鄂西北濒危植物红椿天然种群动态分析
2016-11-17闫魁星滕家喜陈文学汪林波
汪 洋, 闫魁星, 滕家喜, 陈文学, 汪林波, 陈 松,①
(1. 湖北生态工程职业技术学院园林与建工学院, 湖北 武汉 430200; 2. 华中农业大学园艺林学学院, 湖北 武汉 430072;3. 竹山县林业局, 湖北 竹山 442200; 4. 谷城县林业局, 湖北 谷城 441799)
鄂西北濒危植物红椿天然种群动态分析
汪洋1, 闫魁星1, 滕家喜2, 陈文学3, 汪林波4, 陈松4,①
(1. 湖北生态工程职业技术学院园林与建工学院, 湖北 武汉 430200; 2. 华中农业大学园艺林学学院, 湖北 武汉 430072;3. 竹山县林业局, 湖北 竹山 442200; 4. 谷城县林业局, 湖北 谷城 441799)
采用相邻格子法对分布于鄂西北的3个红椿(ToonaciliataRoem.)天然种群(竹山、谷城T1和谷城T2)进行统计;采用立木径级结构代替种群年龄结构的方法,分析了各种群不同龄级的个体数量动态指数(Vn)、静态生命表及存活曲线,并对3个种群进行谱分析。结果表明:3个红椿种群均属于增长型种群,自然状态下竹山、谷城T1和谷城T2种群个体数量动态指数分别为46.27%、53.46%和37.71%,标准化存活曲线分别为Deevey Ⅲ型、Deevey Ⅱ型和Deevey Ⅱ型。3个种群第2龄级〔2.5 cm≤胸径(DBH)<7.5 cm〕幼树的死亡率均较高,竹山种群第3龄级(7.5 cm≤DBH<12.5 cm)幼树的死亡率最高,谷城T1种群第7龄级(27.5 cm≤DBH<32.5 cm)大树的死亡率最高,谷城T2种群第6龄级(22.5 cm≤DBH<27.5 cm)中树的死亡率最高;3个种群第1龄级(DBH<2.5 cm)和第2龄级个体的平均期望寿命均较高。 竹山、 谷城T1和谷城T2种群的基波振幅分别为1.061 6、 1.097 1和 0.788 2,明显的小周期分别出现在第4、第2和第4波序,表明分布于鄂西北的红椿天然种群更新存在着周期性,且存在小周期的多谐波叠加,种群波动与龄级的Vn值相吻合。种群动态分析结果表明:由于红椿的强阳性生理特征,其天然种群受环境筛抑制和环境干扰,造成幼树个体损失和中龄级个体不足,这是造成红椿濒危的主要因素,因此,建议加强人为正向“干扰”以制造林窗、增强种群天然更新能力、引种造林,以保护和利用红椿这一濒危植物资源。
鄂西北; 红椿; 种群动态; 静态生命表; 存活曲线; 谱分析
植物种群结构和动态是植物个体生存能力与外界环境相互作用的结果[1]。种群年龄结构反映了种群在特定时间世代重叠的特征,是研究种群动态的有效方法,被广泛应用于种群动态研究[2]。植物天然种群存在复杂的周期现象,可以由不同振幅和相应的谐波组成[3],种群天然更新动态可通过不同龄级个体的数量分布波动来表现[4-5]。
红椿(ToonaciliataRoem.)又名红楝子,落叶或近常绿乔木;其木材红褐色、纹理通直、结构细致且耐腐,是珍贵速生用材树种[6]。红椿自身的生理特性导致其天然更新缓慢,加之环境变化和人为干扰,致使其天然种群数量少、分布区缩小,已被列为野生濒危物种[7]。目前对红椿的研究涵盖种质资源、生理、育种、栽培、造林、医药及资源保护等方面[8-9],而有关其群落和种群生态学的研究却很少。
作者在红椿种质资源收集和优树选择过程中,通过多次走访和实地踏查,在鄂西北地区的竹山和谷城发现3个红椿天然群落。以此为研究对象,作者开展了种群个体数量动态、特定年龄生命表以及Fourier 级数谱分析,并对群落数量动态进行研究,以期揭示红椿种群的数量动态特征,并为红椿濒危机制研究及其保护策略的制订奠定研究基础,并为红椿天然林资源的保护和可持续利用开发提供参考资料。
1 研究地概况和研究方法
1.1研究地概况
竹山红椿种群位于堵河源国家级自然保护区的核心区,属北亚热带湿润气候,年平均气温12.9 ℃,≥0 ℃年积温4 628 ℃,≥10 ℃年积温3 750 ℃,无霜期219 d,年平均降水量在1 000 mm以上;土壤为石灰岩、板页岩和砂页岩等发育的山地黄棕壤[10]。谷城县的2个红椿种群位于谷城县紫金镇玛瑙观村南河流域的南河国家级自然保护区,属北亚热带季风气候,年平均气温16.0 ℃,无霜期234 d,年均降水量934.5 mm[11];地质结构为武当变质岩和石灰岩,土壤为黄棕壤。参照蔡秀珠[12]和杨梅等[13]的方法,并根据实际调查结果,将3个红椿天然种群的人为干扰程度进行分级和赋值:轻微干扰(0.1~0.3),中度干扰(0.4~0.6),重度干扰(0.7~0.9)。3个红椿群落的环境特征数据具体见表1。
竹山红椿群落与竹林伴生,乔木主要有皂荚(GleditsiasinensisLam.)、枫杨(PterocaryastenopteraC. DC.)和楠木(PhoebezhennanS. Lee);灌木主要有小叶女贞(LigustrumquihouiCarr.)、水麻(DebregeasiaorientalisC. J. Chen)和山鸡椒〔Litseacubeba(Lour.) Pers.〕;草本主要有芒(MiscanthussinensisAnderss.)、马兰〔Kalimerisindica(Linn.) Sch.-Bip.〕和鬼针草(BidenspilosaLinn.)等。谷城红椿群落内乔木主要有红椿和宜昌润楠(MachilusichangensisRehd. et Wils.)等;灌木主要有金花忍冬(LonicerachrysanthaTurcz.)、黄栌(CotinuscoggygriaScop.)和棠叶悬钩子(RubusmalifoliusFocke)等;草本主要有大叶贯众〔Cyrtomiummacrophyllum(Makino) Tagawa〕和大叶冷水花〔Pileamartinii(Levl.) Hand.-Mazz.〕等。群落内蕨类和藤本较为丰富,群落结构较为完整,群落优势种为红椿。
表1鄂西北3个红椿群落的环境特征
Table 1Environmental characteristics of three communities ofToonaciliataRoem. in northwestern Hubei
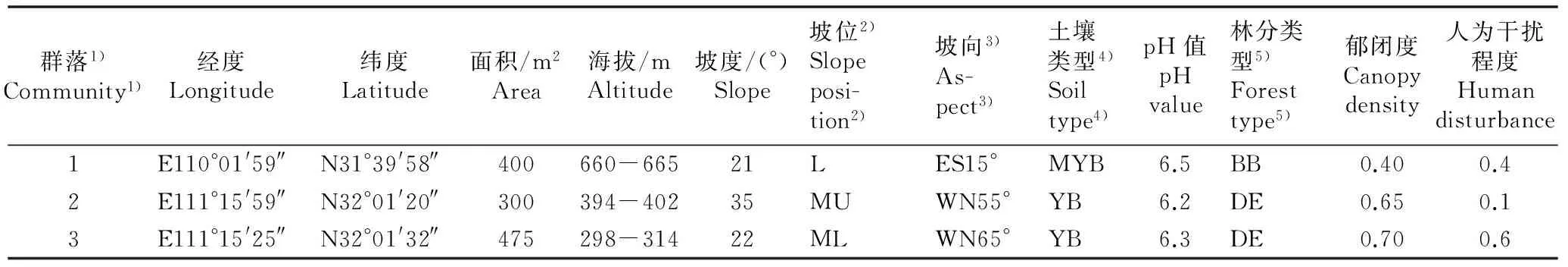
群落1)Community1)经度Longitude纬度Latitude面积/m2Area海拔/mAltitude坡度/(°)Slope坡位2)Slopeposi-tion2)坡向3)As-pect3)土壤类型4)Soiltype4)pH值pHvalue林分类型5)Foresttype5)郁闭度Canopydensity人为干扰程度Humandisturbance1E110°01'59″N31°39'58″400660-66521LES15°MYB6.5BB0.400.42E111°15'59″N32°01'20″300394-40235MUWN55°YB6.2DE0.650.13E111°15'25″N32°01'32″475298-31422MLWN65°YB6.3DE0.700.6
1)1: 竹山Zhushan; 2: 谷城T1 Gucheng T1; 3: 谷城T2 Gucheng T2.
2)L: 下坡位Lower slope; MU: 中上坡位Middle and upper slope; ML: 中下坡位Middle and lower slope.
3)SE: 东偏南East by south; WN: 西偏北West by north.
4)MYB: 山地黄棕壤Mountain yellow-brown soil; YB: 黄棕壤Yellow-brown soil.
5)BB: 阔叶竹叶林Broad-leaved and bamboo forest; DE: 落叶常绿阔叶林Deciduous and evergreen broad-leaved forest.
1.2研究方法
1.2.1样地设计及划分依据红椿种群现存规模,采用相邻格子法设立面积5 m×5 m样方,样地完全覆盖种群。竹山种群设立面积40 m×10 m样地,共400 m2;位于谷城三道沟的T1种群分段划定面积20 m×10 m和10 m×10 m样地各1个,面积共300 m2;位于谷城二道沟的T2种群划定1个面积20 m×10 m样地、2个面积10 m×10 m样地、1个面积5 m×10 m样地和1个面积5 m×5 m样地,面积共475 m2。对红椿进行每木调查,对胸径(DBH)<2.5 cm的个体测量地径和高度;对DBH≥2.5 cm的个体测量株高、DBH、冠幅、枝下高和分枝角等。同时调查并记录环境因子,统计林下乔木、灌木、草本及藤本的种类和数量。
1.2.2龄级划分及动态分析方法根据“红椿种群的年龄与DBH具有显著相关性[14]”的特点,采用“空间替代时间法”,以立木径级结构代替种群年龄结构[15-16]分析红椿种群动态。龄级划分标准为:幼苗级1(DBH<2.5 cm);从第2龄级开始,以DBH 5.0 cm为步长,即幼树级2(2.5 cm≤DBH<7.5 cm), 幼树级3 (7.5 cm≤DBH<12.5 cm), 幼树级4 (12.5 cm ≤DBH<17.5 cm),中树级5(17.5 cm≤DBH<22.5 cm),中树级6(22.5 cm≤DBH<27.5 cm),大树级7(27.5 cm≤DBH<32.5 cm),大树级8(32.5 cm≤DBH<37.5 cm),大树级9(DBH≥37.5 cm)。据此,将竹山种群划分为8个龄级, 谷城T1种群划分为9个龄级,谷城T2种群划分为8个龄级。
采用陈晓德[17]的数学模型,推导种群年龄结构的个体数量动态指数(Vn),对红椿种群个体数量动态进行定量描述。Vn、Vpi(自然状态下种群个体数量动态指数)和Vpi′(外部干扰时种群个体数量动态指数)取正值、负值和零值的意义分别反映相邻龄级或种群个体数量的增长、衰退和稳定的动态关系[18]。
1.2.3生命表编制与标准化存活曲线分析根据龄级划分结果,统计样地内各龄级个体数量,计算生命表数据[15-16,19-20]。由于3个红椿种群各龄级个体数量有波动,不能满足生命表的3个假设,因此,采用匀滑技术对数据进行处理,编制红椿种群静态生命表[15,21]。参照文献[22]的方法,检验分析3个红椿种群标准化存活曲线类型并分析其形成机制。
1.2.4种群谱分析谱分析可以揭示种群数量的周期性波动,是探讨林分分布波动性和年龄更替过程周期性的数学工具。各世代形成的一定时间间隔的规律性即为种群天然更新过程的周期性[4]。利用谱分析公式计算各波形的振幅Ak(k=1,2,3,……,P),其中,A1为基波振幅,A2、A3、A4、……、AP为各谐波振幅;各谐波周期分别是基本周期的1/2、 1/3、……、 1/P;P=n/2;n为种群全部龄级级差。Ak值的大小差异反映了各周期作用大小的差异[3-5]。
2 结果和分析
2.1红椿种群动态分析
鄂西北3个红椿种群个体数量变化动态见表2。结果显示:竹山种群在V2(第2到第3龄级)、V4(第4到第5龄级)和V7(第7到第8龄级)龄级的个体数量动态指数分别为-21.74%、-43.75%和-50.00%,而V1(第1到第2龄级)、V3(第3到第4龄级)、V5(第5到第6龄级)和V6(第6到第7龄级)龄级的个体数量动态指数均为正值,自然状态下种群个体数量动态指数(Vpi)为46.27%;考虑外部干扰时种群个体数量动态指数(Vpi′)为5.78%,竹山种群整体为增长型种群。 谷城T1种群在V2、V3、V5和V7龄级个体数量动
表2鄂西北3个红椿种群不同龄级的个体数量动态指数1)
Table 2Individual quantity dynamic index in different age classes of three populations ofToonaciliataRoem. in northwestern Hubei1)
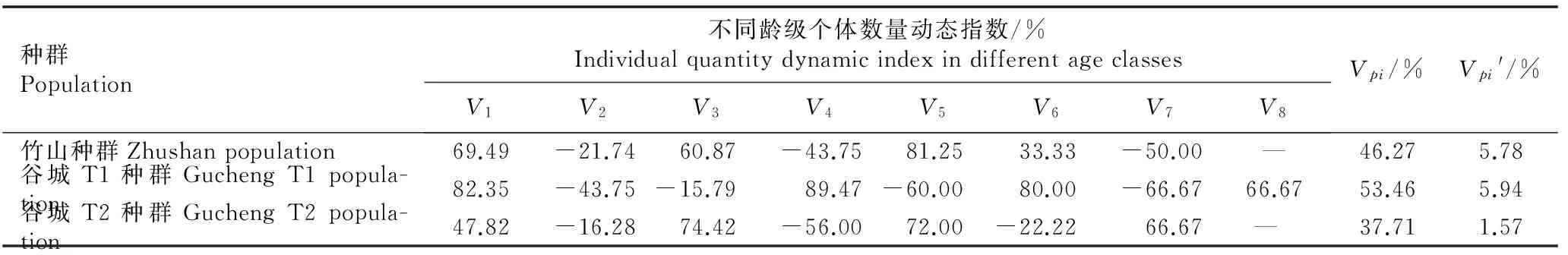
种群Population不同龄级个体数量动态指数/%IndividualquantitydynamicindexindifferentageclassesV1V2V3V4V5V6V7V8Vpi/%Vpi'/%竹山种群Zhushanpopulation69.49-21.7460.87-43.7581.2533.33-50.00—46.275.78谷城T1种群GuchengT1popula-tion82.35-43.75-15.7989.47-60.0080.00-66.6766.6753.465.94谷城T2种群GuchengT2popula-tion47.82-16.2874.42-56.0072.00-22.2266.67— 37.711.57
1)V1: 第1到第2龄级个体数量动态指数Individual quantity dynamic index from the 1st to the 2nd age class;V2: 第2到第3龄级个体数量动态指数Individual quantity dynamic index from the 2nd to the 3rd age class;V3: 第3到第4龄级个体数量动态指数Individual quantity dynamic index from the 3rd to the 4th age class;V4: 第4到第5龄级个体数量动态指数Individual quantity dynamic index from the 4th to the 5th age class;V5: 第5到第6龄级个体数量动态指数Individual quantity dynamic index from the 5th to the 6th age class;V6: 第6到第7龄级个体数量动态指数Individual quantity dynamic index from the 6th to the 7th age class;V7: 第7到第8龄级个体数量动态指数Individual quantity dynamic index from the 7th to the 8th age class;V8: 第8到第9龄级个体数量动态指数Individual quantity dynamic index from the 8th to the 9th age class;Vpi: 自然状态下种群个体数量动态指数Individual quantity dynamic index of population in natural state;Vpi′: 外部干扰时种群个体数量动态指数Individual quantity dynamic index of population under external disturbance. —: 无对应龄级No corresponding age class.
态指数分别为-43.75%、 -15.79%、 -60.00%和 -66.67%,说明该种群这4个龄级更新较差,但V1、V4、V6和V8(第8到第9龄级)龄级个体数量保持较强的增长性,Vpi′值为5.94%,在3个种群中最大,说明该种群为增长型种群,抗风险能力最强。谷城T2种群中虽然仅有V2、V4和V6龄级的个体数量动态指数出现负增长,但V1龄级个体数量动态指数较低,人为干扰最大, 导致种群Vpi值为37.71%, 且Vpi′值仅为1.57%,远小于前2个种群;谷城T2种群虽然也为增长型种群,但抗干扰能力下降,种群增长缓慢。3个种群不同龄级个体数量动态指数的变化,与人为干扰及其生理习性相关。
2.2红椿种群静态生命表与存活曲线分析
2.2.1静态生命表分析鄂西北不同红椿种群的静态生命表见表3。3个红椿种群的年龄结构均较为完整,幼苗数量较大,第2龄级幼树均损失较大,第3龄级存活个体数均较第2龄级有所提高;竹山和谷城T2种群第4龄级以及谷城T1种群第5龄级的存活个体数损失也较大,3个种群高龄级的存活个体数均较少。竹山和谷城T1种群标准化死亡个体数(dx)在第1至第8龄级随龄级增大逐步下降,第9龄级又升高;而谷城T2种群dx值在第1至第9龄级随龄级增大逐步下降。竹山种群个体死亡率(qx)在第3龄级最高(为0.609), 在第4龄级最低(为0.111), 说明在该种群第1至第3龄级间环境筛的作用逐渐加强,到第4龄级时因光照条件改善,qx值下降,种群逐渐稳定发展。谷城T1种群受人为干扰最小,第7龄级qx值最高,为0.750,表明群落内的种内和种间竞争使自疏加剧,红椿个体在进入林冠层前损失较大;第2龄级qx值为0.600,表明在生长过程中可能由于光照条件不足,幼树个体损失较大;第8龄级qx值为0.000,说明该龄级的红椿个体已经进入群落上层,获得竞争优势。谷城T2种群受人为干扰最强,在第1龄级qx值最低,为0.194, 幼苗数量也最多; 在第7龄级qx值最高, 为0.667,在此龄级受到的干扰除种群自疏外,还存在人为砍伐现象。
表3鄂西北3个红椿种群的静态生命表1)
Table 3Static life table of three populations ofToonaciliataRoem. in northwestern Hubei1)
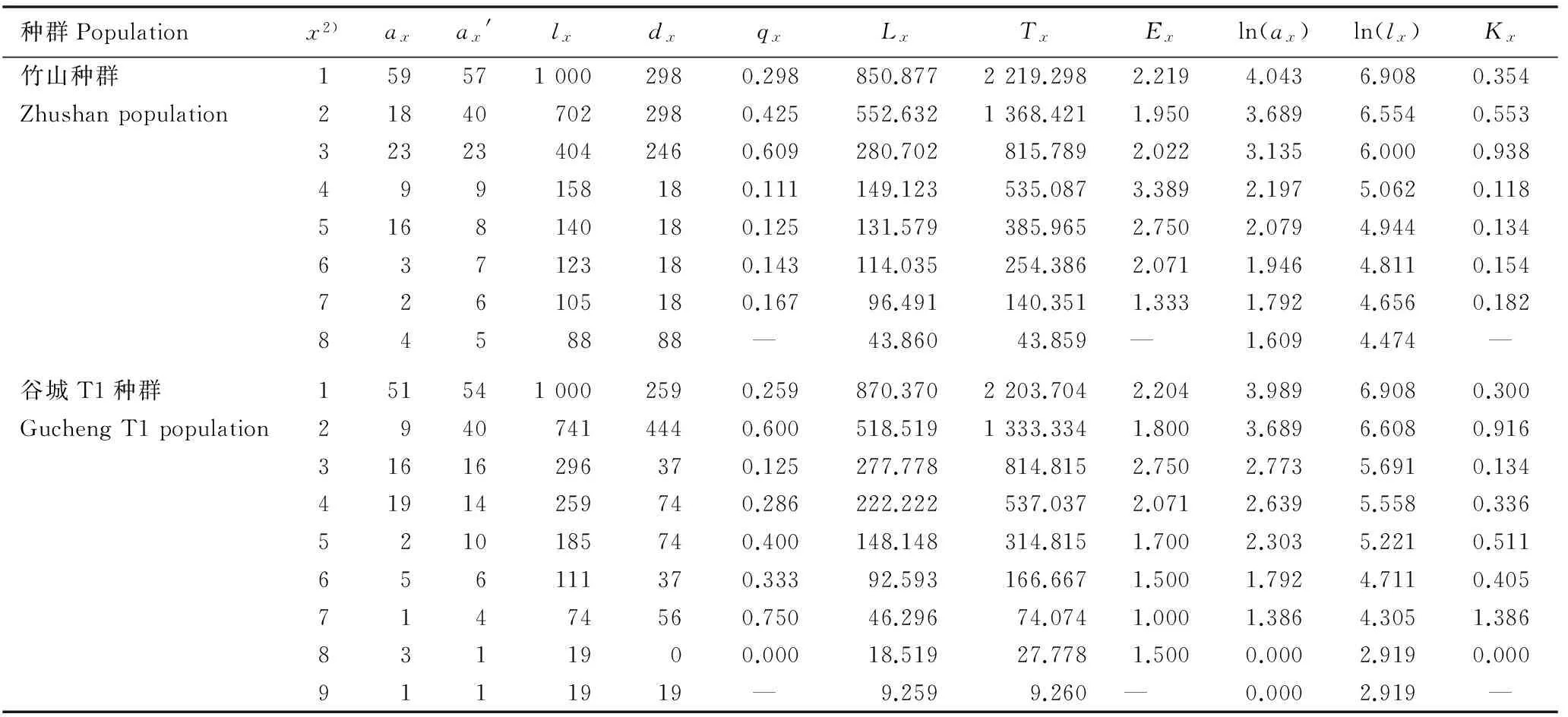
种群Populationx2)axax'lxdxqxLxTxExln(ax)ln(lx)Kx竹山种群1595710002980.298850.8772219.2982.2194.0436.9080.354Zhushanpopulation218407022980.425552.6321368.4211.9503.6896.5540.553323234042460.609280.702815.7892.0223.1356.0000.938499158180.111149.123535.0873.3892.1975.0620.1185168140180.125131.579385.9652.7502.0794.9440.134637123180.143114.035254.3862.0711.9464.8110.154726105180.16796.491140.3511.3331.7924.6560.1828458888— 43.86043.859— 1.6094.474—谷城T1种群1515410002590.259870.3702203.7042.2043.9896.9080.300GuchengT1population29407414440.600518.5191333.3341.8003.6896.6080.91631616296370.125277.778814.8152.7502.7735.6910.13441914259740.286222.222537.0372.0712.6395.5580.3365210185740.400148.148314.8151.7002.3035.2210.511656111370.33392.593166.6671.5001.7924.7110.40571474560.75046.29674.0741.0001.3864.3051.3868311900.00018.51927.7781.5000.0002.9190.0009111919— 9.2599.260— 0.0002.919—
续表3Table 3 (Continued)
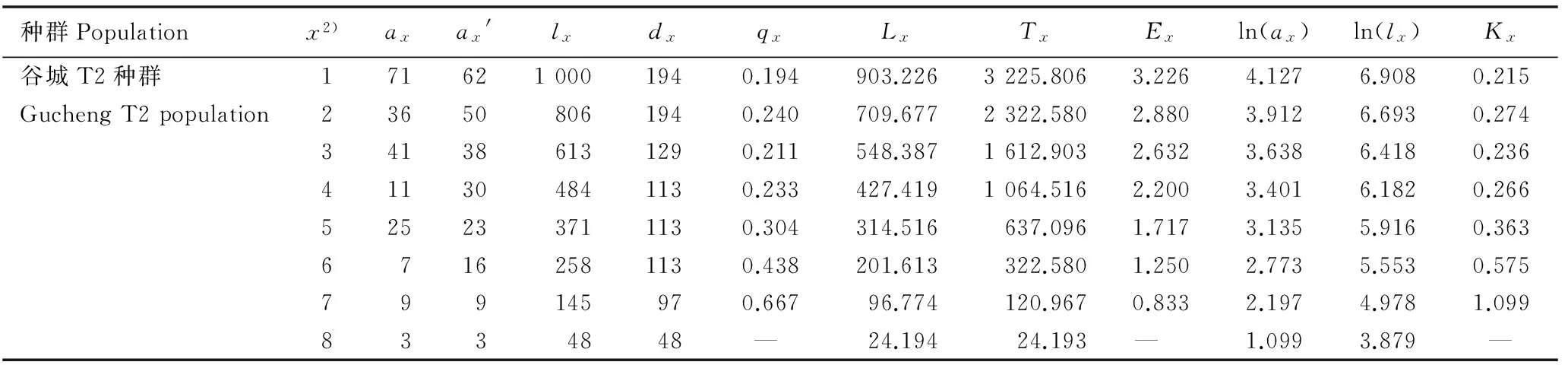
种群Populationx2)axax'lxdxqxLxTxExln(ax)ln(lx)Kx谷城T2种群1716210001940.194903.2263225.8063.2264.1276.9080.215GuchengT2population236508061940.240709.6772322.5802.8803.9126.6930.274341386131290.211548.3871612.9032.6323.6386.4180.236411304841130.233427.4191064.5162.2003.4016.1820.266525233711130.304314.516637.0961.7173.1355.9160.36367162581130.438201.613322.5801.2502.7735.5530.575799145970.66796.774120.9670.8332.1974.9781.0998334848— 24.19424.193— 1.0993.879—
1)x: 龄级Age class;ax:x龄级存活个体数Survival number atxage class;ax′: 匀滑处理后的x龄级存活个体数 Homogenized survival number atxage class;lx:x龄级开始时的标准化存活个体数Standardized survival number starting fromxage class;dx:x到x+1龄级间隔期内标准化死亡个体数Standardized mortality fromxtox+1 age class;qx:x到x+1龄级间隔期内死亡率Mortality rate fromxtox+1 age class;Lx:x到x+1龄级间隔期间内存活个体数Survival number fromxtox+1 age class;Tx:x龄级及x龄级以上的存活个体总数Total survival number fromxage class to higher age class;Ex: 进入x龄级个体平均期望寿命Average life expectancy of individual intoxage class; ln(ax):ax的自然对数Natural logarithm ofax; ln(lx):lx的自然对数Natural logarithm oflx;Kx: 损失度Loss rate. —: 无对应理论值 No corresponding theoretical value.
2)1: DBH<2.5 cm; 2: 2.5 cm≤DBH<7.5 cm; 3: 7.5 cm≤DBH<12.5 cm; 4: 12.5 cm≤DBH<17.5 cm; 5: 17.5 cm≤DBH<22.5 cm; 6: 22.5 cm≤DBH<27.5 cm; 7: 27.5 cm≤DBH<32.5 cm; 8: 32.5 cm≤DBH<37.5 cm; 9: DBH≥37.5 cm.
期望寿命(Ex)反映各龄级内个体的平均生存能力[23]。竹山种群的Ex值在第4龄级达到峰值, 为3.389,体现出最强生命力和生存质量。谷城T1种群的Ex值在第3龄级达到峰值,为2.750,其较低的峰值说明该种群中个体竞争较强,种群处于强烈淘汰期,也是红椿种群的自然发展模式。谷城T2种群的Ex值在第1龄级达到峰值,为3.226,高于同龄级其他2个种群,说明该种群幼苗期生长活动最旺盛,生存质量最高,且随龄级增加个体间竞争逐渐加强,生存质量逐渐下降。3个红椿种群损失度(Kx)与qx值变化幅度略有不同,但各种群Kx值与qx值的变化趋势一致。
2.2.2存活曲线分析存活曲线是借助于存活个体数量来描述特定龄级存活数量,或通过特定年龄组对应个体数量的自然对数值作图[19-20]。本文以某龄级开始时的标准化存活个体数的自然对数ln(lx)为纵坐标,以龄级为横坐标作图,获得红椿种群的存活曲线(图1)。对标准化存活曲线回归,对比决定系数R2和F值,选择不同存活曲线方程,结果见表4。竹山、谷城T1和谷城T2种群的标准化存活曲线分别为Deevey Ⅲ型、Deevey Ⅱ型和Deevey Ⅱ型。随龄级增大,竹山种群个体数量逐渐减少;谷城T1种群在第2、第4和第5龄级个体数量损失较大, 其他龄级较稳定, 表现出红椿强阳性的特点及环境筛的抑制作用;谷城T2种群在第2至第7龄级个体数量损失均较大,体现了红椿天然种群的自疏特点,同时人为干扰是不可忽视的重要因素,这与恩施红椿种群动态的特点一致[24]。

—◆—: 竹山种群Zhushan population; —■—: 谷城T1种群Gucheng T1 population; —▲—: 谷城T2种群Gucheng T2 population.
ln(lx): 某龄级开始时的标准化存活个体数的自然对数 Natural logarithm of standardized survival number starting from some age class.
1: DBH<2.5 cm; 2: 2.5 cm≤DBH<7.5 cm; 3: 7.5 cm≤DBH<12.5 cm; 4: 12.5 cm≤DBH<17.5 cm; 5: 17.5 cm≤DBH<22.5 cm; 6: 22.5 cm≤DBH<27.5 cm; 7: 27.5 cm≤DBH<32.5 cm; 8: 32.5 cm≤DBH<37.5 cm; 9: DBH≥37.5 cm.
图1鄂西北3个红椿种群标准化存活曲线
Fig. 1Standardized survival curve of three populations ofToonaciliataRoem. in northwestern Hubei
表4鄂西北3个红椿种群标准化存活曲线的回归方程
Table 4Regression equation of standardized survival curve of three populations ofToonaciliataRoem. in northwestern Hubei
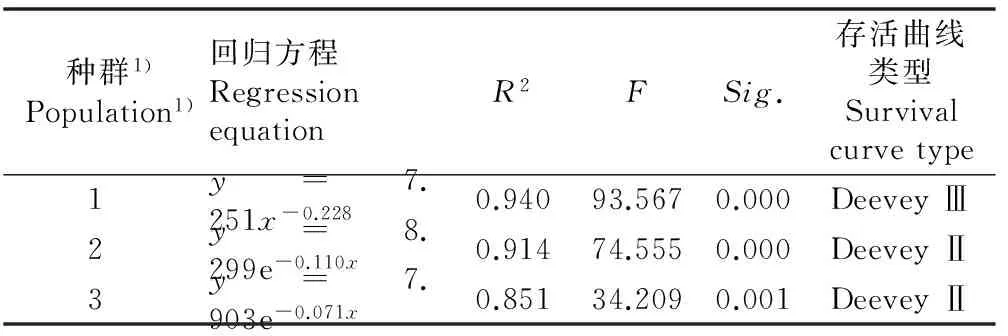
种群1) Population1)回归方程RegressionequationR2FSig.存活曲线类型Survivalcurvetype1y=7.251x-0.2280.94093.5670.000DeeveyⅢ2y=8.299e-0.110x0.91474.5550.000DeeveyⅡ3y=7.903e-0.071x0.85134.2090.001DeeveyⅡ
1)1: 竹山种群Zhushan population; 2: 谷城T1种群Gucheng T1 population; 3: 谷城T2种群Gucheng T2 population.
2.3红椿种群的谱分析
通过种群不同龄级与未经匀滑处理的实际存活数量分布波动探讨红椿种群的动态,以生命表的龄级作为波谱分析的级差,3个种群的龄级级差(n)均按8龄级处理,总波序(k)=n/2=4。鄂西北3个红椿种群的周期性波动见表5。
由表5可见:竹山种群基波振幅(A1)为1.061 6,为种群本身所固有,反映种群波动特性[25];竹山种群个体数量动态除受基波影响外,还显示出明显的小周期波动,第4波序振幅(A4)为0.548 1,对应的胸径为10 cm,即第3龄级,对应种群个体数量动态指数(V2)为-21.74%,说明小周期的形成来自于种群数量动态变化;竹山种群小周期波动区间约为9~12 a(A4),是该种群理论上的最小周期。谷城T1种群A1值为1.097 1, 明显的小周期波动在第3波序, 第3波序振幅(A3)为0.849 0,该种群小周期波动约13~16 a,显示在第4龄级,是该种群理论上的最小周期。谷城T2种群A1值为0.788 2,明显的小周期波动在第3波序,A4值为1.006 7,反映在第3龄级,与竹山种群的小周期波动一致,可能与这2个种群均受到较强人为干扰有关。
表5鄂西北3个红椿种群的周期性波动
Table 5Periodical fluctuation of three populations ofToonaciliataRoem. in northwestern Hubei

种群1)Population1)n2)各谐波振幅3)Amplitudeofeachharmonicwave3)A1A2A3A4181.06160.45020.04120.5481291.09710.15020.84900.0006380.78820.34640.04301.0067
1)1: 竹山种群Zhushan population; 2: 谷城T1种群Gucheng T1 population; 3: 谷城T2种群Gucheng T2 population.
2)n: 种群全部龄级级差Age class range of each population.
3)A1: 基波振幅Amplitude of fundamental wave;A2: 第2波序振幅Amplitude of the 2nd harmonic wave;A3: 第3波序振幅Amplitude of the 3rd harmonic wave;A4: 第4波序振幅Amplitude of the 4th harmonic wave.
综上所述,红椿种群天然更新存在周期性,且所表现出的波动不是单一周期。除基波振幅A1外,各种群在不同龄级表现出小周期,种群由2个或以上的周期迭加,因此认为红椿天然种群大周期内有小周期的多谐波叠加特征。小周期波动可使种群的自我稳定性得以维持与延续[26]。
3 讨论和结论
红椿结实有大小年现象,种子数量较大,可以生产相对较大数量的幼苗,并且红椿根部有萌蘖特性,也是其维系种群的基础。红椿是强阳性植物,除幼苗期外,需要强光照才能正常生长。但由于其幼树对光资源的竞争力较弱,很难进入主林层,幼林较高的死亡率引起种群中龄级个体的缺失,导致种群负增长,部分红椿种群甚至出现中龄级个体中断现象[24],因此,推测红椿强阳性的特点与环境中光照资源不足,不仅造成红椿种群在群落中的地位不稳定,也是导致该物种濒危的主要原因。3个红椿种群各龄级的个体数量动态指数都存在3至4次负增长,说明各龄级个体的补充是不连续的,红椿种群处于中间断层的不稳定状态。推测一方面是由于环境筛的抑制导致种内和种间竞争加剧,另一方面是来自环境的干扰使得种群空间生态位重新分配。竹山种群和谷城T2种群受到中等程度干扰,人为干扰程度分别为0.4和0.6,因而这2个种群自然状态下种群个体数量动态指数(Vpi)均低于谷城T1种群(人为干扰程度为0.1)。
长寿命乔木植物的种群动态主要为Deevey Ⅱ型和Ⅲ型存活曲线,Deevey Ⅱ型存活曲线表示种群在生活史各个阶段具有稳定的死亡率;Deevey Ⅲ型为凹曲线,种群早期死亡率高,其后种群数量和死亡率均基本稳定[3]。竹山红椿种群标准化存活曲线近似于Deevey Ⅲ型,而谷城T1和谷城T2种群的标准化存活曲线均为Deevey Ⅱ型,符合长寿命乔木的种群存活曲线特征。种群的死亡率和生存率是种群内在变化与极端环境条件相互作用的结果[26]。3个红椿种群的静态生命表显示:竹山种群第3龄级死亡率最高,其后较为平稳;谷城T1种群第2和第7龄级的死亡率较高;谷城T2种群第7龄级的死亡率最高。红椿种群不同龄级的高死亡率是否会引起存活曲线的变化?其中原因十分复杂,可能与其生理生态学特性及环境因素有关,也可能与特殊条件和极端气候变化(如冻害、地形条件、病虫害等)使生境发生改变有关。竹山种群最高龄级个体数量动态指数为-50.00%,谷城T1和谷城T2种群均为66.67%, 这可能决定了3个红椿种群的存活曲线类型。
种群数量动态既有基波的大格局定位,也有种群自然生长特性和环境因素造成的小周期波动叠加。3个红椿种群的个体损失均集中在第2至第4龄级,这一现象与小周期波动龄级一致,也与红椿生理特性和环境筛矛盾相吻合。由于湖北省内所有红椿天然种群均为次生林,红椿种群周期受调查面积和年龄的限制,谱分析不能完整反映种群的完整生命周期,但3个种群的小周期波动与其生命表特征吻合。推测结构完整的天然红椿种群,大周期内必然存在不同小周期多谐波迭加。
上述研究结果表明:红椿强阳性的特点造成幼龄个体损失继而引起中龄级个体不足,这是导致红椿天然种群濒危的重要原因;从环境变化上看,人为干扰也减少了天然种群的幼苗数量。针对红椿种群的濒危现状,必须开展相应的保护工作。首先,加强人为正向“干扰”,制造“林窗”,改善红椿的生存条件,为种群更新层个体发育创造良好的生境,增强种群天然更新能力;其次,对群落不同林层并用适当疏伐和人工抚育等手段,完善种群分布格局;第三,进行种质资源收集和引种实验,扩大优质人工林规模,使这一濒危优良用材树种资源得到保护和合理开发。
[1]张文辉, 卢彦昌, 周建云, 等. 巴山北坡不同干扰条件下栓皮栎种群结构与动态[J]. 林业科学, 2008, 44(7): 11-16.
[2]胡尔查, 王晓江, 刘永宏, 等. 乌拉山自然保护区油松种群结构与分布格局研究[J]. 林业科学研究, 2011, 24(3): 363-369.
[3]陈远征, 马祥庆, 冯丽贞, 等. 濒危植物沉水樟的种群生命表和谱分析[J]. 生态学报, 2006, 26(12): 4267-4272.
[4]伍业钢, 韩进轩. 阔叶红松林红松种群动态的谱分析[J]. 生态学杂志, 1988, 7(1): 19-23.
[5]刘金福, 洪伟, 吴则焰, 等. 孑遗植物水松(Glyptostrobuspensilis)种群生命表和谱分析[J]. 武汉植物学研究, 2008, 26(3): 259-263.
[6]中国科学院中国植物志编辑委员会. 中国植物志: 第四十三卷第三分册[M]. 北京: 科学出版社, 1997: 36-37.
[7]傅立国. 中国植物红皮书: 稀有濒危植物(第一册)[M]. 北京: 科学出版社, 1991: 142-669.
[8]汪洋, 冉勇军, 王清河, 等. 红椿和毛红椿种植与医药化工研究进展[J]. 湖北林业科技, 2014, 43(6): 57-61.
[9]ZHOU H, CHEN S, FU C, et al. Research progress in cultivation and pharmaceutical chemical ofToonaciliataandToonaciliatavar.pubescens[J].AgriculturalScienceandTechnology,2015,16:722-726, 760.
[10]蒲云海, 张应坤, 江明喜, 等. 神农架北坡堵河源自然保护区植物多样性研究[J]. 武汉植物学研究, 2006, 24(4): 327-332.
[11]高彬, 郭再华, 杜爽爽, 等. 基于GIS的谷城县耕地地力评价[J]. 湖北农业科学, 2013, 52(17): 4099-4102.
[12]蔡秀珠. 人为干扰强度对马尾松人工林生物量及其分配的影响[J]. 中南林学院学报, 2006, 26(2): 51-54.
[13]杨梅, 林思祖, 曹光球. 不同人为干扰强度下甜槠群落物种多样性比较分析[J]. 东北林业大学学报, 2009, 37(7): 30-32.
[14]龙汉利, 冯毅, 向青, 等. 四川盆周山地红椿生长特性研究[J]. 四川林业科技, 2011, 32(3): 37-41, 68.
[15]江洪. 云杉种群生态学[M]. 北京: 中国林业出版社, 1992: 8-26.
[16]丁国民, 刘兴明, 倪自银, 等. 甘肃祁连山青海云杉种群数量动态的初步研究[J].植物资源与环境学报, 2005, 14(4): 36-41.
[17]陈晓德. 植物种群与群落结构动态量化分析方法研究[J]. 生态学报, 1998, 18(2): 214-217.
[18]赵志刚, 郭俊杰, 曾杰, 等. 广西大明山格木种群的空间分布格局与数量动态特征[J]. 林业科学, 2014, 50(10): 1-7.
[19]李妍, 李登武, 韩东辰. 贺兰山东坡青海云杉种群动态[J]. 浙江农林大学学报, 2014, 31(1): 50-56.
[20]周纪纶, 郑师章, 杨持. 植物种群生态学[M]. 北京: 高等教育出版社, 1992: 32-42.
[21]张志祥, 刘鹏, 蔡妙珍, 等. 九龙山珍稀濒危植物南方铁杉种群数量动态[J]. 植物生态学报, 2008, 32(5): 1146-1156.
[22]孙儒勇, 李博, 诸葛阳, 等. 普通生态学[M]. 北京: 高等教育出版社, 1997: 59-66.
[23]申仕康, 马海英, 王跃华, 等. 濒危植物猪血木(EuryodendronexcelsumH. T. Chang)自然种群结构及动态[J]. 生态学报, 2008, 28(5): 2404-2412.
[24]汪洋, 冷艳芝, 苏长江, 等. 恩施天然红椿种群结构及空间分布格局[J]. 浙江农林大学学报, 2016, 33(1): 17-25.
[25]郭连金, 徐卫红. 武夷山米槠种群结构及谱分析[J]. 植物研究, 2007, 27(3): 325-330.
[26]闫淑君, 洪伟, 吴承祯, 等. 丝栗栲种群生命过程及谱分析[J]. 应用与环境生物学报, 2002, 8(4): 351-355.
(责任编辑: 张明霞)
Analysis on natural population dynamics of endangered speciesToonaciliatain northwestern Hubei
WANG Yang1, YAN Kuixing1, TENG Jiaxi2, CHEN Wenxue3, WANG Linbo4, CHEN Song4,①
(1.Landscape and Construction School,Hubei Ecology Vocational College,Wuhan 430200,China; 2. College of Horticulture and Forestry Sciences, Huazhong Agricultural University, Wuhan 430072, China; 3. Zhushan Forestry Bureau, Zhushan 442200, China; 4. Gucheng Forestry Bureau, Gucheng 441799, China),J.PlantResour. &Environ., 2016, 25(3): 96-102
Statistics on three natural populations (Zhushan, Gucheng T1 and Gucheng T2) ofToonaciliataRoem. in northwestern Hubei were carried out by contiguous grid quadrate method, individual quantity dynamic index (Vn) in different age classes, static life table and survival curve of different populations were analyzed by method of diameter class structure of tree replacing age structure of population, and spectrum analysis of three populations was also conducted. The results show that all of three populations ofT.ciliatabelong to growth population, individual quantity dynamic index of Zhushan, Gucheng T1 and Gucheng T2 populations in natural state is 46.27%, 53.46% and 37.71%, respectively. And standardized survival curve of three populations is conformed to be Deevey Ⅲ type, Deevey Ⅱ type and Deevey Ⅱ type, respectively. Mortality of sapling in the 2nd age class 〔2.5 cm≤DBH (diameter at breast height)<7.5 cm〕 of three populations is comparatively high, that of sapling in the 3rd age class (7.5 cm≤DBH<12.5 cm) of Zhushan population is the highest, that of big tree in the 7th age class (27.5 cm≤DBH<32.5 cm) of Gucheng T1 population is the highest, and that of middle aged tree in the 6th age class (22.5 cm≤DBH<27.5 cm) of Gucheng T2 population is the highest. Average life expectancy of individuals in the 1st age class (DBH<2.5 cm) and the 2nd age class of three populations is relatively high. Amplitude of fundamental wave of Zhushan, Gucheng T1 and Gucheng T2 populations is 1.061 6, 1.097 1 and 0.788 2, respectively, and their obvious minor cycles appear in the 4th, 2nd and 4th harmonic waves accordingly, which is indicated that there is periodicity in regeneration of natural population ofT.ciliatain northwestern Hubei with multi-harmonic wave superposition of minor cycle, and fluctuation of population is in accordance withVnvalue of age class. The analysis result of population dynamics shows that due to strong light-demanding physiological feature, natural population ofT.ciliatais under restriction of environmental sieve and experiences environmental disturbance, resulting in loss of sapling and deficiency of middle-aged plants, which is the principal extinctive factor ofT.ciliata. Therefore, some suggestions forT.ciliataare proposed, including intensification of artificial positive “disturbance” to make forest gap, increase of natural regeneration capacity of population, and species introduction and afforestation, to preserve and develop this endangered plant resource.
northwestern Hubei;ToonaciliataRoem.; population dynamics; static life table; survival curve; spectrum analysis
2015-10-08
湖北省科学技术厅公益性研究项目(40 2012DBA40001)
汪洋(1968—),男,湖北武汉人,硕士,副教授,主要从事生态学和植物种质资源的教学与科研工作。
E-mail: 489288208@qq.com
Q948.15; S718.54
A
1674-7895(2016)03-0096-07
10.3969/j.issn.1674-7895.2016.03.12
