广西扶绥县壮族人群ITGβ1基因多态性与肝癌家系遗传易感性的关系
2016-11-29魏斐斐廖燕赵瑞强王洪学谢裕安
魏斐斐廖燕赵瑞强王洪学谢裕安
作者单位:530021南宁1广西医科大学附属肿瘤医院实验研究部;2广西医科大学生物化学与分子生物学教研室
基础研究
广西扶绥县壮族人群ITGβ1基因多态性与肝癌家系遗传易感性的关系
魏斐斐1廖燕1赵瑞强2王洪学1谢裕安1
作者单位:530021南宁1广西医科大学附属肿瘤医院实验研究部;2广西医科大学生物化学与分子生物学教研室
目的探讨广西扶绥县壮族人群中肝癌高发家系ITGβ1基因rs2298141位点多态性与肝细胞癌(hepatocellular carcinoma,HCC)遗传易感性的关系。方法采用病例-对照研究方法,选取广西扶绥县壮族人群正常对照家系组40名;肝癌高发家系组79例,其中肝癌患者20例,直系亲属59例。采用质谱方法检测ITGβ1基因rs2298141位点基因型分布频率,非条件logistic回归分析不同基因型与HCC发病风险的关系。结果ITGβ1基因rs2298141位点存在AA、AG、GG 3种基因型。ITGβ1基因rs2298141位点基因型分布遵循Hardy-Weinberg遗传平衡定律。ITGβ1基因rs2298141位点A等位基因在正常对照组、肝癌高发家系直系亲属组、肝癌高发家系肝癌患者组中的分布频率分别为63.75%、61.02%、70%,G等位基因分布频率分别36.25%、38.98%和30%,与A等位基因型个体比较,正常对照家系人群中携带G等位基因型的个体发生HCC的风险增加0.75倍(95%CI:0.33~1.7,P=0.50),肝癌高发家系直系亲属组人群发生HCC的风险增加0.67倍(95%CI:0.31~1.45,P=0.31)。正常对照家系组、肝癌高发家系直系亲属组和肝癌高发家系肝癌患者组人群AA基因型分布频率分别为40%、42.73%和50%,AG基因型分别为47.5%、37.29%和40%,GG基因型分别为12.5%、20.34%和10%。与AA基因型个体相比,正常家系组和肝癌高发家系直系亲属组中AG基因型个体发生HCC的风险分别增加0.67倍(95%CI:0.22~2.1,P=0.50)和0.9倍(95%CI:0.31~2.71,P=0.86);GG基因型个体发生HCC的风险分别增加1.05倍(95%CI:0.17~6.6,P=0.96)和2.2倍(95%CI:0.40~11.96,P=0.37),差异均无统计学意义(P>0.05)。结论未发现ITGβ1基因rs2298141位点多态性与广西扶绥县肝癌家族聚集的遗传易感性存在显著相关。
肝肿瘤;ITGβ1基因;单核苷酸多态性;家族聚集性;遗传易感性
肝细胞癌(hepatocellular carcinoma,HCC,以下简称“肝癌”)是常见的恶性肿瘤之一,据统计,2012年约有782 500例肝癌新增病例和745 500例死亡病例,而中国占新发和死亡病例的50%左右[1]。除了环境等外在综合致癌因素外,肝癌的发生和发展与个体间基因型的遗传多态性密切相关。肝癌的发生存在明显的家族聚集倾向,说明遗传因素在肝癌发病中具有重要作用[2]。单核苷酸多态性(single nucleotide polymor-phisms,SNP)作为第3代遗传标记,充分解释了个体之间遗传易感性的差异,是肝癌遗传易感性研究的重要物质[3]。整合素β1(Intergrin β1,ITGβ1)是生物体内一类重要的黏附分子,主要通过介导细胞与胞外基质、细胞与细胞之间的黏附而发挥作用。目前有研究提示,ITGβ1在肿瘤侵袭与转移、血管生成、细胞增殖与凋亡等方面起重要作用[4],但ITGβ1与肝癌的关系尚未见报道。质谱技术具有快速、准确、自动化程度高和高通量等特点。本研究采用质谱技术对广西扶绥县肝癌高发家系与正常对照家系中ITGβ1基因rs2298141位点多态性进行检测与分型,以探讨ITGβ1基因多态性与HCC遗传易感性的关系,为进一步探索HCC发病机制和诊断提供实验依据。
1 资料与方法
1.1研究对象
采用病例-对照研究方法,本研究20个肝癌高发家系,10个正常对照家系均来自肝癌高发区广西扶绥县。20个肝癌高发家系组由肝癌患者20例及其直系亲属59例组成,共79例。肝癌患者为2003年1月至2011年10月在广西医科大学附属肿瘤医院进行外科手术的患者,术后均经组织病理学诊断为HCC,术前未经放化疗或生物治疗。直系亲属指与肝癌患者有直接血缘关系的未患肝癌的亲属,包括一级亲属(父母、子女、兄弟姐妹)和二级亲属(叔伯、堂兄弟、堂姐妹)。经病史采集显示其直系亲属中至少有1人确诊为肝癌。肝癌高发家系组79例中,男性45例,女性34例,年龄16~86岁,中位年龄44岁;AFP(+)14例,AFP(-)65例;HbsAg(+)50例,HbsAg(-)29例;饮酒(≥100 g/d)64例,不饮酒(<100 g/d)15例;吸烟(≥10支/d)19例,不吸烟(<10支/d)60例。
正常对照家系指与肝癌高发家系居住在同一个村(或街道)或生活环境相似的临近村(或街道),民族相同、家庭成员构成和经济状况相似,但与肝癌高发家系无血缘关系的家族。本研究正常对照家系组共入选40名,均选自扶绥县医院的健康体检人群,体检结果均正常,无肝炎、肿瘤及遗传病史,其中男性26名,女性14名,年龄16~85岁,中位年龄47岁;饮酒(≥100 g/d)17名,不饮酒(<100 g/d)23名;吸烟(≥10支/d)15名,不吸烟(<10支/d)25名。正常对照家系组与肝癌高发家系组年龄、性别等比较,差异无统计学意义(P>0.05),具有可比性。本研究按照国家人类基因组研究伦理准则进行,所有入选者均签署知情同意书。
1.2方法
1.2.1主要试剂及仪器血液、细胞、组织DNA提取试剂盒购自北京天根生物科技有限公司;iPLEX反应试剂、384-well SpectroCHIP生物芯片、质谱检测仪及点样仪均购自美国Sequenom公司;HOTSTART Tap酶购自美国Agena Bioscience公司;384孔双头PCR仪购自美国ABI公司,PCR扩增引物、单碱基延伸引物购自深圳华大基因公司。
1.2.2DNA提取实验分析ITGβ1基因型的基因组DNA,采用血液、细胞、组织DNA提取试剂盒提取手术切除的癌组织(20例)标本的DNA。采用血液DNA提取试剂盒提取肝癌高发家系组中的直系亲属(59例)及正常对照家系组静脉血液标本的DNA,置于-20℃冰箱保存备用。
1.3ITGβ1基因型分析
1.3.1引物设计及合成采用美国Sequenom公司Assay Designer 3.1 software设计引物,每个SNP位点对应上下游两条PCR扩增引物和一条延伸引物。PCR上游引物序列为5′-CATGCAAGTTGCAGTTTGC-3′;下游引物序列为5′-CATGCAAGTTGCAGTTTGT-3′;延伸引物序列为5′-CATGCAAGTTGCAGTTTG-3′,引物均由深圳华大基因公司合成。
1.3.2PCR扩增及芯片点样采用HOTSTART Tap酶在384孔板上常规进行40个循环以扩增待测片段,然后将2 μL碱性酸化酶(SAP,美国Sequenom公司)消化液(双蒸水1.53 μL,HME缓冲液0.17 μL,SAP 0.474 U)加到PCR产物中,消化反应(37℃60 min,85℃10 min),以除去未用尽的dNTP。根据Sequenom程序进行引物单碱基延伸反应和延伸产物树脂除盐纯化反应。最后将检测样品从384孔反应板转移到表面覆盖基质的MassARRAY SpectroCHIP芯片[5]。
1.3.3飞行质谱技术检测SNP位点采用美国Sequenom公司Mass ARRAY时间飞行质谱技术进行ITGβ1的SNP基因分型,TYPER 4.0软件分析实验结果,获得分型数据,由深圳华大基因公司的SEQUENOM实验平台支持。SNP分型根据引物延伸反应产物质谱峰的位置确定其分子质量而得出SNP分型结果。同一种颜色可以对应2个或3个峰值,分别为SNP位点的延伸引物峰及两个等位基因峰,若该位点属于纯合子,则仅存在1个等位基因峰;若为杂合子,则存在2个等位基因峰[5]。
1.4统计学处理
采用SPSS 16.0统计软件对数据进行分析。各组基因型分布的Hardy-Weinberg遗传平衡采用拟合优度χ2检验,以判断各组对象是否具有人群代表性。采用χ2检验和非条件logistic回归分析法计算不同等位基因及基因型的比值比(OR)及95%可信区间(95% CI)。以P<0.05为差异有统计学意义。
2 结果
2.1基因型检测结果
检测结果显示ITGβ1基因rs2298141位点存在GG、AG、AA 3种基因型,119例样本中GG基因型19例、AG基因型49例、AA基因型51例。见图1。
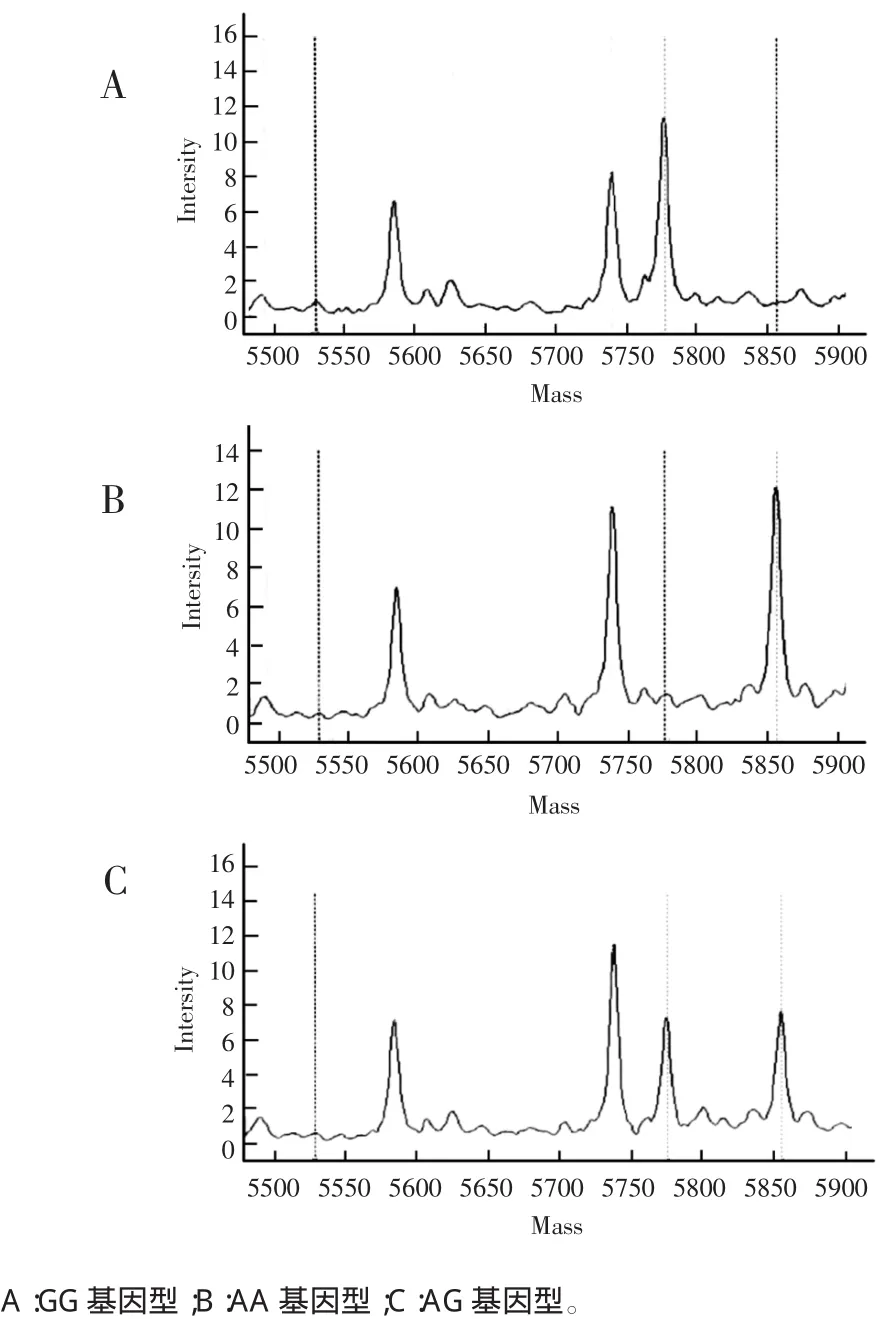
图1 ITGβ1基因(rs2298141)SNP位点基因型分型图
2.2Hardy-Weinberg遗传平衡检验
在正常对照家系组及肝癌高发家系各组中进行Hardy-Weinberg遗传平衡检验,以明确ITGβ1基因rs2298141位点基因型的频率是否遵循遗传平衡定律。结果表明,AA、AG、GG基因型频率在各组中观察值与期望值之间的差异均无统计学意义(P>0.05),符合Hardy-Weinberg遗传平衡,提示各组均具有人群代表性。见表1。
2.3ITGβ1基因rs2298141位点多态性与HCC的关系
ITGβ1基因rs2298141位点等位基因A和G在正常对照家系组中的分布频率分别为63.75%和36.25%;在肝癌高发家系肝癌患者组中的分布频率分别为70%和30%;在肝癌高发家系直系亲属组中的分布频率分别为61.02%和38.98%。经分析,与A等位基因型个体相比,ITGβ1基因rs2298141位点正常对照人群中携带G等位基因型的个体发生HCC的风险增加0.75倍(95%CI:0.33~1.7),差异无统计学意义(P=0.50)。肝癌高发家系直系亲属组人群中携带G等位基因型的个体发生HCC的风险增加0.67倍(95%CI:0.31~1.45),差异亦无统计学意义(P=0.31)。见表2。
ITGβ1基因rs2298141位点AA基因型在正常家系组、肝癌高发家系直系亲属组和肝癌高发家系肝癌患者组中的分布频率分别为40%、42.73%和50%;AG基因型分布频率分别为47.50%、37.29%和40%;GG基因型分布频率分别为12.50%、20.34%和10%。经分析,与AA基因型个体相比,正常家系组和肝癌高发家系直系亲属组成员AG基因型个体发生HCC的风险分别增加0.67倍(95%CI:0.22~2.1,P=0.50)和0.9倍(95%CI:0.31~2.71,P=0.86);正常家系组和肝癌高发家系直系亲属成员GG基因型个体发生HCC的风险分别增加1.05倍(95%CI:0.17~6.6,P=0.96)和2.2倍(95%CI:0.40~11.96,P=0.37),但差异均无统计学意义(P>0.05)。见表2。
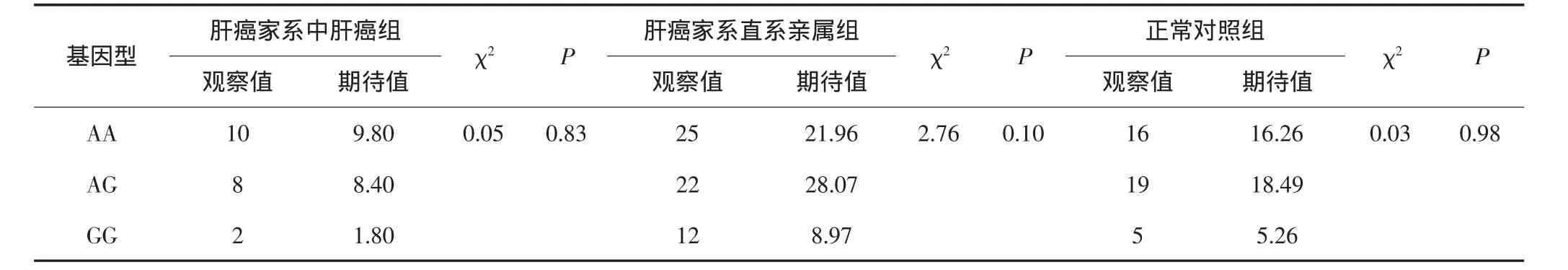
表1 ITGβ1基因rs2298141位点基因型Hardy-Weinberg遗传平衡检验(n)
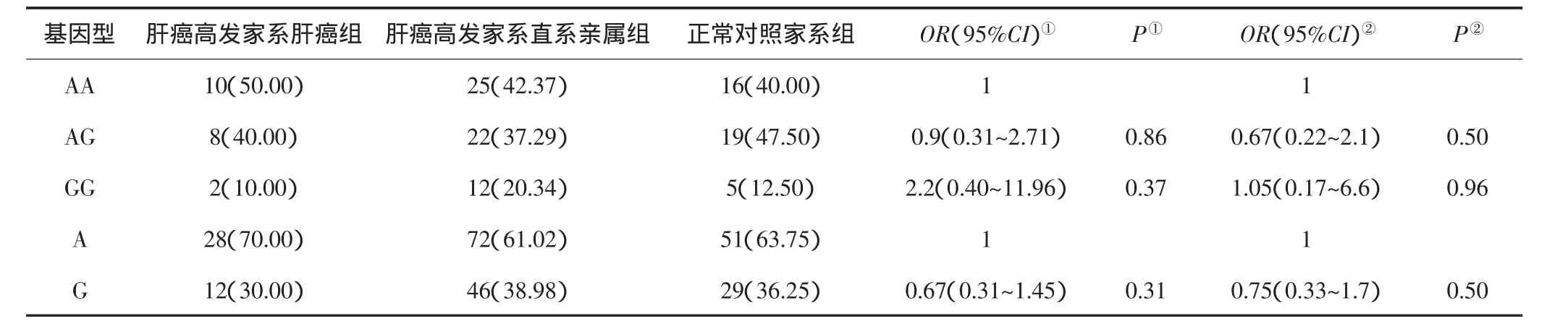
表2 ITGβ1基因rs2298141位点多态性与HCC易感性的关系[n(%)]
3 讨论
肝癌是常见的消化系统肿瘤,流行病学表明,HBV/HCV感染、黄曲霉毒素B1暴露、饮用水污染是肝癌发生的高危因素。但研究发现在同样的致癌因素作用下,只有少部分个体罹患肝癌,这现象说明基于个体差异的遗传易感性的存在对于肝癌的发生具有重要作用。肝癌的家族聚集现象也表明了遗传因素对肝癌发生的作用[2]。研究表明SNP在人群中发生频率>1%,与疾病易感性的改变密切相关[6,7]。ITGβ1作为一种重要的细胞黏附分子,被认为在肿瘤的发生、发展中发挥重要作用。研究发现ITGβ1在甲状腺乳头状瘤[8]、胃癌[9]、乳腺癌[10]、卵巢癌[11]、肺癌[12]、膀胱癌[13]中高表达,并与肿瘤恶性程度呈正相关。在肝癌中也有类似发现,整合素β1蛋白在肝癌组织中的表达显著高于癌旁组织和正常组织[14],抑制整合素β1表达能抑制肝癌细胞增殖及转移[15,16]。
ITGβ基因多态性与各种疾病的关系受到越来越多的关注。Zhou等[17]研究ITGβ1基因rs2230395、rs1187075和rs1187076位点与结直肠癌的关系,发现rs2230395和rs1187075与其总生存期有关,提示ITGβ1的SNP可作为预后判断标志物。Ye等[18]研究发现,ITGβ1基因rs17468位点T基因型出现的频率更高(P<0.05);rs17468的CT和TT基因型与结直肠癌发生危险显著增加有关(OR=1.67,95%CI:1.090~2.559,CTvs TT/CC)。研究发现,ITGβ2的启动子区SNP(rs2070946)与乳头状甲状腺癌发展显著相关[8];ITGβ1-1949/ITGβ1+31804多态性与胃癌关系密切[9];ITGβ1 rs1557150位点多态性与1型糖尿病密切相关[19]。但回顾以往文献,未发现关于ITGβ1基因rs2298141与肝癌遗传易感性的研究,尤其是在肝癌家族聚集性方面。
本研究探讨了广西扶绥县壮族人群肝癌高发家系ITGβ1基因rs2298141位点多态性与HCC遗传易感性的关系,研究结果显示,ITGβ1基因rs2298141位点中,与A等位基因相比,正常对照家系和肝癌高发家系直系亲属组中携带G等位基因型的个体发生HCC的风险分别增加0.75倍和0.67倍,但差异无统计学意义。与AA基因型相比,正常对照家系组和肝癌高发家系直系亲属组GG基因型个体发生HCC的风险分别增加1.05倍和2.2倍,AG基因型个体发生HCC的风险分别增加0.67倍和0.9倍,差异亦无统计学意义。提示ITGβ1基因rs2298141位点与HCC发生无显著相关性。
综上所述,本研究初步探讨了广西扶绥县壮族人群中肝癌高发家系ITGβ1基因rs2298141位点多态性与HCC遗传易感性的关系,结果未发现ITGβ1基因rs2298141位点与HCC遗传易感性的相关性。但因本研究收集病例数有限,且研究人群局限于广西扶绥县的壮族人群等,存在一定局限性,故有关结论仍需扩大样本量及研究范围进一步深入研究。
[1]Torre LA,Bray F,Siegel RL,et al.Global cancer statistics,2012[J]. CA Cancer J Clin,2015,65(2):87-108.
[2]Zhang CY,Huang TR,Yu JH,et al.Epidemiological analysis of primary liver cancer in the early 21st century in Guangxi province of China[J].Chin J Cancer,2010,29(5):545-550.
[3]Taylor JG,Choi EH,Foster CB,et al.Using genetic variation to study human disease[J].Trends Mol Med,2001,7(11):507-512.
[4]Xiong J,Balcioglu HE,Danen EH.Integrin signaling in control of tumor growth and progression[J].Int J Biochem Cell Biol,2013,45(5):1012-1015.
[5]赵辉,王威,张清润,等.高通量飞行时间质谱基因分型方法的研究[J].生物化学与生物物理进展,2005,32(7):667-672.
[6]Chen TP,Lee HL,Huang YH,et al.Association of intercellular adhesion molecule-1 single nucleotide polymorphisms with hepatocellular carcinoma susceptibility and clinicopathologic development[J]. Tumour Biol,2016,37(2):2067-2074.
[7]潘欢欢,苏成豪,林勇,等.驱动蛋白家族成员1B基因rs17401966位点多态性与原发性肝细胞癌遗传易感性的关联研究[J].中华预防医学杂志,2015,49(5):419-423.
[8]Eun YG,KimSK,Chung JH,et al.Association study of integrins beta 1 andbeta2 genepolymorphismandpapillary thyroidcancer[J].AmJ Surg,2013,205(6):631-635.
[9]Tsai YC,HsiaoWH,Lin SH,et al.Genomic singlenucleotidepolymorphisms in the offspring of gastric cancer patients predispose to spasmolytic polypeptide-expressingmetaplasiaafterH.pylori infection[J].J BiomedSci,2015,22(1):16.
[10]Klahan S,Wu MS,Hsi E,et al.Computational analysis of mRNA expression profiles identifies the ITG family and PIK3R3 as crucial genes for regulating triple negative breast cancer cell migration[J]. BiomedResInt,2014,2014:536591.
[11]Villegas-Pineda JC,Garibay-Cerdenares OL,Hernández-Ramírez VI,et al.Integrins and haptoglobin:Molecules overexpressed in ovarian cancer[J].Pathol ResPract,2015,211(12):973-981.
[12]Puig M,Lugo R,Gabasa M,et al.Matrix stiffening and β1 integrin drive subtype-specific fibroblast accumulation in lung cancer[J].Mol CancerRes,2015,13(1):161-173.
[13]Isaac J,TaraporeP,ZhangX,et al.Site-specific S-nitrosylation of integrin alpha 6 increases the extent of prostate cancer cell migration by enhancing integrin beta1 association and weakening adherence to laminin-1[J].Biochemistry,2012,51(48):9689-9697.
[14]ZhaoG,Cui J,QinQ,etal.Mechanical stiffnessof livertissuesinrelation to integrin beta1 expression may influence the development ofhepatic cirrhosis and hepatocellular carcinoma[J].J Surg Oncol,2010,102(5):482-489.
[15]JingY,JiaD,WongCM,etal.SERPINA5 inhibitstumorcell migration by modulatingthefibronectin-integrinbeta1 signalingpathway inhepatocellularcarcinoma[J].Mol Oncol,2014,8(2):366-377.
[16]Zha R,Guo W,Zhang Z,et al.Genome-wide screening identified that miR-134 acts as ametastasis suppressor by targetingintegrin beta1 in hepatocellularcarcinoma[J].PLoS One,2014,9(2):e87665.
[17]Zhou F,Huang X,Zhang Z,et al.Functional polymorphisms of ITGβ1 are associated with clinical outcome of Chinese patients with resected colorectal cancer[J].CancerChemotherPharmacol,2015,75(6):1207-1215.
[18]Ye P,Li Z,Jiang H,et al.SNPs in microRNA-binding sites in the ITGB1 and ITGB3 3'-UTR increase colorectal cancer ris[J]k.Cell BiochemBiophys,2014,70(1):601-607.
[19]Evangelou M,Smyth DJ,Fortune MD,et al.A method for gene-based pathway analysis using genomewide association study summary statistics reveals ninenewtype1 diabetes associations[J].Genet Epidemiol,2014,38(8):661-670.
[2016-06-27收稿][2016-07-24修回][编辑罗惠予]
ITGβ1 polymorphism and genetic susceptibility to hepatocellular carcinoma in families from the Zhuang community in Fusui County,Guangxi
Wei Feifei1,Liao Yan1,Zhao Ruiqiang2,Wang Hongxue1,Xie Yu'an1(1Department of Experimental Research,Affiliated Tumor Hospital of Guangxi Medical University;2Graduate School of Guangxi Medical University,Nanning 530021,P.R.China)
Corresponding author:Xie Yu′an.E-mail:gxxya@aliyun.com
Objective To investigate the correlation between polymorphism at rs2298141 in the integrin β1(ITGβ1)gene and susceptibility of families in Guangxi to hepatocellular carcinoma(HCC).Methods In this study,20 HCC family groups comprising 79 members served as cases,and 10 normal family groups comprising 40 members served as controls.Time-of-flight mass spectrometry was used to determine genotype frequencies at rs2298141,and possible correlations between genotype and HCC risk were explored using non-conditional logistic regression.Results AA,AG and GG genotypes were detected at rs2298141,and their frequencies followed Hardy-Weinberg equilibrium.Frequencies of alleles A and G at rs2298141 were similar among controls,core individuals in HCC families and HCC patients(allele A:63.75%,61.02%,70%;allele G:36.25%,38.98%,30%;P>0.05).Relative to risk of HCC in individuals with allele A,risk of HCC in controls with allele G was 0.75(95%CI 0.33-1.7,P=0.50).Relative to risk of HCC in families with allele A,risk of HCC in HCC families with allele G was 0.67(95%CI 0.31-1.45,P=0.31).AA,AG,and GG genotypes occurred at respective frequencies of 40%,42.73%,and 50% among controls;47.5%,37.29%,and 40%among core individuals in HCC families;and 12.5%,20.34%,and 10%among HCC patients(P>0.05).The corresponding frequencies of genotypes AA,AG and GG in the three groups was 40.00%,47.50%and 12.50%;50.00%,42.73%,37.29%and 20.34%;and 50%,40%and 10%.Allele distributions did not differ significantly between cases and controls.Relative to risk of HCC in individuals with the AA genotype,risk of HCC in control families with genotypes AG or GG was,respectively,0.67(95%CI 0.22-2.1,P=0.50)and 1.05(95%CI 0.17-6.6,P=0.96),while the risk of HCC in HCC families with genotypes AG or GG was,respectively,0.9(95%CI 0.31-2.71,P=0.86)and 2.2(95%CI 0.40-11.96,P=0.37). Conclusion The ITGβ1 rs2298141 polymorphism does not appear to correlate with susceptibility to HCC based on family clustering in Fusui County,Guangxi.
Liver neoplasms;ITGβ1 gene;Single nucleotide polymorphism;Family clustering;Genetic susceptibility
R735.7
A
1674-5671(2016)05-05
10.3969/j.issn.1674-5671.2016.05.02
国家自然科学基金资助项目(81260320)
谢裕安。E-mail:gxxya@aliyun.com
