暴马丁香不同种源种子和幼苗的表型性状变异及其与地理-气候因子的相关性
2016-11-17杨晓霞冷平生胡增辉刘学娅杨晓红窦德泉
杨晓霞, 冷平生, 郑 健, 胡增辉, 刘学娅, 杨晓红, 窦德泉
(北京农学院园林学院 北京市乡村景观规划设计工程技术研究中心 城乡生态环境北京实验室, 北京 102206)
暴马丁香不同种源种子和幼苗的表型性状变异及其与地理-气候因子的相关性
杨晓霞, 冷平生, 郑健, 胡增辉, 刘学娅, 杨晓红, 窦德泉①
(北京农学院园林学院 北京市乡村景观规划设计工程技术研究中心 城乡生态环境北京实验室, 北京 102206)
为探究不同产地暴马丁香〔Syringareticulatasubsp.amurensis(Rupr.) P. S. Green et M. C. Chang〕种子和幼苗的表型性状变异规律及其与地理-气候因子的关系,以来源于黑龙江、河北、北京、内蒙古、青海和山西的10个种源(包括6个野生种源和4个栽培种源)的种子和幼苗为实验材料,对种子长和宽、种翅长和宽、百粒质量、发芽率、叶长和宽、叶长与叶宽的比值、叶柄长、株高和地径12个表型性状进行测量;在此基础上,进行了方差分析、变异系数分析、主成分分析、聚类分析及相关性分析。结果显示:暴马丁香不同种源间种子和幼苗的表型性状均有极显著差异(P=0.01),差异较大的表型性状为发芽率、百粒质量、种翅长和种翅宽。种源内和种源间各表型性状的变异系数(CV)均差异较大,种源内CV值为1.39%~45.56%,种源间CV值为6.45%~42.76%;其中,种源内种翅长和种翅宽的CV平均值较大,发芽率和百粒质量的CV平均值均较小,而种源间发芽率和百粒质量的CV值则较大。主成分分析结果表明:暴马丁香表型性状间的差异主要体现在种子长、种子宽和叶长,其次为株高、发芽率、地径、叶宽和种翅宽。根据聚类分析结果,供试的10个种源被划分为4类,其中,来源于北京和内蒙古及3个来源于河北的种源聚为Ⅰ类,来源于黑龙江哈尔滨的2个种源聚为Ⅱ类,来源于山西的2个种源聚为Ⅲ类,来源于青海的1个种源单独聚为Ⅳ类,聚类结果与各种源的地理分布有关。相关性分析结果表明:各表型性状间及表型性状与地理-气候因子间均存在不同的相关性,其中,种子发芽率与种子的其他5个表型性状均无显著相关性;经度和纬度与各表型性状均无显著相关性,而海拔、年均温和无霜期则是影响表型性状变异的主要因子。综合分析结果显示:暴马丁香各种源间存在明显的地理遗传分化,地理隔离和生态因子的大幅变化导致其种子和幼苗的表型性状变异丰富。根据研究结果,对暴马丁香的保护和利用提出了一些建议。
暴马丁香; 表型性状; 地理-气候因子; 聚类分析; 相关性分析; 地理变异
暴马丁香〔Syringareticulatasubsp.amurensis(Rupr.) P. S. Green et M. C. Chang〕又名暴马子、白丁香、荷花丁香、中国菩提等,为木犀科(Oleaceae)丁香属(SyringaLinn.)落叶小乔木或落叶大灌木,株高可达10 m;原产中国,是丁香属中分布比较广泛的树种之一[1]。暴马丁香抗逆性强,是良好的园林绿化树种,其树皮、树干及枝条等部位均可药用,综合利用价值较高。在中国,暴马丁香的分布虽然很广,但随市场需求的不断增加,暴马丁香的野生资源遭到滥采乱挖,成片自然分布的群体日益减少[2]。因此,有必要针对暴马丁香野生资源的保护及合理开发利用开展相关研究。目前,对暴马丁香的研究主要集中在种子萌发[3]、栽培管理[4]、园林应用[5]、化学成分[6-7]、药用价值[8-9]和生理特性[10]等方面,但关于其表型性状地理变异的研究较少,对其遗传特性的研究也不全面,不利于对暴马丁香优良种源的保存和遗传改良。
表型是植物各种形态特征的组合,是环境多样性和遗传多样性的综合表现,表型变异在植物适应环境与物种进化上具有重要意义。利用遗传上比较稳定的性状研究植物的表型多样性,不仅可以揭示群体的遗传规律和变异程度,而且有助于评价群体的遗传多样性水平[11-13],这也是遗传育种工作的基础。近年来,对大丽花(DahliapinnataCav.)[14]、白皮松(PinusbungeanaZucc. ex Endl.)[15]、伯乐树(BretschneiderasinensisHemsl.)[16]、无患子(SapindusmukorossiGaertn.)[17]、蜡梅〔Chimonanthuspraecox(Linn.)Link〕[18]、花楸树〔Sorbuspohuashanensis(Hance) Hedl.〕[19]、皂荚(GleditsiasinensisLam.)[20]和杏(ArmeniacavulgarisLam.)[21]等植物的表型变异特征已有较多的研究报道,且研究结果均表明运用表型性状变异的方法能够系统全面地揭示植物各种源间的遗传变异规律。
为揭示暴马丁香表型变异规律及其与地理-气候因子的关系,作者在全面调查暴马丁香在中国分布区域的基础上,选择分布区内有代表性的10个种源为研究对象,对种子和幼苗的主要表型性状进行分析,以期为暴马丁香种质资源的收集和保存、优良种源的选择和遗传改良及良种选育提供基础研究数据。
1 材料和方法
1.1材料
于2013年9月末至10月中旬,在暴马丁香分布区的黑龙江、河北、北京、内蒙古、青海和山西选择10个种源地采集果实,各种源地的自然概况见表1。各种源地均位于中温带或寒温带,其中,来源于北京八达岭国家森林公园(BDL)、河北承德围场(WC)、河北崇礼张麻沟(ZMG)、河北张家口宣化(XH)、黑龙江哈尔滨阿城(AC)和山西吕梁山(LL)的6个种源为野生群体,而来源于黑龙江哈尔滨电塔街(DT)、内蒙古呼和浩特植物园(HO)、山西侯马中条山国家森林公园(ZTS)和青海西宁植物园(XN)的4个种源为栽培群体。在每个种源地中选择20株以上的优良母树采种,母树间距50 m以上,待果实完全干燥后,按照各种源分别收集种子;将每株母树采集的种子等量混合,充分混匀后作为该种源的供试材料。
于2014年4月播种,采用随机区组试验设计,重复3次。播种地位于北京农学院科技园丁香种质资源圃内,地理坐标为东经116°18′、北纬40°05′,海拔43.83 m;属北温带半湿润季风气候区,四季分明,冬季较长、夏季雨量充沛;年平均气温12 ℃、年平均降水量612 mm、年均日照时间2 720 h、无霜期200 d。
表1供试10个暴马丁香种源的产地自然概况
Table 1Natural status of location of ten provenances tested ofSyringareticulatasubsp.amurensis(Rupr.) P. S. Green et M. C. Chang
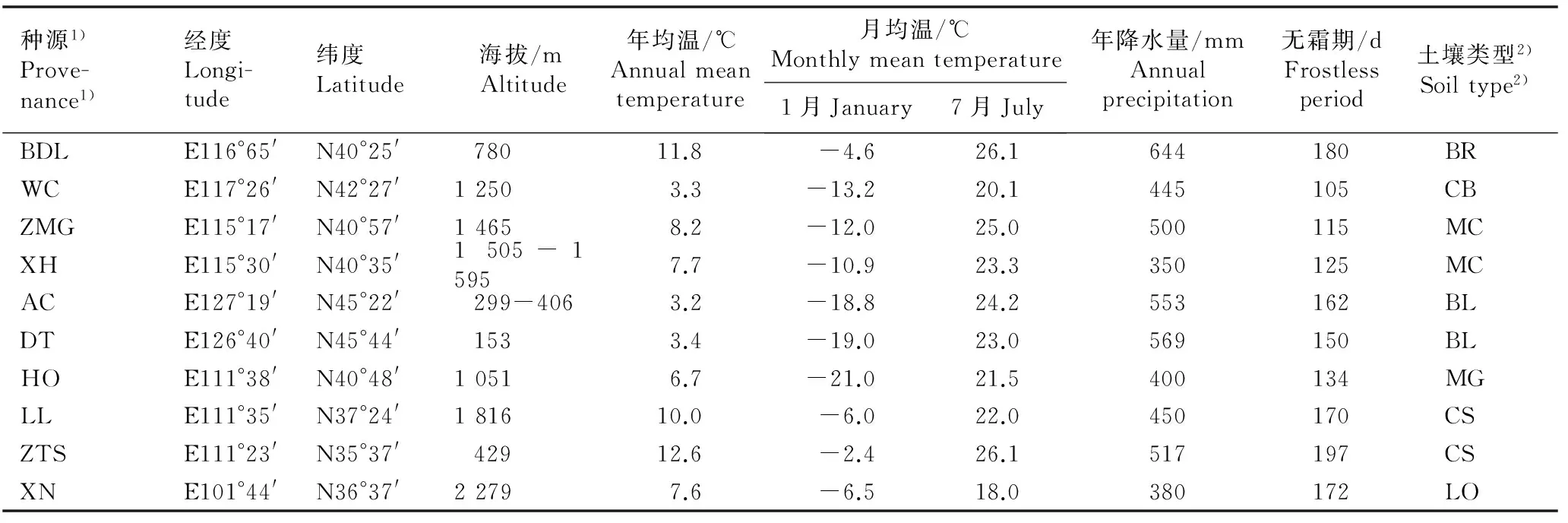
种源1)Prove-nance1)经度Longi-tude纬度Latitude 海拔/m Altitude年均温/℃Annualmeantemperature月均温/℃Monthlymeantemperature1月January7月July年降水量/mmAnnualprecipitation无霜期/dFrostlessperiod土壤类型2)Soiltype2)BDLE116°65'N40°25'78011.8-4.626.1644180 BRWCE117°26'N42°27'12503.3-13.220.1445105 CBZMGE115°17'N40°57'14658.2-12.025.0500115 MCXHE115°30'N40°35'1505-15957.7-10.923.3350125 MCACE127°19'N45°22'299-4063.2-18.824.2553162 BLDTE126°40'N45°44'1533.4-19.023.0569150 BLHOE111°38'N40°48'10516.7-21.021.5400134 MGLLE111°35'N37°24'181610.0-6.022.0450170 CSZTSE111°23'N35°37'42912.6-2.426.1517197 CSXNE101°44'N36°37'22797.6-6.518.0380172 LO
1)BDL: 北京八达岭国家森林公园Badaling National Forest Park of Beijing; WC: 河北承德围场Weichang in Chengde of Hebei; ZMG: 河北崇礼张麻沟Zhangmagou in Chongli of Hebei; XH: 河北张家口宣化Xuanhua in Zhangjiakou of Hebei; AC: 黑龙江哈尔滨阿城Acheng in Harbin of Heilongjiang; DT: 黑龙江哈尔滨电塔街Dianta Street in Harbin of Heilongjiang; HO: 内蒙古呼和浩特植物园Hohhot Botanical Garden in Inner Mongolia; LL: 山西吕梁山Lüliang Mountains in Shanxi; ZTS: 山西侯马中条山国家森林公园Zhongtiaoshan National Forest Park in Houma of Shanxi; XN: 青海西宁植物园Xining Botanical Garden in Qinghai.
2)BR: 棕壤Brown soil; CB: 栗钙土和黑土Chestnut soil and black soil; MC: 山地棕壤和栗钙土Mountain brown soil and chestnut soil; BL: 黑土Black soil; MG: 草甸土和灰褐土Meadow soil and grey cinnamon soil; CS: 褐土Cinnamon soil; LO: 黄土Loess.
1.2方法
1.2.1种子表型性状测量和统计方法各种源随机抽取40粒种子,用直尺(精度0.1 cm)分别测定每粒种子的长和宽,并计算种翅的长和宽;其中,以无种翅时种子两端的长度为种子长,以无种翅时种子中部的宽度为种子宽;种翅长(宽)分别为有种翅时种子长(宽)与无种翅时种子长(宽)的差值。各种源均随机取100粒种子,用电子天平(精度0.01 g)称量质量,重复8次, 取平均值。 按照 《林木种子检验规程》 (GB 2772—1999)的方法测定各种源的种子发芽率。
1.2.2幼苗表型性状测量方法于2014年11月幼苗停止生长后,从各种源中分别随机选取30株幼苗,测量株高和地径,其中,株高为幼苗基部地面处到最高叶面的距离;在每一植株中部选择成熟且完整的叶片3片,分别测定叶长、叶宽和叶柄长,其中,叶长为叶尖与叶柄基部之间的长度,叶宽为叶片中部的宽度,叶柄长为叶柄顶端与叶柄基部之间的长度。上述指标中,地径用游标卡尺(精度0.01 mm)测量,其他指标均用直尺(精度0.1 cm)测量。
1.3数据处理和统计分析
采用EXCEL 2007软件对测定数据进行统计,采用SPSS 19.0软件进行相关性和聚类分析,分析前对数据进行标准化处理。计算不同种源各表型性状的平均值和标准差,并对各表型性状进行表型变异系数和巢式方差分析;以各种源表型性状测定值的平均值为单元,利用样本相关矩阵进行主成分分析,并采用平均连接法进行聚类分析;对各表型性状间及其与各种源地地理-气候因子间的相关性进行分析。
2 结果和分析
2.1暴马丁香不同种源的表型性状比较
对暴马丁香不同种源的种子长、种子宽、种翅长、种翅宽、百粒质量、发芽率、叶长、叶宽、叶长与叶宽的比值(LL/WL)、叶柄长、株高和地径12个表型性状的测定结果见表2。结果表明:暴马丁香不同种源的表型性状分化十分明显,种源间差异较大的性状为发芽率、百粒质量、种翅长和种翅宽。发芽率为种源间变幅最大的性状,其中,产自河北张家口宣化的种子发芽率最高,达到89.35%;而产自青海西宁植物园的种子发芽率最低,仅10.21%。种子百粒质量在种源间的变幅也较大,其中,产自黑龙江哈尔滨电塔街的种子百粒质量最大,为2.44 g;产自河北崇礼张麻沟的种子百粒质量最小,仅0.97 g。从种翅性状看,产自黑龙江哈尔滨阿城的种子种翅长度最长(0.54 cm),产自北京八达岭国家森林公园的种子种翅长度最短(0.27 cm);产自黑龙江哈尔滨阿城的种子种翅宽度最大(0.24 cm),而产自山西侯马中条山国家公园的种子种翅宽度最小(0.12 cm)。除前述的4个性状外,其余8个性状的变幅相对较小。

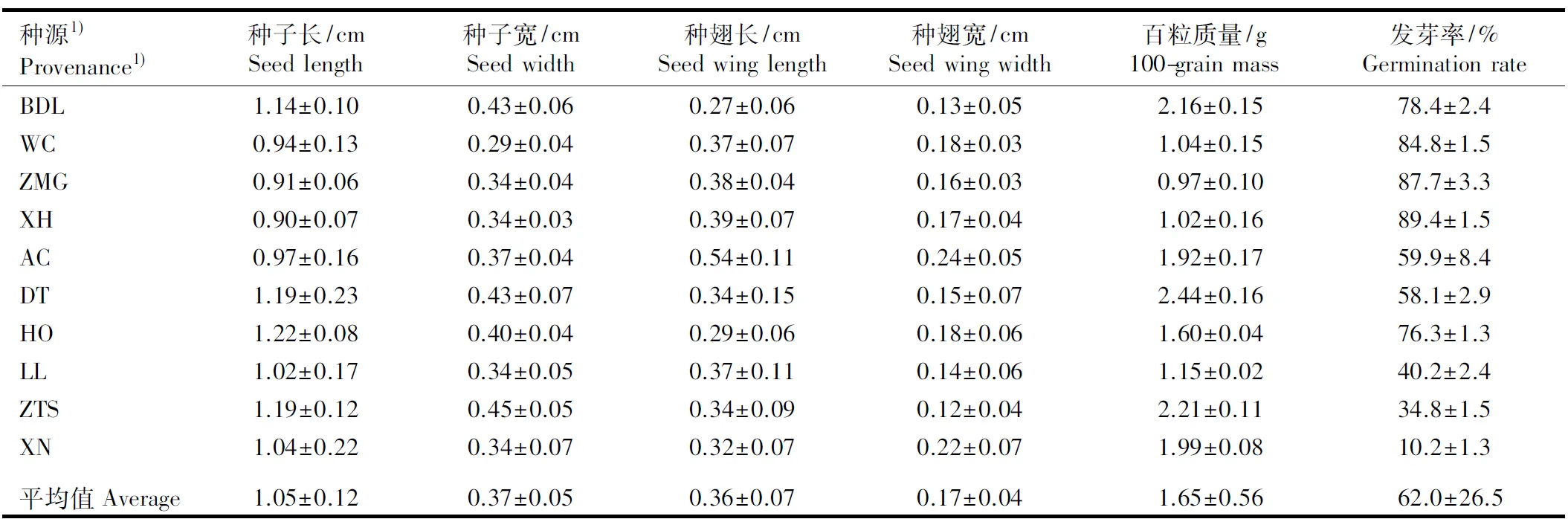
种源1)Provenance1)种子长/cmSeedlength种子宽/cmSeedwidth种翅长/cmSeedwinglength种翅宽/cmSeedwingwidth百粒质量/g100-grainmass发芽率/%GerminationrateBDL1.14±0.100.43±0.060.27±0.060.13±0.05 2.16±0.1578.4±2.4WC0.94±0.130.29±0.040.37±0.070.18±0.031.04±0.1584.8±1.5ZMG0.91±0.060.34±0.040.38±0.040.16±0.030.97±0.1087.7±3.3XH0.90±0.070.34±0.030.39±0.070.17±0.041.02±0.1689.4±1.5AC0.97±0.160.37±0.040.54±0.110.24±0.051.92±0.1759.9±8.4DT1.19±0.230.43±0.070.34±0.150.15±0.072.44±0.1658.1±2.9HO1.22±0.080.40±0.040.29±0.060.18±0.061.60±0.0476.3±1.3LL1.02±0.170.34±0.050.37±0.110.14±0.061.15±0.0240.2±2.4ZTS1.19±0.120.45±0.050.34±0.090.12±0.042.21±0.1134.8±1.5XN1.04±0.220.34±0.070.32±0.070.22±0.071.99±0.0810.2±1.3平均值Aver-age1.05±0.120.37±0.050.36±0.070.17±0.041.65±0.5662.0±26.5
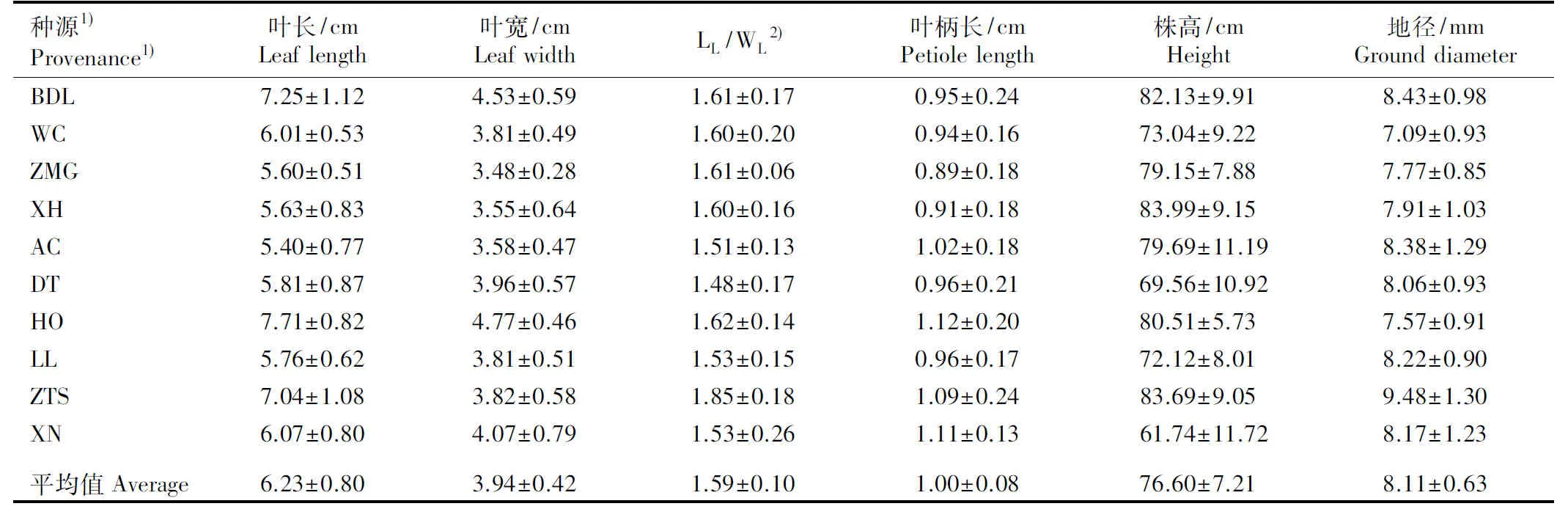
种源1)Provenance1)叶长/cmLeaflength叶宽/cmLeafwidthLL/WL2)叶柄长/cmPetiolelength株高/cmHeight地径/mmGrounddiameterBDL7.25±1.124.53±0.591.61±0.170.95±0.2482.13±9.918.43±0.98WC6.01±0.533.81±0.491.60±0.200.94±0.1673.04±9.227.09±0.93ZMG5.60±0.513.48±0.281.61±0.060.89±0.1879.15±7.887.77±0.85XH5.63±0.833.55±0.641.60±0.160.91±0.1883.99±9.157.91±1.03AC5.40±0.773.58±0.471.51±0.131.02±0.1879.69±11.198.38±1.29DT5.81±0.873.96±0.571.48±0.170.96±0.2169.56±10.928.06±0.93HO7.71±0.824.77±0.461.62±0.141.12±0.2080.51±5.737.57±0.91LL5.76±0.623.81±0.511.53±0.150.96±0.1772.12±8.018.22±0.90ZTS7.04±1.083.82±0.581.85±0.181.09±0.2483.69±9.059.48±1.30XN6.07±0.804.07±0.791.53±0.261.11±0.1361.74±11.728.17±1.23平均值Aver-age6.23±0.803.94±0.421.59±0.101.00±0.0876.60±7.218.11±0.63
1)BDL: 北京八达岭国家森林公园Badaling National Forest Park of Beijing; WC: 河北承德围场Weichang in Chengde of Hebei; ZMG: 河北崇礼张麻沟Zhangmagou in Chongli of Hebei; XH: 河北张家口宣化Xuanhua in Zhangjiakou of Hebei; AC: 黑龙江哈尔滨阿城Acheng in Harbin of Heilongjiang; DT: 黑龙江哈尔滨电塔街Dianta Street in Harbin of Heilongjiang; HO: 内蒙古呼和浩特植物园Hohhot Botanical Garden in Inner Mongolia; LL: 山西吕梁山Lüliang Mountains in Shanxi; ZTS: 山西侯马中条山国家森林公园Zhongtiaoshan National Forest Park in Houma of Shanxi; XN: 青海西宁植物园Xining Botanical Garden in Qinghai.
2)LL/WL: 叶长与叶宽的比值Ratio of leaf length to leaf width.
采用巢式方差分析对10个种源间各表型性状的差异进行显著性分析,结果(表3)表明:不同种源间12个表型性状的差异均达到极显著水平(P=0.01),说明暴马丁香种源间种子和幼苗的各项表型性状均存在丰富的变异,种源间的遗传分化显著,优良种源的选择潜力很大。
表3暴马丁香不同种源的种子和幼苗表型性状的方差分析结果
Table 3Variance analysis result on phenotypic traits of seed and seedling ofSyringareticulatasubsp.amurensis(Rupr.) P. S. Green et M. C. Chang from different provenances
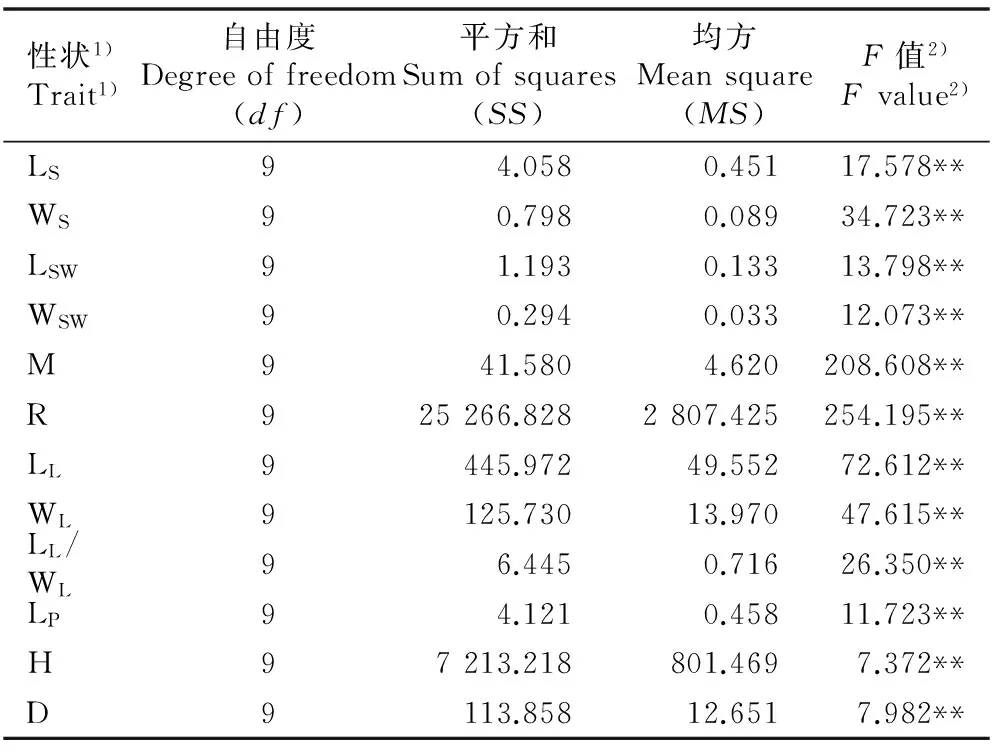
性状1)Trait1)自由度Degreeoffreedom(df)平方和Sumofsquares(SS)均方Meansquare(MS)F值2)Fvalue2)LS9 4.058 0.45117.578**WS9 0.7980.08934.723**LSW9 1.1930.13313.798**WSW9 0.2940.03312.073**M941.580 4.620208.608**R9 25266.8282807.425254.195**LL9 445.97249.55272.612**WL9 125.73013.97047.615**LL/WL9 6.4450.71626.350**LP9 4.1210.45811.723**H9 7213.218801.4697.372**D9 113.85812.6517.982**
1)LS: 种子长 Seed length; WS: 种子宽 Seed width; LSW: 种翅长 Seed wing length; WSW: 种翅宽 Seed wing width; M: 百粒质量 100-grain mass; R: 发芽率 Germination rate; LL: 叶长 Leaf length; WL: 叶宽Leaf width; LL/WL: 叶长与叶宽的比值Ratio of leaf length to leaf width; LP: 叶柄长Petiole length; H: 株高 Height; D: 地径 Ground diameter.
2)**:P=0.01.
2.2暴马丁香不同种源种子和幼苗表型性状的变异系数比较
暴马丁香不同种源种子和幼苗12个表型性状的变异系数(CV)见表4。
由表4可以看出:12个表型性状的CV平均值在种源内存在明显差异,12个表型性状按种源内CV平均值由大到小依次排序为种翅宽、种翅长、叶柄长、叶宽、种子长、地径、叶长、种子宽、株高、叶长与叶宽的比值(LL/WL)、百粒质量、发芽率。其中,种翅宽和种翅长的种源内CV平均值较大, 分别为28.80%和 23.12%;百粒质量和发芽率的种源内CV平均值较小,分别为7.55%和5.39%,说明同一种源内百粒质量和发芽率是较稳定的表型性状,而种翅性状在同一种源内变异较大。
由各表型性状的种源间CV值可见:暴马丁香种子的表型性状的种源间CV值总体上高于幼苗的表型性状,发芽率和百粒质量的种源间CV值较大,分别为42.76%和34.23%,LL/WL的种源间CV值最小,仅为6.45%,说明暴马丁香不同种源间种子发芽率和百粒质量有明显差异。
由表4还可以看出:来源于黑龙江哈尔滨电塔街和青海西宁植物园的2个栽培种源的表型性状CV平均值大于6个野生种源,说明这2个栽培种源表型性状的变异丰富度优于野生种源;但来源于内蒙古呼和浩特植物园和青海西宁植物园的2个栽培种源的表型性状CV平均值与6个野生种源无明显差异。另外,在暴马丁香种子性状中,不论种源间还是种源内,种子宽和种翅宽的CV值均分别大于种子长和种翅长,说明相较于种子长和种翅长,种子宽和种翅宽更易受环境因子的影响。
2.3暴马丁香不同种源种子和幼苗表型性状的主成分分析
对暴马丁香种子长(x1)、种子宽(x2)、种翅长(x3)、种翅宽(x4)、百粒质量(x5)、发芽率(x6)、叶长(x7)、叶宽(x8)、叶长与叶宽的比值(x9)、叶柄长(x10)、株高(x11)和地径(x12)12个表型性状进行主成分分析,结果见表5。由表5可知:第1主成分(y1)的贡献率最高,为42.807%,第2主成分(y2)的贡献率也达到20.832%,第3主成分(y3)和第4主成分(y4)的贡献率分别为16.129%和8.893%,前4个主成分的累计贡献率达88.661%,高于“累计贡献率≥85%[22]”的标准,故选取第1、第2、第3和第4主成分即可。
根据前4个主成分中各表型性状的特征向量,推导出第1主成分相关方程为y1=0.937x1+0.858x2-0.639x3- 0.460x4+ 0.727x5- 0.353x6+ 0.850x7+ 0.713x8+0.506x9+0.634x10+0.153x11+0.581x12;第2主成分相关方程为y2=-0.108x1+0.091x2-0.191x3-0.624x4- 0.425x5+ 0.777x6+ 0.263x7- 0.030x8+ 0.511x9-0.530x10+0.807x11-0.084x12;第3主成分相关方程为y3=-0.130x1+0.286x2+0.560x3-0.096x4+ 0.229x5- 0.289x6- 0.341x7- 0.647x8+ 0.374x9- 0.025x10+0.323x11+0.773x12;第4主成分相关方程为y4=-0.070x1-0.104x2+0.339x3+0.577x4-0.128x5+ 0.150x6+ 0.268x7+0.143x8+0.246x9+0.497x10+ 0.398x11-0.084x12。第1主成分中,种子长、种子宽和叶长的特征向量相对较大,表明第1主成分是种子长、种子宽和叶长的综合因子;第2主成分中,株高和发芽率的特征向量相对较大,表明第2主成分主要是株高和发芽率的综合因子;第3主成分中,地径和叶宽的特征向量相对较大;第4主成分中,种翅宽的特征向量相对较大。因此,暴马丁香不同种源间种子和幼苗表型性状的差异主要表现在种子长、种子宽和叶长,其次是株高、发芽率、地径、叶宽和种翅宽。
表4暴马丁香不同种源的种子和幼苗表型性状的变异系数
Table 4Coefficient of variation of phenotypic traits of seed and seedling ofSyringareticulatasubsp.amurensis(Rupr.) P. S. Green et M. C. Chang from different provenances
1)BDL: 北京八达岭国家森林公园Badaling National Forest Park of Beijing; WC: 河北承德围场Weichang in Chengde of Hebei; ZMG: 河北崇礼张麻沟Zhangmagou in Chongli of Hebei; XH: 河北张家口宣化Xuanhua in Zhangjiakou of Hebei; AC: 黑龙江哈尔滨阿城Acheng in Harbin of Heilongjiang; DT: 黑龙江哈尔滨电塔街Dianta Street in Harbin of Heilongjiang; HO: 内蒙古呼和浩特植物园Hohhot Botanical Garden in Inner Mongolia; LL: 山西吕梁山Lüliang Mountains in Shanxi; ZTS: 山西侯马中条山国家森林公园Zhongtiaoshan National Forest Park in Houma of Shanxi; XN: 青海西宁植物园Xining Botanical Garden in Qinghai.
2)LS: 种子长 Seed length; WS: 种子宽 Seed width; LSW: 种翅长 Seed wing length; WSW: 种翅宽 Seed wing width; M: 百粒质量 100-grain mass; R: 发芽率 Germination rate; LL: 叶长 Leaf length; WL: 叶宽Leaf width; LL/WL: 叶长与叶宽的比值 Ratio of leaf length to leaf width; LP: 叶柄长Petiole length; H: 株高 Height; D: 地径 Ground diameter.
表5暴马丁香不同种源种子和幼苗12个表型性状的主成分分析结果1)
Table 5Result of principal component analysis of twelve phenotypic traits of seed and seedling ofSyringareticulatasubsp.amurensis(Rupr.) P. S. Green et M. C. Chang from different provenances1)
1)x1: 种子长Seed length;x2: 种子宽Seed width;x3: 种翅长Seed wing length;x4: 种翅宽Seed wing width;x5: 百粒质量100-grain mass;x6: 发芽率Germination rate;x7: 叶长Leaf length;x8: 叶宽Leaf width;x9: 叶长与叶宽的比值Ratio of leaf length to leaf width;x10: 叶柄长Petiole length;x11: 株高Height;x12: 地径Ground diameter. E: 特征值Eigenvalue; CCR: 累计贡献率Cumulative contribution rate.
2.4暴马丁香不同种源的聚类分析
采用SPSS 19.0统计分析软件,基于种子和幼苗的表型性状数据对暴马丁香不同种源进行Q型聚类分析,结果见图1。由图1可以看出:在欧氏距离为7处,10个种源被划分为4大类。其中,北京八达岭国家森林公园(BDL)、内蒙古呼和浩特植物园(HO)以及河北的崇礼张麻沟(ZMG)、张家口宣化(XH)和承德围场(WC)5个种源聚为Ⅰ类;黑龙江哈尔滨阿城(AC)和黑龙江哈尔滨电塔街(DT)2个种源聚为Ⅱ类;山西吕梁山(LL)和山西侯马中条山国家森林公园(ZTS)2个种源聚为Ⅲ类;青海西宁植物园(XN)种源单独聚为Ⅳ类。另外,Ⅰ类的5个种源又进一步被分为2个亚类, 其中,BDL和HO 2个种源聚为一个亚类,ZMG、XH和WC 3个种源聚为另一个亚类。综合分析结果表明:10个种源的聚类结果与各种源的地理分布有关。
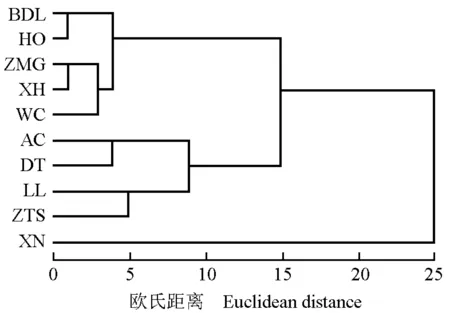
BDL: 北京八达岭国家森林公园Badaling National Forest Park of Beijing; HO: 内蒙古呼和浩特植物园Hohhot Botanical Garden in Inner Mongolia; ZMG: 河北崇礼张麻沟Zhangmagou in Chongli of Hebei; XH: 河北张家口宣化Xuanhua in Zhangjiakou of Hebei; WC: 河北承德围场Weichang in Chengde of Hebei; AC: 黑龙江哈尔滨阿城Acheng in Harbin of Heilongjiang; DT: 黑龙江哈尔滨电塔街Dianta Street in Harbin of Heilongjiang; LL: 山西吕梁山Lüliang Mountains in Shanxi; ZTS: 山西侯马中条山国家森林公园Zhongtiaoshan National Forest Park in Houma of Shanxi; XN: 青海西宁植物园Xining Botanical Garden in Qinghai.
图1基于种子和幼苗表型性状的暴马丁香10个种源的聚类图
Fig. 1Cluster diagram of ten provenances ofSyringareticulatasubsp.amurensis(Rupr.) P. S. Green et M. C. Chang based on phenotypic traits of seed and seedling
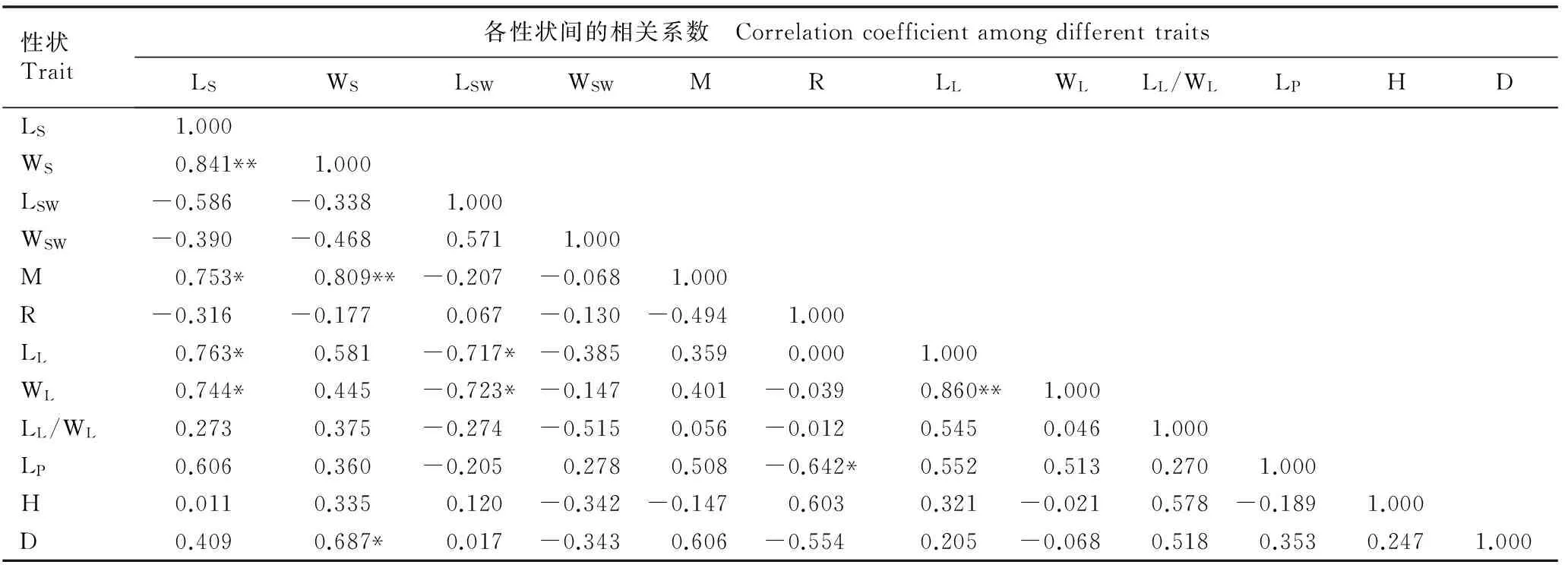
2.5暴马丁香种子和幼苗表型性状间的相关性分析
对暴马丁香种子和幼苗12个表型性状的平均值进行相关性分析,结果见表6。由表6可见:暴马丁香的种子长与种子宽呈极显著正相关,与百粒质量、叶长和叶宽均呈显著正相关,相关系数分别为0.841、0.753、0.763和0.744;种子宽与百粒质量和地径分别呈极显著和显著正相关, 相关系数分别为0.809和
0.687;种翅长与叶长和叶宽均呈显著负相关,相关系数分别为-0.717和-0.723;发芽率与叶柄长呈显著负相关,相关系数为-0.642;叶长与叶宽呈极显著正相关,相关系数为0.860。另外,其种子发芽率与种子的其他5个表型性状均无显著相关性,种翅宽、叶片长与叶片宽的比值和幼苗株高与其他表型性状也均无显著相关性。
2.6暴马丁香种子和幼苗的表型性状与种源地地理-气候因子间的相关性分析
暴马丁香种子和幼苗的表型性状与种源地地理-气候因子间的相关系数见表7。结果显示:种子宽与海拔和无霜期分别呈显著负相关和显著正相关,说明产地的无霜期越长、海拔越低,种子越宽。种翅宽与年均温呈显著负相关,叶长与叶宽的比值与年均温呈显著正相关,说明产地的年均温越高,种翅宽越小、叶长与叶宽的比值越大(即叶片越长)。地径与无霜期呈极显著正相关,百粒质量与无霜期呈显著正相关,而发芽率与无霜期呈显著负相关,说明产地的无霜期越长,幼苗的地径越粗、种子百粒质量越大,而种子发芽率则越低。总体上看,不同产地的地理因子(经度、纬度和海拔)与种子长、叶长、叶宽、叶长与叶宽的比值、叶柄长和地径均呈负相关,与种翅宽均呈正相关;而不同产地的气候因子(年均温、年降水量和无霜期)与种子长、种子宽、百粒质量、叶长、叶宽、叶长与叶宽的比值和地径均呈正相关,与种翅宽均呈负相关。
表6暴马丁香种子和幼苗各表型性状间的相关系数1)
Table 6Correlation coefficient among phenotypic traits of seed and seedling ofSyringareticulatasubsp.amurensis(Rupr.) P. S. Green et M. C. Chang1)
性状Trait各性状间的相关系数 CorrelationcoefficientamongdifferenttraitsLSWSLSWWSWMRLLWLLL/WLLPHDLS1.000WS0.841**1.000LSW-0.586-0.3381.000WSW-0.390-0.4680.5711.000M0.753*0.809**-0.207-0.0681.000R-0.316-0.1770.067-0.130-0.4941.000LL0.763*0.581-0.717*-0.3850.3590.0001.000WL0.744*0.445-0.723*-0.1470.401-0.0390.860**1.000LL/WL0.2730.375-0.274-0.5150.056-0.0120.5450.0461.000LP0.6060.360-0.2050.2780.508-0.642*0.5520.5130.2701.000H0.0110.3350.120-0.342-0.1470.6030.321-0.0210.578-0.1891.000D0.4090.687*0.017-0.3430.606-0.5540.205-0.0680.5180.3530.2471.000
1)LS: 种子长 Seed length; WS: 种子宽 Seed width; LSW: 种翅长 Seed wing length; WSW: 种翅宽 Seed wing width; M: 百粒质量 100-grain mass; R: 发芽率 Germination rate; LL: 叶长 Leaf length; WL: 叶宽Leaf width; LL/WL: 叶长与叶宽的比值 Ratio of leaf length to leaf width; LP: 叶柄长Petiole length; H: 株高 Height; D: 地径 Ground diameter. *:P=0.05; **:P=0.01.
表7暴马丁香种子和幼苗的表型性状与种源地地理-气候因子间的相关系数
Table 7Correlation coefficient of phenotypic traits of seed and seedling ofSyringareticulatasubsp.amurensis(Rupr.) P. S. Green et M. C. Chang with geographic-climatic factors of provenance locations
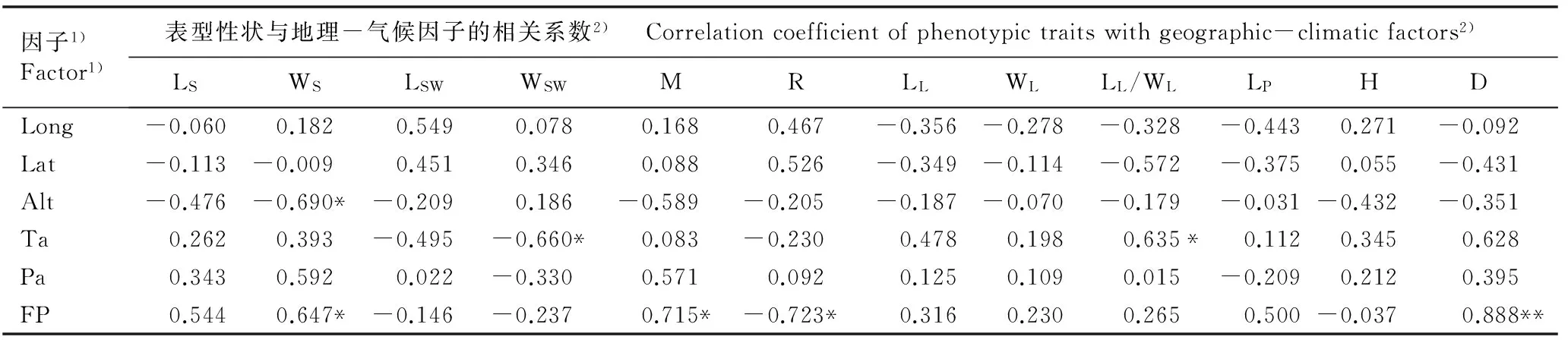
因子1)Factor1)表型性状与地理-气候因子的相关系数2) Correlationcoefficientofphenotypictraitswithgeographic-climaticfactors2)LSWSLSWWSWMRLLWLLL/WLLPHD Long-0.0600.1820.5490.0780.1680.467-0.356-0.278-0.328-0.4430.271-0.092 Lat-0.113-0.0090.4510.3460.0880.526-0.349-0.114-0.572-0.3750.055-0.431Alt-0.476-0.690*-0.2090.186-0.589-0.205-0.187-0.070-0.179-0.031-0.432-0.351Ta0.2620.393-0.495-0.660*0.083-0.2300.4780.1980.635*0.1120.3450.628Pa0.3430.5920.022-0.3300.5710.0920.1250.1090.015-0.2090.2120.395FP0.5440.647*-0.146-0.2370.715*-0.723*0.3160.2300.2650.500-0.0370.888**
1)Long: 经度 Longitude; Lat: 纬度 Latitude; Alt: 海拔 Altitude; Ta: 年均温 Annual mean temperature; Pa: 年降水量 Annual precipitation; FP: 无霜期 Frostless period.
2)LS: 种子长 Seed length; WS: 种子宽 Seed width; LSW: 种翅长 Seed wing length; WSW: 种翅宽 Seed wing width; M: 百粒质量 100-grain mass; R: 发芽率 Germination rate; LL: 叶长 Leaf length; WL: 叶宽Leaf width; LL/WL: 叶长与叶宽的比值 Ratio of leaf length to leaf width; LP: 叶柄长Petiole length; H: 株高 Height; D: 地径 Ground diameter. *:P=0.05; **:P=0.01.
3 讨 论
3.1暴马丁香不同种源种子和幼苗表型性状的变异特征分析
形态性状变异是遗传变异的重要构成部分,形态变异越大,遗传变异的可能性就越大。对暴马丁香10个种源的种子长、种子宽、种翅长、种翅宽、百粒质量、发芽率、叶长、叶宽、叶长与叶宽的比值、叶柄长、株高和地径的测定结果表明:暴马丁香种子和幼苗的表型性状变异丰富,种源间12个表型性状的差异均达到极显著水平,说明暴马丁香种源间存在明显的地理遗传分化,具有优良种源的选择潜力。该结果与花楸树[19]、浙江楠(PhoebechekiangensisC. B. Shang)[23]、木荷(SchimasuperbaGardn. et Champ.)[24]和短枝木麻黄(Casuarinaequisetifoliasubsp.equisetifoliaL. Johnson)[25]等树种的研究结果相似。从表型性状的变异系数(CV)看,暴马丁香种源内种子和幼苗的表型性状变异丰富,而种源间各表型性状的CV值为6.45%~42.76%,差异极显著;其中,种源内种翅宽的CV平均值最大(28.80%),种源内发芽率的CV平均值最小(5.39%),说明在种源内发芽率稳定性最高;种源间,发芽率的CV值最大(42.76%),叶长与叶宽的比值的CV值最小(6.45%),与暴马丁香对环境变化的敏感性有关。在暴马丁香的种子表型性状中,种子宽和种翅宽的CV值分别大于种子长和种翅长,说明暴马丁香种子的横向生长变异程度大于纵向生长,与王娅丽等[26]对青海云杉(PiceacrassifoliaKom.)的观测分析结果一致。研究结果表明:供试的暴马丁香10个种源种子和幼苗的12个表型性状在种源间和种源内均存在丰富的变异,即不同种源的环境异质性导致群体表型变异的差异,这些变异在受到生境条件影响的同时也反映出种质的遗传多样性特点,因此,暴马丁香优良种源的选择潜力很大,有利于其新品种的选育和种质创新。
暴马丁香地理种源间种子和幼苗的表型性状分化显著,可能与其分布范围广、分布区内环境因素多样有关。一般而言,植物的分布范围广则变异较大,分布范围窄则变异较小;分布区的环境条件越复杂,种源内群体的遗传变异就越大。种源间变异能够反映植物在地理生殖隔离上的变异,是种内多样性的重要组成部分[27];而种源内变异则能反映植物对不同环境的适应状况,种源内变异越大则植物的适应范围越广。暴马丁香广泛分布于中国北部地区,本研究选择的10个地理种源分布区跨度较大,主要分布于暖温带至北温带的大部分地区,气候条件差异较大,年均温的变幅从3.2 ℃(黑龙江哈尔滨阿城)到12.6 ℃(山西侯马中条山国家森林公园),地理隔离和生态因子的大幅变化造就了表型性状的较大变异。
聚类分析结果表明:依据种子和幼苗的表型性状对10个暴马丁香种源进行聚类分析,聚类结果与各种源的地理分布相关。北京八达岭国家森林公园与河北的崇礼张麻沟、张家口宣化、承德围场的地理位置相对较近,环境条件比较一致,而种植于内蒙古呼和浩特植物园的暴马丁香栽培种源的表型性状与这4个种源差异较小,因而这5个种源聚为Ⅰ类;来源于黑龙江哈尔滨电塔街的栽培种源与来源于黑龙江哈尔滨阿城的野生种源聚为Ⅱ类,来源于山西侯马中条山国家森林公园的栽培种源与来源于山西吕梁山的野生种源聚为Ⅲ类,这2类均包含栽培种源和野生种源,栽培种源尽管来源不明,但由于移栽时间较长,在相似的生态环境长期作用下,表型性状与地理分布相近的野生种源趋于相似,因而聚在一起;而来源于青海西宁植物园的种源与其他种源的地理位置相距最远,生态条件差异最大,所以单独聚为Ⅳ类。
3.2暴马丁香不同种源种子和幼苗的表型性状与地理-气候因子间的关系分析
从暴马丁香种子和幼苗表型性状与地理-气候因子间的相关性看,种子宽与海拔和无霜期分别呈显著负相关和显著正相关;年均温与种翅宽和叶长与叶宽的比值分别呈显著负相关和显著正相关;无霜期与地径呈极显著正相关,与百粒质量和发芽率分别呈显著正相关和显著负相关,说明无霜期、年均温和海拔是影响暴马丁香不同种源种子和幼苗表型性状的主要因子。幼苗的叶长、叶宽、株高和地径均与年均温和年降水量呈正相关,说明年均温高、年降水量大有利于暴马丁香幼苗的生长。但各表型性状与产地的经度和纬度的相关性均不显著,说明暴马丁香种子和幼苗的表型性状无明显的经纬度地理变异模式,这一现象也存在于同属植物巧玲花(SyringapubescensTurcz.)[28]中,但与乳源木莲(ManglietiayuyuanensisLaw)[29]和毛红椿〔Toonaciliatavar.pubescens(Franch.) Hand.-Mazz.〕[30]等其他科属植物的研究结果不一致,推测这一现象为丁香属植物的共性。因此,在进行暴马丁香优良种源选择时不能只考虑产地的经度和纬度,应将产地的海拔和温度等也作为选择因子。
3.3对暴马丁香保护和合理利用的建议
暴马丁香原产中国,为珍贵的用材树种,经过长期的地理隔离、遗传突变、自然选择等过程的共同作用,产生了丰富的表型变异,且表型变异的幅度越大,遗传变异的可能性就越大。目前,暴马丁香的生存生境比较脆弱,自然更新困难,加之人们对其生境的破坏以及砍伐和采挖利用,其天然林受到破坏,种群数量减少。据此,对暴马丁香的保护和合理利用提出如下建议:1)加强对暴马丁香现有天然林的保护,保护其生境条件并严禁人为砍伐和采挖,促进其野生群体的天然更新;2)开展暴马丁香遗传多样性研究,并结合表型性状变异特征分析其遗传结构,制定合理的保护策略;3)同时开展原地保存和异地保存,构建种子库,营建暴马丁香优良种质资源圃;4)筛选优良种质和变异植株,并进行人工驯化和栽培,以此满足其人工林发展的需求,避免对现有自然资源的破坏。
综上所述,暴马丁香不同种源的种子和幼苗的表型性状存在丰富的变异,优良种源的选择潜力很大,且产地的海拔、年均温和无霜期是影响暴马丁香表型性状的主要地理-气候因子,可作为暴马丁香引种、栽培、保护和利用的科学依据。
[1]郑万钧. 中国树木志: 第四卷[M]. 北京: 中国林业出版社, 2004: 4422-4423.
[2]杨兆静, 张钦弟, 李豪, 等. 山西五鹿山自然保护区暴马丁香群落木本植物种间联结性分析[J]. 植物科学学报, 2013, 31(1): 42-48.
[3]张芹, 徐学山, 李保会, 等. 不同处理条件对暴马丁香种子萌发的影响[J]. 北方园艺, 2014(1): 54-57.
[4]任光宇, 黄婕, 穆蕾. 暴马丁香育苗技术及管理措施[J]. 安徽农学通报, 2012, 18(16): 171-172.
[5]马辉. 丁香的美学特征及其在园林中的应用[J]. 安徽农业科学, 2010, 38(14): 7641-7643, 7676.
[6]BI X, LI W, SASAKI T, et al. Secoiridoid glucosides and related compounds fromSyringareticulataand their antioxidant activities[J]. Bioorganic and Medicinal Chemistry Letters, 2011, 21: 6426-6429.
[7]王丽华, 张生, 王少明, 等. 暴马丁香中桦木酸的鉴定及质量分数测定[J]. 东北林业大学学报, 2013, 41(8): 144-145, 159.
[8]张崇禧, 蔡恩博, 陈艳, 等. 暴马丁香抑菌活性、抗氧化作用研究及微量元素的测定[J]. 食品科学, 2010, 31(7): 131-134.
[9]宋光西, 魏锋, 马双成. 高效液相色谱法测定暴马子皮药材中2个成分的含量[J]. 药物分析杂志, 2012, 32(10): 1858-1860, 1869.
[10]马艳丽, 王鹏. 低温胁迫对2种丁香属植物抗寒生理指标的影响[J]. 河北农业大学学报, 2013, 36(2): 54-59.
[11]GANATSASP,TSAKALDIMIM,THANOSC.Seedandcone diversity and seed germination ofPinuspineain Strofylia Site of the Natura 2000 Network[J]. Biodiversity and Conservation, 2008, 17: 2427-2439.
[12]ANISZEWSKI T, KUPARI M H, LEINONEN A J. Seed number, seed size and diversity in Washington Lupin (LupinuspolyphyllusLindl.)[J]. Annals of Botany, 2001, 87: 77-82.
[13]高秀琴, 兰进好, 穆平, 等. 小麦遗传多样性研究进展[J]. 山东农业科学, 2007(3): 33-36.
[14]冯立娟, 苑兆和, 尹燕雷, 等. 大丽花花型群体表型性状遗传多样性研究[J]. 山东农业科学, 2010(7): 12-16.
[15]李斌, 顾万春, 卢宝明. 白皮松天然群体种实性状表型多样性研究[J]. 生物多样性, 2002, 10(2): 181-188.
[16]张季, 田华林, 王玉奇, 等. 3个不同地理种源的伯乐树种子和苗期生长差异比较[J]. 热带农业科学, 2011, 31(5): 12-15.
[17]刁松锋, 邵文豪, 姜景民, 等. 基于种实性状的无患子天然群体表型多样性研究[J]. 生态学报, 2014, 34(6): 1451-1460.
[18]赵冰, 张启翔. 蜡梅种质资源表型多样性[J]. 东北林业大学学报, 2007, 35(5): 10-13, 35.
[19]郑健, 胡增辉, 郑勇奇, 等. 花楸树种源间表型性状的地理变异分析[J]. 植物资源与环境学报, 2012, 21(3): 50-56.
[20]李伟, 林富荣, 郑勇奇, 等. 皂荚南方天然群体种实表型多样性[J]. 植物生态学报, 2013, 37(1): 61-69.
[21]刘娟, 廖康, 刘欢, 等. 新疆野杏种质资源表型性状多样性研究[J]. 西北植物学报, 2015, 35(5): 1021-1030.
[22]张力. SPSS 19.0(中文版)在生物统计中的应用[M]. 3版. 厦门: 厦门大学出版社, 2013.
[23]李因刚, 柳新红, 马俊伟, 等. 浙江楠种群表型变异[J]. 植物生态学报, 2014, 38(12): 1315-1324.
[24]王秀花, 陈柳英, 马丽珍, 等. 7年生木荷生长和木材基本密度地理遗传变异及种源选择[J]. 林业科学研究, 2011, 24(3): 307-313.
[25]胡盼, 仲崇禄, 张勇, 等. 短枝木麻黄种群苗期表型多样性评价[J]. 西北植物学报, 2015, 35(5): 1013-1020.
[26]王娅丽, 李毅. 祁连山青海云杉天然群体的种实性状表型多样性[J]. 植物生态学报, 2008, 32(2): 355-362.
[28]龚梅香. 巧玲花天然群体遗传多样性研究[D]. 北京: 北京林业大学园林学院, 2007.
[29]李因刚, 周志春, 金国庆, 等. 乳源木莲苗生长和形态的地理种源分化[J]. 林业科学研究, 2007, 20(1): 35-39.
[30]刘军, 张海燕, 姜景民, 等. 毛红椿种实和苗期生长性状地理种源变异[J]. 南京林业大学学报(自然科学版), 2011, 35(3): 55-59.
(责任编辑: 张明霞)
Variation of phenotypic traits of seed and seedling ofSyringareticulatasubsp.amurensisfrom different provenances and their correlations with geographic-climatic factors
YANG Xiaoxia, LENG Pingsheng, ZHENG Jian, HU Zenghui, LIU Xueya, YANG Xiaohong, DOU Dequan①
(College of Landscape Architecture, Beijing Engineering Research Center of Rural Landscape Planning and Design, Beijing Laboratory of Urban and Rural Ecological Environment, Beijing University of Agriculture, Beijing 102206, China),J.PlantResour. &Environ., 2016, 25(3): 80-89
To reveal variation rule of phenotypic traits of seed and seedling ofSyringareticulatasubsp.amurensis(Rupr.) P. S. Green et M. C. Chang from different locations and their relation with geographic-climatic factors, taking seed and seedling from ten provenances of Heilongjiang, Hebei, Beijing, Inner Mongolia, Qinghai and Shanxi (including six wild provenances and four cultivated provenances) as experimental materials, twelve phenotypic traits including length and width of seed, length and width of seed wing, 100-grain mass, germination rate, length and width of leaf, ratio of leaf length to leaf width, petiole length, height and ground diameter were measured. On this basis, variance analysis, coefficient of variation analysis, principal component analysis, cluster analysis and correlation analysis were carried out. The results show that there are obviously significant differences (P=0.01) in phenotypic traits of seed and seedling ofS.reticulatasubsp.amurensisamong different provenances, and phenotypic traits with great difference are germination rate, 100-grain mass, seed wing length and seed wing width. There are great differences in coefficient of variation (CV) of phenotypic traits within and among provenances,CVvalue within provenances is 1.39%-45.56%, and that among provenances is 6.45%-42.76%. In which, within provenances, averageCVvalues of seed wing length and seed wing width are larger, and those of germination rate and 100-grain mass are smaller, while among provenances, those of germination rate and 100-grain mass are larger. The result of principal component analysis shows that differences in phenotypic traits ofS.reticulatasubsp.amurensisare mainly concentrated in seed length, seed width and leaf length, and differences in height, germination rate, ground diameter, leaf width and seed wing width are the second. According to result of cluster analysis, ten provenances tested can be divided into four categories, in which, provenances from Beijing and Inner Mongolia and three provenances from Hebei are clustered to category Ⅰ, two provenances from Harbin of Heilongjiang are clustered to category Ⅱ, two provenances from Shanxi are clustered to category Ⅲ, one provenance from Qinghai is clustered to category Ⅳ alone, and the cluster result is related to geographical distribution of all provenances. The result of correlation analysis shows that there are different correlations among phenotypic traits and phenotypic traits with geographic-climatic factors, in which, there is no significant correlation of seed germination rate with other five phenotypic traits of seed, and correlation of longitude and latitude with all phenotypic traits is not significant, while altitude, annual mean temperature and frostless period are the main factors affecting variation of phenotypic traits. The comprehensive analysis result shows that there is obvious geographic genetic differentiation among different provenances ofS.reticulatasubsp.amurensis, and geographic isolation and great change in ecological factor cause rich variation of phenotypic traits of its seed and seedling. Based on research result, some suggestions about protection and utilization ofS.reticulatasubsp.amurensisare put forward.
Syringareticulatasubsp.amurensis(Rupr.) P. S. Green et M. C. Chang; phenotypic trait; geographic-climatic factor; cluster analysis; correlation analysis; geographic variation
2015-11-11
北京市科学技术委员会科技计划项目(Z121100007412003); 国家林木(含竹藤花卉)种质资源平台(2014年度); 北京市属高等学校创新团队建设与教师职业发展计划项目(IDHT20150503); 北京市教育委员会科技提升计划(PM2014014207)
杨晓霞(1989—),女,山东青岛人,硕士研究生,主要从事园林植物遗传资源与育种方面的研究。
E-mail: doudequan@bua.edu.cn
Q944; S685.26
A
1674-7895(2016)03-0080-10
10.3969/j.issn.1674-7895.2016.03.10
