大于2 000 m的区域,匍匐茎长度的变异系数最大,为61.37%,其次是地上生物量、中叶面积、株高,最小为中叶长,仅为23.21%.
2.2 形态学形状相关性分析
白三叶的株高与地上生物量、匍匐茎长度、中叶宽、中叶长、中叶面积、生长习性及生长点数呈极显著相关性(见表4),与有无“V”形斑和海拔高度无显著相关;地上生物量与匍匐茎长度、中叶宽、中叶长、中叶面积及生长习性呈极显著相关,与海拔高度呈显著相关;匍匐茎长度与生长点数极显著相关;中叶宽与中叶长、中叶面积、生长习性及生长点数呈极显著相关;中叶长与中叶面积、生长点数及生长习性呈极显著相关;中叶面积与生长习性及生长点数极显著相关;生长习性与生长点数呈极显著相关;生长点数与海拔呈极显著相关性.即随海拔的提高,地上生物量也随之增大,生长习性有由直立生长到匍匐分枝到匍匐不分枝生长的趋势.
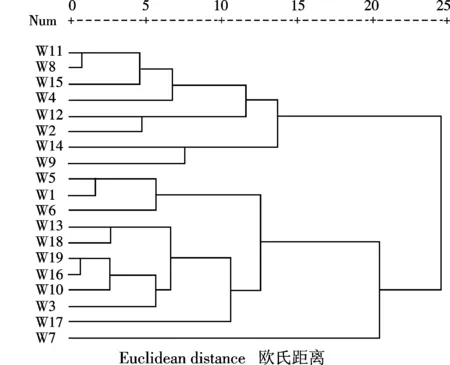
图1 聚类分析Fig.1 Cluster analysis
2.3 基于白三叶形态性状的聚类分析
根据贵州野生白三叶的形态鉴定数据进行聚类分析,在欧氏距离为14.5截距处,可将19份白三叶划分为3个聚类(图1),3个聚类性状平均值见表5.第Ⅰ类包括8份材料,平均株高最矮,地上生物量、匍匐茎长度、中叶宽、中叶长、中叶面积和生长点数的平均值最小,只有少数无“V”形斑点;第Ⅱ类包括10份材料,其生长点数最多,其余性状指标的平均值介于第Ⅰ、Ⅲ类之间,少数无“V”形斑点,大多是匍匐分枝生长;第Ⅲ类只有材料7(汇川区董公市镇,987 m)独自成一类,除匍匐茎长度最短外,生长点数介于第Ⅰ、Ⅱ类之间,全部有“V”形斑点,均匍匐分枝生长,其余形状指标的平均值均大于第Ⅰ、Ⅱ类.由此可知,聚类分组与海拔高度没有明显的关系.
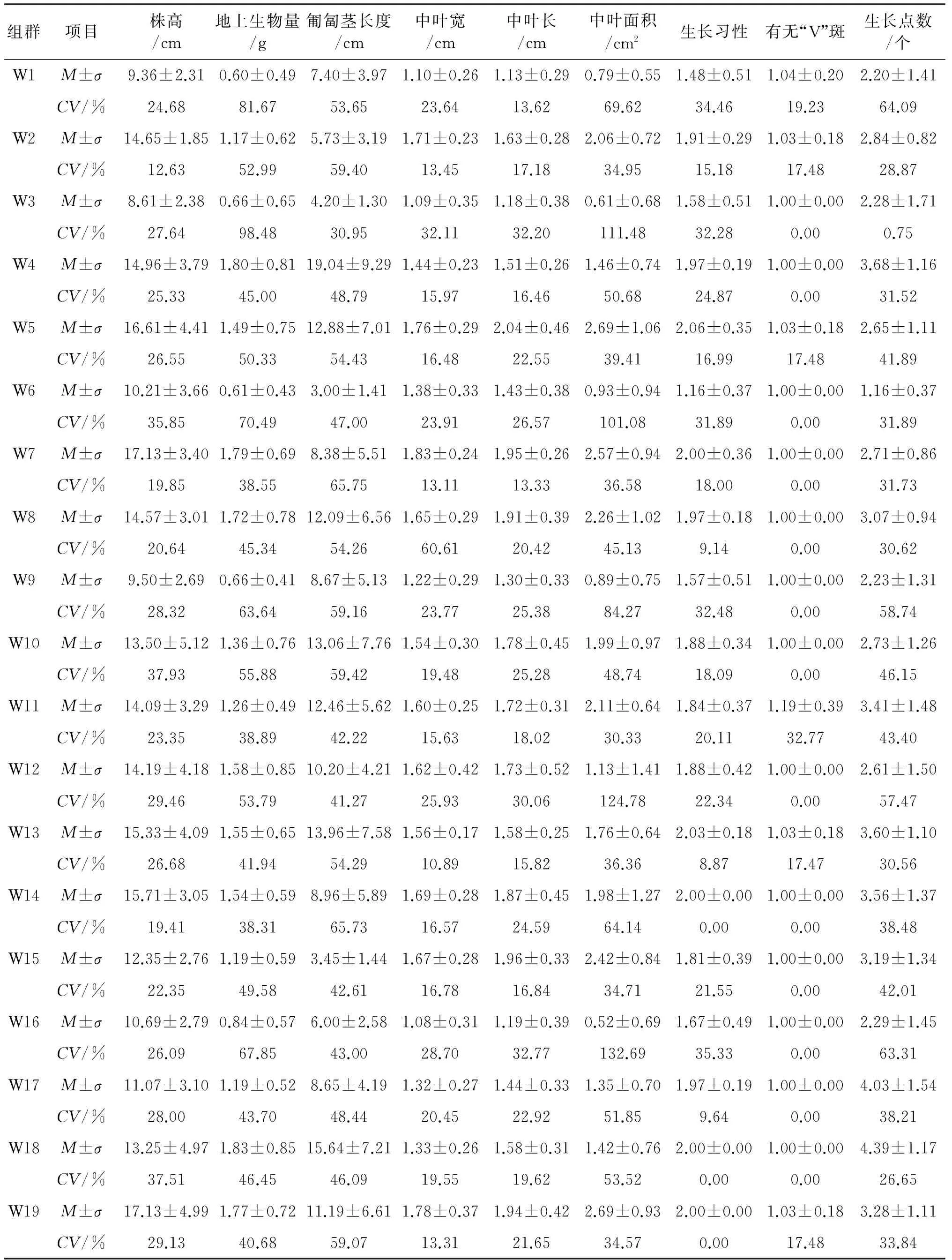
表2 白三叶形态学数量性状分析
注:M,σ,CV分别为平均值、标准差、变异系数.
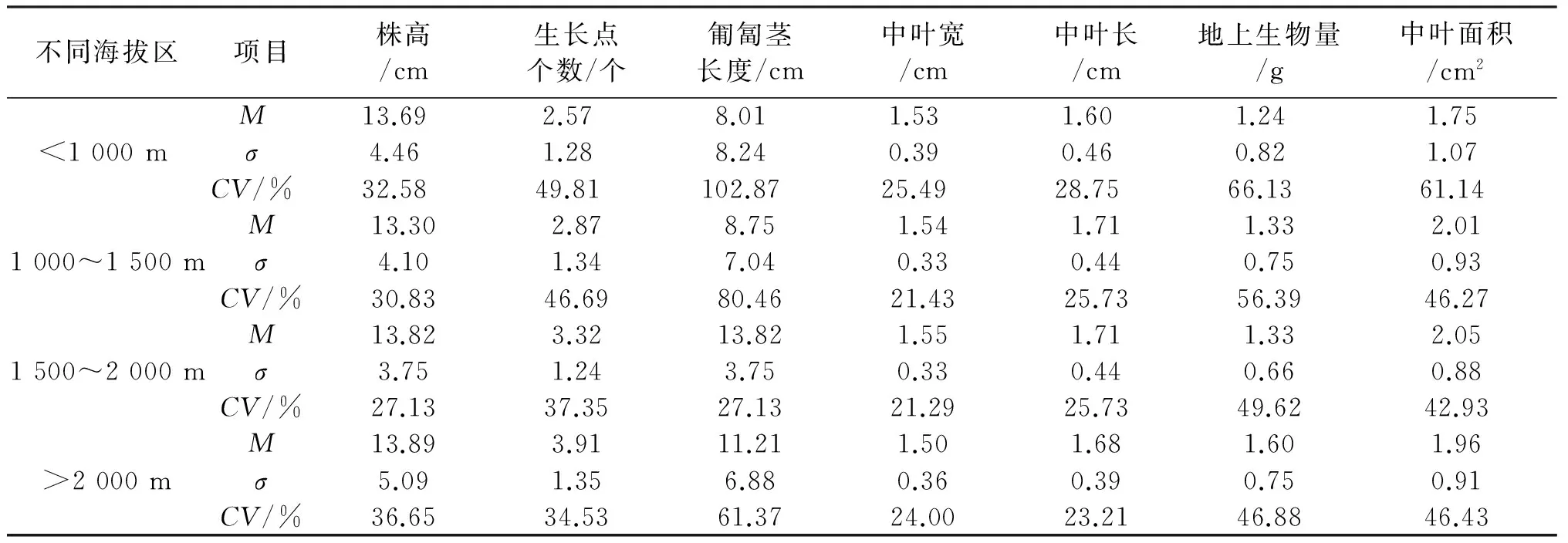
表3 不同海拔高度白三叶形态学数量性状分析
注:M,σ,CV分别为平均值、标准差、变异系数.
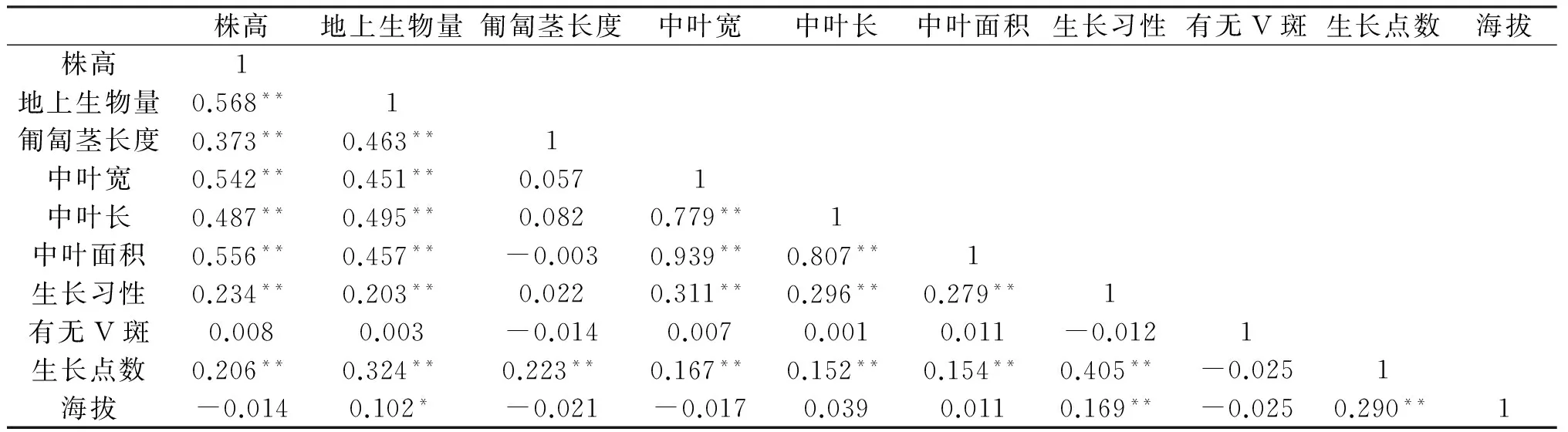
表4 22份贵州白三叶草试验材料形态学性状的相关分析
注:*表示0.05水平下显著,**表示0.01水平下显著.
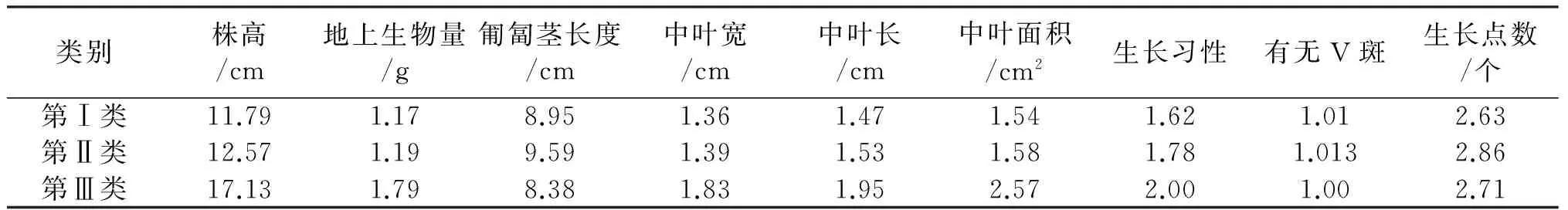
表5 3个类群主要性状平均值
3 讨论与结论
3.1 岩溶地区不同海拔野生白三叶同一花序形态多样性
种质资源是生物遗传变异和生物多样性遗传的物质基础,运用形态学标记研究植物外部形态,受基因及所处生态环境的共同影响.目前,关于开花植物花序内变异原因的研究有很多[7],有研究证明,对不同生态环境下的同种植物的形态学研究,可以了解环境对基因表达的影响[8],而不同海拔区域带来的环境差异会使各类昆虫的活动受到限制或促进,导致不同海拔地区访花者种类和访问比例的差异[9].在同一花序水平上研究不同海拔区域白三叶表型特征,既可以减少基因漂移、花粉污染等外部影响,还可了解由海拔带来的环境差异对其形态的影响,为下一步研究工作提供更精密的数据.本研究结果显示,贵州19份野生白三叶同一花序水平上总变异系数大于300%的有7个居群,其中W16总变异系数最大,无论是居群内还是居群间贵州白三叶形态差异都比新疆[10]白三叶的大;各指标中总变异系数为最大的是中叶面积,其次是地上生物量、匍匐茎长度、生长点数、株高,说明其可塑性较大.贵州因地势起伏较大,生境复杂,蕴藏着丰富的野生植物资源,例如钟理[11]发现贵州野生匐剪股颖各形态学性状均存在较大的变异,形态遗传变异多产生于居群内.本研究材料采集地间海拔跨度较大,小气候类型多样,生境差异明显,为适应不同的生境,白三叶在长期的进化过程中,形态性状也随之改变.
3.2 岩溶地区同一花序野生白三叶的多样性与育种
在育种工作中,借助部分表型性状及其组合,可实现对白三叶目标性状的有效选择.本研究中不同海拔区域甚至同一花序白三叶栽培群体内都存在较丰富的遗传变异,可塑性较大,可为优良类型的选择提供材料.目前,对白三叶形态农艺性状已有较多研究,但主要集中在居群间和居群内水平上[10,12-13],在花序水平上的研究还未见报道.目前分子生物学技术已成为植物育种研究的有效工具,如李润芳[14]运用细胞学方法和SRAP分子标记技术对8种三叶草的种质资源遗传多样性进行了初步评价,张婧源[15]对来自30个国家的70份野生白三叶从表型性状、SRAP分子标记和SSR分子标记上进行了不同产地白三叶种质资源遗传多样性研究.在花序水平上研究其形态多样性,通过人工选育成的优良品种经济性状整齐、遗传基因稳定.因此,对白三叶进行优良品种选育,最终建立良种繁育体系成为获得质量稳定、可控优质白三叶的必经之路.
3.3 岩溶地区贵州野生白三叶的利用价值和开发前景
贵州地处我国云贵高原,属长江、珠江上游地区,是我国典型喀斯特区域之一,地理位置特殊,其生态环境的优劣将直接影响整个长江、珠江流域的生态环境安全.目前,对喀斯特地区生态系统植物恢复多集中在森林生态系统的恢复研究[16],对喀斯特山地草地相关研究还比较少[17].对贵州野生白三叶种质资源进行研究,揭示喀斯特地区白三叶种质资源丰富度、多样性、蕴藏量等特征,可为喀斯特石漠化地区草地植被恢复提供支持.本研究通过统计分析白三叶同一花序水平上形态多样性,发现其可塑性较大,在育种工作中可利用差异较大的材料W16和W12作为亲本培育优质高效品种.还可利用白三叶进行矿山植被修复,有研究发现其能同时富集污染土壤中的铜、镉、铅,且富集量大[18],是很好的矿山修复植被之一.此外,白三叶还是重要的优质豆科牧草之一,影响当地农牧业发展.因此,下一步将全面系统地调查和收集贵州、四川、广西、新疆及云南等地区野生白三叶种质资源,增加其多样性,为充分发挥白三叶资源的生态和经济效益提供支持.
[1] 韦祖生,田益农,马崇熙.作物基因漂移研究综述[J].现代农业科技,2011(13):13-15.
[2] 牟 彤.白三叶化学诱变后代遗传变异的研究[D].哈尔滨:东北农业大学,2013.
[3] 何 静,陈瑞祥,刘秀峰.俄罗斯白三叶在贵州南部的引种试验初报[J].种子,2013,32(3):59-61.
[4] GUSTINE D L,HUFF D R.Genetic variation within and among white clover populations from managed permanent pastures of the Northeastern USA[J].Crop Sci,1999,39(2):524-530.
[5] 曲若竹,侯 林,吕红丽,等.群体遗传结构中的基因流[J].遗传,2004,26(3):377-382.
[6] 李志勇,王宗礼,师文贵,等.牧草种质资源描述规范和数据标准[M].北京:中国农业出版社,2005.
[7] 谢田朋,杜国祯,张格非,等.黄帚橐吾种子生产的花序位置效应及其对幼苗建植的影响[J].植物生态学报,2010,34(4):418-426.
[8] 苗佳敏,钟金城,陈智华.披碱草属种质资源研究现状[J].草业与畜牧,2007,27(11):2222-2227.
[9] 段元文,张挺峰,刘建全.山莨菪(茄科)的传粉生物学[J].生物多样性,2007,15(5):584-591.
[10] 王玉祥,张 博.新疆野生白三叶表型性状变异研究[J].草地学报,2012,20(6):1163-1168.
[11] 钟 理.贵州野生匍匐剪股颖种质资源遗传多样性及越夏生理动态研究[D].成都:四川农业大学,2008.
[12] 李 州,彭 燕,张婧源,等.白三叶种质资源形态变异与地理起源的关系[J].草业科学,2012,29(11):1706-1714.
[13] 王建丽,申忠宝,潘多锋,等.白三叶种质资源主要数量性状的变异与相关性研究[J].中国农学通报,2012,28(26):19-23.
[14] 李润芳.三叶草的细胞学、分子标记及多倍体诱导研究[D].武汉:华中农业大学,2007.
[15] 张婧源.世界范围野生白三叶种质资源的遗传多样性研究[D].成都:四川农业大学,2013.
[16] 魏 媛,张金池,俞元春,等.贵州高原退化喀斯特森林恢复过程中土壤微生物生物量碳、微生物熵的变化[J].农业现代化研究,2009,30(4):487-490.
[17] 张 文,张建利,莫本田,等.喀斯特山地草地植物群落物种数量特征及多样性分析[J].生态环境学报,2011,(5):849-854.
[18] 况 武,田伟莉,高全喜.白三叶在铜、镉、铅复合污染土壤修复上的应用[J].能源工程,2012(6):53-56.
(编辑 WJ)
Study on the Same Inflorescence Morphological Diversity of WildTrifoliumRepensin Karst Region
WUYong-jie1,WANGYuan-su2,LILi1*
(1.College of Life Science,Guizhou Normal University,Guiyang 550001,China; 2.Grassland Monitoring Station of Guizhou Province,Guiyang 550001,China)
By observing the shape index of 19 wildTrifoliumrepens,we compared the morphological diversity of the same inflorescence at different altitudes.Our results imply that the morphological diversity of 19 wildTrifoliumrepenson the same inflorescence at different altitudes has a markedly difference.The total coefficient variation of all above indices obeys the following order: leaf area>above-ground biomass>stolon length>growing point>height >leaf length>leaf width> growth habit>V shape stripe.Between different altitudes inTrifoliumrepensform shape in the group have difference degrees of variations.The clustering analysis found little correlation with the altitude origin.
Karst region; Guizhou;Trifoliumrepens; the same inflorescence; morphological diversity
10.7612/j.issn.1000-2537.2016.05.006
2016-01-18
国家自然科学基金资助项目(31260115);贵州省社发(施秉)项目(11904-0502212Y0043);贵州省重大科技攻关专项(黔科合重大专项字[2011]6009号);贵州省自然科学基金项目(黔科合[J]2009-2091)
*通讯作者,E-mail:liligznu@163.com
S651
A
1000-2537(2016)05-0038-06
