5种甘草属植物花序和种子生产的位置效应及繁殖资源分配模式初步研究
2016-11-17陆嘉惠陈晓翠牛清东陈超南
韩 春, 陆嘉惠,b,c,①, 陈晓翠, 牛清东, 宋 凤, 陈超南
(石河子大学: a. 生命科学学院, b. 新疆生产建设兵团绿洲生态农业重点实验室, c. 甘草研究所, 新疆 石河子 832003)
5种甘草属植物花序和种子生产的位置效应及繁殖资源分配模式初步研究
韩春a, 陆嘉惠a,b,c,①, 陈晓翠a, 牛清东a, 宋凤a, 陈超南a
(石河子大学: a. 生命科学学院, b. 新疆生产建设兵团绿洲生态农业重点实验室, c. 甘草研究所, 新疆 石河子 832003)
以种植于新疆石河子的光果甘草(GlycyrrhizaglabraLinn.)、胀果甘草(G.inflataBatal.)、乌拉尔甘草(G.uralensisFisch.)、黄甘草(G.eurycarpaP. C. Li)和蜜腺甘草(G.glabravar.glandulosaX. Y. Li)为研究对象,对植株不同部位的花序数量、花序正常发育率、每花序单花数量和果穗干质量,以及植株不同部位和花序不同部位的生物量投入比、座果率、结籽率、种子投影面积和种子千粒质量进行测定;在此基础上,对供试5种甘草属(GlycyrrhizaLinn.)植物的繁殖资源分配模式和种子生产策略进行分析。结果表明:同一植株内,光果甘草、乌拉尔甘草、黄甘草和蜜腺甘草的花序数量、花序正常发育率、每花序单花数量和果穗干质量从植株下部到上部总体上依次递减,而胀果甘草植株不同部位间这4项指标总体上无显著差异。同一植株内,胀果甘草植株中部的生物量投入比和座果率均较高,但其生物量投入比、座果率和结籽率在植株不同部位间均无显著差异;而供试另4种植物的生物量投入比、座果率和结籽率从植株下部到上部总体上依次递减。同一花序内,胀果甘草花序中部的生物量投入比明显高于花序上部和下部,座果率从花序下部到上部依次递减,结籽率则在花序不同部位间无显著差异,而供试另4种植物的生物量投入比、座果率和结籽率从花序下部到上部总体上依次递减。供试5种植物的种子投影面积和种子千粒质量在植株不同部位间和花序不同部位间均无显著差异。综合研究结果显示:在资源竞争、结构效应和花粉限制的影响下,供试5种甘草属植物存在2种不同的资源分配模式和种子生产格局。其中,光果甘草、乌拉尔甘草、黄甘草和蜜腺甘草通过减少对晚发育的花或果实的资源投入来保障早发育的花或果实获得较多的资源,达到繁殖成功的目的;而胀果甘草则采取对花和果实随机败育的方式减小资源竞争的压力,这2种繁殖资源分配模式和种子生产策略对提高甘草属植物的繁殖成功率具有重要作用。
甘草属植物; 花序发育; 繁殖资源分配; 种子生产; 位置效应
花是植物的有性繁殖器官,其大小和结构对植物繁殖效率和交配系统进化有重要意义,因此,花的繁殖差异受到人们的广泛关注[1-3]。在植物多个花或由许多小花组成的花序中,不同花或花序之间存在不同程度的繁殖资源分配和竞争问题,即植株不同部位的花甚至同一个花序内不同部位的花的地位是不平等的,存在明显的位置效应[4],导致同株植物甚至同一花序不同位置的小花、果实和种子的数量、大小、形态及繁殖效率可能存在差异[5],其原因包括资源限制、传粉限制、同胞竞争和虫害作用等[6-7]。
豆科(Fabaceae)甘草属(GlycyrrhizaLinn.)植物全世界约20种[8],其中光果甘草(G.glabraLinn.)、乌拉尔甘草(G.uralensisFisch.)和胀果甘草(G.inflataBatal.)为《中华人民共和国药典》收载的药用甘草原植物[9],此外,黄甘草(G.eurycarpaP. C. Li)和蜜腺甘草(G.glabravar.glandulosaX. Y. Li)也可作为药用甘草的替代植物。由于野生甘草资源遭到过度采挖,造成甘草种群退化,野生资源匮乏[10],因此,人工种植甘草已成为甘草资源利用的主要途径。目前,对甘草植冠种子库[11]、种子结实特性[12]以及有性生殖过程和生殖败育[13]等方面的研究为甘草属植物的种子繁育特性和人工种植研究奠定了基础[14-15],但甘草属植物通常具有多个腋生总状花序,植株不同部位的花甚至同一花序内不同部位花的大小、结实率和结果率差异很大,其种子生产格局尚不清楚。揭示甘草属植物的花序和种子生产的位置效应,并比较不同种或者植株不同部位的繁殖资源分配的差异,对野生甘草属植物种群适应机制的研究和野生甘草资源的保护具有重要意义。
作者从种子生产格局和繁殖资源分配模式的角度,对光果甘草、乌拉尔甘草、胀果甘草、黄甘草和蜜腺甘草植株不同部位和花序不同部位的生物量投入情况、结实结籽特性及种子大小进行观察统计,分析甘草属植物结实和结籽格局的生态进化意义,以期为解决甘草属植物种子生产问题(获得质优种子)及野生甘草资源的可持续应用提供理论依据。
1 材料和方法
1.1材料
供试5种甘草属植物中,光果甘草、黄甘草、蜜腺甘草和乌拉尔甘草种植于石河子大学农学院试验站(东经123°45′、北纬44°45′),种植时间约30 a,均处于半野生状态;胀果甘草采集于石河子150团治沙站(东经123°45′、北纬45°06′),处于野生状态。种植区海拔350~450 m,年均气温6 ℃~10 ℃,平均无霜期155~186 d,≥10 ℃年有效积温3 000 ℃~3 500 ℃,年均降水量300~500 mm,降水主要集中在6月份至8月份,年均蒸发量1 500~2 000 mm。
各种类均随机标记15株样株(各样株间距5 m以上),于2014年9月份至10月份(果熟期)采集完整植株,带回实验室并自然风干。
1.2方法
根据甘草属植物的开花和结实特性,从第1个花序的生长部位开始,将植株按株高平均分为上部、中部和下部,分别统计植株各部位的总状花序数量、每花序单花数量(按花序轴上花柄留下的痕迹和果荚数量进行计算)和果荚数量,并称量果穗干质量。从每个花序第1朵花的着生部位开始,将花序按照长度平均分为上部、中部和下部,分别统计花序各部位的单花数量(按花序轴上花柄留下的痕迹和果荚数量进行计算)和果荚数量,称量果荚干质量,并计算座果率。分别统计各果荚内的种子数量并计算结籽率,称量种子千粒质量,测量种子投影面积。
用OHAUS千分之一电子天平(美国奥豪斯公司)称量果穗干质量、果荚干质量和种子千粒质量。采用数码图像处理方法测量种子投影面积:将种子随机铺在EPSON扫描仪玻璃板上,以1 mm2刻度纸为对照,调整像素为300 dpi,24位全彩对种子进行扫描,图片保存格式为“.tif”;采用Photoshop CS 5.0软件进行图片处理,调整图片为黑白格式,以减小色彩对数据的误差;依据1 mm2对应的像素点计算种子的投影面积[16-17]。
1.3数据计算和统计分析
分别按照下列公式计算各指标:花序正常发育率=(正常发育的花序数量/总花序数量)×100%; 座果率=(果荚数量/单花数量)×100%;结籽率=(种子数量/胚珠数量)×100%; 植株不同部位生物量投入比=(植株不同部位果穗干质量/整株果穗总干质量)×100%;花序不同部位生物量投入比=(同花序不同部位果荚干质量/同花序果穗总干质量)×100%。
采用SPSS 19.0软件对实验数据进行统计分析;采用one-way ANOVA法对相关数据进行方差分析,用Duncan’s新复极差法(SSR)进行多重比较。
2 结果和分析
2.1植株不同部位的繁殖资源分配和种子生产模式比较
2.1.1植株不同部位花序数量、花序正常发育率、每花序单花数量和果穗干质量的比较5种甘草属植物植株不同部位的花序数量、花序正常发育率、每花序单花数量和果穗干质量的比较结果见表1。
由表1可以看出:光果甘草、乌拉尔甘草、黄甘草和蜜腺甘草的植株中部和下部的花序数量、每花序单花数量和果穗干质量均较高,且总体上显著高于植株上部(P<0.05);其中,黄甘草的花序正常发育率从植株下部到上部依次递减,花序有明显的退化现象, 另外3种植物植株不同部位的花序正常发育率均高于90%,花序基本都能正常发育。胀果甘草植株不同部位间的花序数量、每花序单花数量和果穗干质量均无显著差异,但植株下部的花序正常发育率较高,且与植株上部和中部差异显著。研究结果表明:胀果甘草和黄甘草花序退化和每花序单花数量不足是导致其繁殖力低的主要原因。
2.1.2植株不同部位生物量投入比、座果率、结籽率、种子投影面积和千粒质量的比较5种甘草属植物植株不同部位的生物量投入比、座果率、结籽率、种子投影面积和千粒质量的比较结果见表2。
由表2可以看出:光果甘草、乌拉尔甘草、黄甘草和蜜腺甘草的生物量投入比均从植株下部到上部依次递减,繁殖资源投入的递减趋势明显;而胀果甘草植株不同部位间的生物量投入比无显著差异。5种甘草属植物中,乌拉尔甘草和黄甘草植株中部和下部的座果率和结籽率高于植株上部,且总体上差异显著;其余3种甘草属植物植株不同部位间的座果率和结籽率均无显著差异。供试5种甘草属植物植株不同部位间的种子投影面积和千粒质量均无显著差异,说明这5种甘草属植物植株不同部位的种子大小受繁殖资源竞争的影响较小。
2.2植株上部、中部和下部同一花序的不同部位种子生产模式比较
在5种甘草属植物植株上部、中部和下部,同花序内不同部位的生物量投入比、座果率、结籽率、种子投影面积和种子千粒质量的比较结果分别见表3、表4和表5。


种类Species部位Position花序数量Inflorescencenumber花序正常发育率/%Inflorescencenormaldevelopmentrate每花序单花数量Single-flowernumberperinflorescence果穗干质量/gEardryweight光果甘草G.glabra上部Upper3.4±0.4b96.66±2.24a25.5±1.9b1.85±0.30c中部Middle3.8±0.4b95.83±4.16a32.8±2.0a3.20±0.43b下部Lower6.8±1.2a95.27±4.15a33.5±2.7a4.69±0.62a胀果甘草G.inflata上部Upper3.3±0.5a79.50±7.67b15.5±1.0a0.49±0.11a中部Middle4.5±0.8a80.78±4.47b16.6±1.1a0.76±0.14a下部Lower3.5±1.1a91.67±4.81a14.7±1.2a0.59±0.20a乌拉尔甘草G.uralensis上部Upper3.2±0.5b93.59±4.44a25.3±2.4a1.59±0.43b中部Middle6.3±1.1a96.38±2.02a26.1±1.9a4.70±0.56a下部Lower6.0±0.9a98.86±0.94a28.5±1.6a5.54±0.78a黄甘草G.eurycarpa上部Upper2.3±0.6c24.00±11.22c19.1±1.3b0.25±0.16b中部Middle6.1±1.1b75.45±7.56b19.3±1.2b0.91±0.23a下部Lower9.3±1.2a91.21±5.04a22.3±1.1a2.40±0.30a蜜腺甘草G.glabravar.glandulosa上部Upper3.0±0.2b93.33±4.49a23.2±1.6b1.26±0.21c中部Middle3.3±0.2b100.00±0.00a31.1±1.7a3.29±0.41b下部Lower4.8±0.5a98.99±1.01a34.9±2.4a7.47±0.76a
1)同列中不同的小写字母表示同一指标在同种类的不同部位间差异显著(P<0.05) Different small letters in the same column indicate the significant difference of the same index among different positions of the same species (P<0.05).
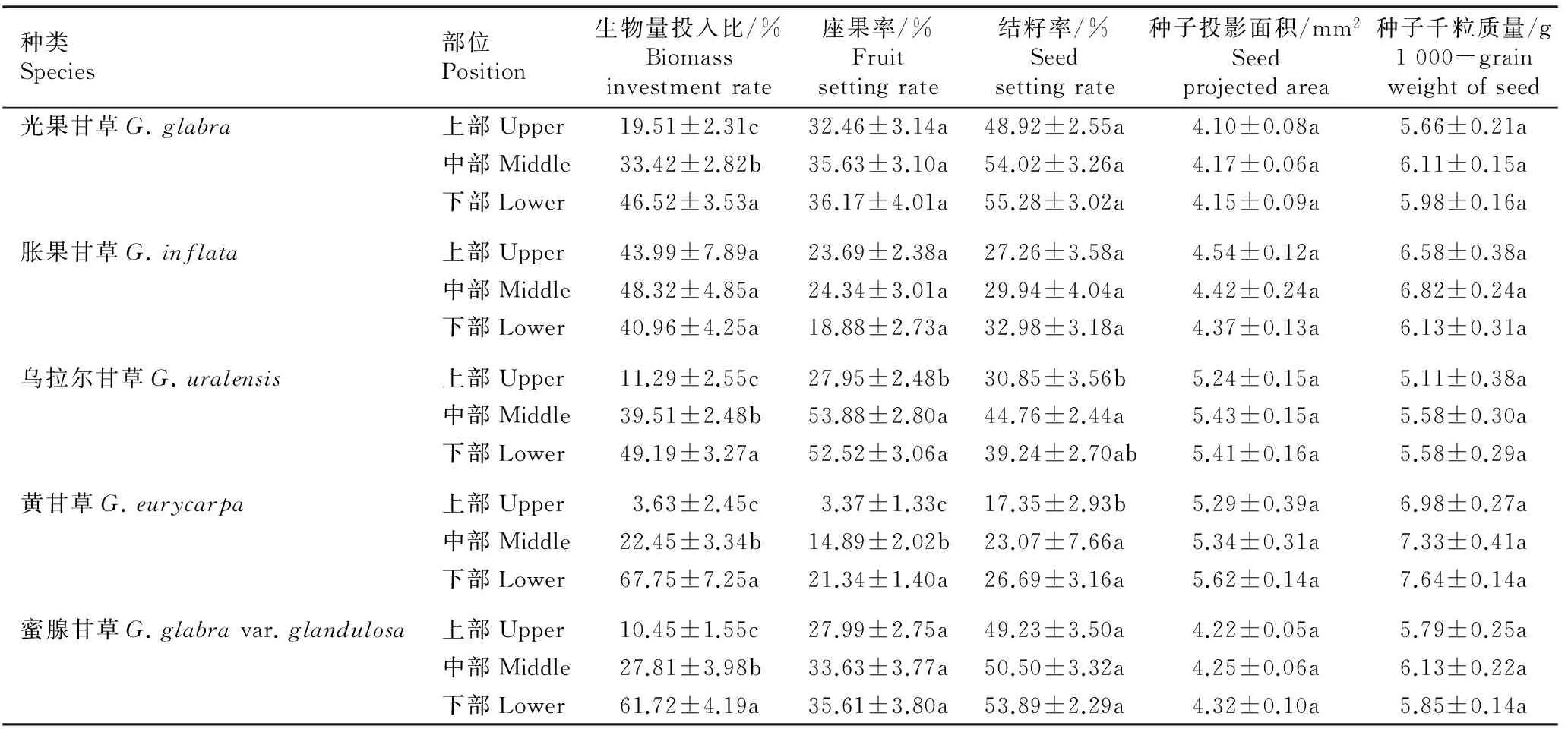
种类Species部位Position生物量投入比/%Biomassinvestmentrate座果率/%Fruitsettingrate结籽率/%Seedsettingrate种子投影面积/mm2Seedprojectedarea种子千粒质量/g1000-grainweightofseed光果甘草G.glabra上部Upper19.51±2.31c32.46±3.14a48.92±2.55a4.10±0.08a5.66±0.21a中部Middle33.42±2.82b35.63±3.10a54.02±3.26a4.17±0.06a6.11±0.15a下部Lower46.52±3.53a36.17±4.01a55.28±3.02a4.15±0.09a5.98±0.16a胀果甘草G.inflata上部Upper43.99±7.89a23.69±2.38a27.26±3.58a4.54±0.12a6.58±0.38a中部Middle48.32±4.85a24.34±3.01a29.94±4.04a4.42±0.24a6.82±0.24a下部Lower40.96±4.25a18.88±2.73a32.98±3.18a4.37±0.13a6.13±0.31a乌拉尔甘草G.uralensis上部Upper11.29±2.55c27.95±2.48b30.85±3.56b5.24±0.15a5.11±0.38a中部Middle39.51±2.48b53.88±2.80a44.76±2.44a5.43±0.15a5.58±0.30a下部Lower49.19±3.27a52.52±3.06a39.24±2.70ab5.41±0.16a5.58±0.29a黄甘草G.eurycarpa上部Upper3.63±2.45c3.37±1.33c17.35±2.93b5.29±0.39a6.98±0.27a中部Middle22.45±3.34b14.89±2.02b23.07±7.66a5.34±0.31a7.33±0.41a下部Lower67.75±7.25a21.34±1.40a26.69±3.16a5.62±0.14a7.64±0.14a蜜腺甘草G.glabravar.glandulosa上部Upper10.45±1.55c27.99±2.75a49.23±3.50a4.22±0.05a5.79±0.25a中部Middle27.81±3.98b33.63±3.77a50.50±3.32a4.25±0.06a6.13±0.22a下部Lower61.72±4.19a35.61±3.80a53.89±2.29a4.32±0.10a5.85±0.14a
1)同列中不同的小写字母表示同一指标在同种类的不同部位间差异显著(P<0.05) Different small letters in the same column indicate the significant difference of the same index among different positions of the same species (P<0.05).
由表3至表5的数据可以看出:在光果甘草、乌拉尔甘草、黄甘草和蜜腺甘草植株的上部、中部和下部,同一花序内的生物量投入比、座果率和结籽率总体上从花序下部到上部依次递减;而在胀果甘草植株的上部、中部和下部,同花序内中部的生物量投入比明显高于花序上部和下部,其座果率则从花序下部到上部依次递减,其结籽率则在花序不同部位间无显著差异。在供试5种甘草属植物植株的不同部位,同花序不同部位间的种子投影面积和千粒质量均无显著差异,说明这5种甘草属植物的花序繁殖资源分配对其种子大小的影响较小,一旦繁殖成功便能形成正常大小的种子。

种类Species花序部位Positionofinflores-cence生物量投入比/%Biomassinvestmentrate座果率/%Fruitsettingrate结籽率/%Seedsettingrate种子投影面积/mm2Seedprojectedarea种子千粒质量/g1000-grainweightofseed光果甘草G.glabra上部Upper21.84±5.83c20.60±4.93c45.80±4.90b3.97±0.14a5.31±0.34a中部Middle33.64±3.65b39.17±6.00b49.14±3.67a4.11±0.16a5.53±0.39a下部Lower44.52±6.21a48.73±7.02a52.78±4.82a4.22±0.13a6.11±0.34a胀果甘草G.inflata上部Upper24.46±8.27b12.90±2.91b23.92±5.20a4.19±0.30a5.61±0.48a中部Middle40.61±6.26a15.27±3.90b26.69±4.81a4.30±0.25a6.02±0.58a下部Lower34.93±4.29a28.47±5.72a32.54±3.43a4.61±0.08a6.35±0.35a乌拉尔甘草G.uralensis上部Upper22.21±4.68c22.60±2.06b25.20±3.69b5.38±0.23a4.89±0.24a中部Middle30.86±4.60b29.86±2.70a27.50±3.70b5.39±0.25a5.09±0.67a下部Lower46.93±2.93a33.39±3.87a38.04±5.18a5.45±0.32a5.22±0.47a黄甘草G.eurycarpa上部Upper5.75±4.33c1.49±1.32a8.60±1.60b4.61±0.00a—中部Middle31.01±15.51b5.02±2.99a26.51±3.11a5.18±0.58a7.43±0.10a下部Lower63.24±18.62a3.61±2.41a32.94±3.73a5.59±0.32a7.86±0.15a蜜腺甘草G.glabravar.glandulosa上部Upper16.23±14.86c18.83±4.20b36.85±3.92b4.16±0.20a5.64±0.53a中部Middle36.86±12.45b30.16±4.17ab54.44±4.58a4.29±0.13a5.89±0.35a下部Lower46.92±16.15a34.99±4.78a48.74±5.06a4.44±0.18a6.08±0.39a
1)同列中不同的小写字母表示同一指标在同种类的花序不同部位间差异显著(P<0.05) Different small letters in the same column indicate the significant difference of the same index among different positions of inflorescence of the same species (P<0.05). —: 表示数据缺失 Showing data missing.

种类Species花序部位Positionofinflores-cence生物量投入比/%Biomassinvestmentrate座果率/%Fruitsettingrate结籽率/%Seedsettingrate种子投影面积/mm2Seedprojectedarea种子千粒质量/g1000-grainweightofseed光果甘草G.glabra上部Upper19.10±4.66b27.75±6.31b48.50±3.90b4.11±0.12a5.70±0.30a中部Middle37.28±2.75a36.03±4.29ab52.79±2.76a4.18±0.10a6.21±0.20a下部Lower43.62±4.57a43.10±4.65a56.51±3.71a4.21±0.10a6.42±0.24a胀果甘草G.inflata上部Upper30.39±5.38b19.76±3.23b26.08±3.16a4.18±0.62a6.69±0.26a中部Middle44.95±5.23a24.81±3.79b27.79±2.78a4.54±0.17a6.76±0.44a下部Lower22.61±4.66c26.48±3.22a29.70±3.70a4.56±0.33a6.99±0.52a乌拉尔甘草G.uralensis上部Upper25.43±1.71b47.75±3.99b41.73±4.09b5.39±0.27a5.34±0.57a中部Middle37.03±1.70a56.69±5.13ab44.66±4.32a5.39±0.27a5.69±0.55a下部Lower37.54±2.34a57.19±5.26a47.89±4.39a5.52±0.28a5.71±0.47a黄甘草G.eurycarpa上部Upper18.78±3.93c10.19±2.20b19.37±2.17a5.42±0.29a6.59±0.53a中部Middle33.18±4.47b14.70±2.53ab18.88±2.07a5.08±0.39a7.42±0.63a下部Lower44.04±6.72a18.95±2.34a21.20±2.60a5.63±0.29a7.88±0.46a蜜腺甘草G.glabravar.glandulosa上部Upper16.24±3.79b24.79±4.78b47.12±5.63a4.03±0.11a5.75±0.39a中部Middle40.26±3.54a48.29±4.72a53.06±3.65a4.30±0.09a6.17±0.30a下部Lower43.50±3.83a55.45±6.18a50.71±4.20a4.37±0.09a6.34±0.46a
1)同列中不同的小写字母表示同一指标在同种类的花序不同部位间差异显著(P<0.05) Different small letters in the same column indicate the significant difference of the same index among different positions of inflorescence of the same species (P<0.05).

种类Species花序部位Positionofinflores-cence生物量投入比/%Biomassinvestmentrate座果率/%Fruitsettingrate结籽率/%Seedsettingrate种子投影面积/mm2Seedprojectedarea种子千粒质量/g1000-grainweightofseed光果甘草G.glabra上部Upper19.38±4.99c19.19±4.65c49.34±5.53b4.06±0.21a5.78±0.26a中部Middle36.38±2.39b32.80±3.81b54.99±5.37ab4.10±0.15a5.91±0.27a下部Lower44.25±3.78a45.39±4.61a62.75±3.96a4.28±0.11a6.21±0.28a胀果甘草G.inflata上部Upper25.27±5.94b17.46±3.44b31.19±4.67a4.86±0.16a6.28±0.73a中部Middle46.33±8.51a21.95±5.94ab32.02±5.89a4.47±0.11a6.51±0.66a下部Lower28.40±6.78b33.62±5.00a37.19±4.26a4.29±0.19a7.10±0.54a乌拉尔甘草G.uralensis上部Upper27.11±1.60c43.55±4.77b35.48±2.97b5.23±0.29a5.17±0.52a中部Middle40.08±2.05a53.00±5.23ab38.91±4.19a5.36±0.26a5.63±0.46a下部Lower32.81±1.43b60.99±5.13a43.32±4.92a5.13±0.26a5.95±0.54a黄甘草G.eurycarpa上部Upper20.52±3.38c15.20±2.81b23.02±2.51b5.38±0.29a6.29±0.46a中部Middle43.34±2.30a23.23±1.42a28.00±2.77ab5.46±0.18a7.15±0.44a下部Lower36.14±3.35b25.39±1.83a32.45±2.20a5.96±0.27a7.39±0.49a蜜腺甘草G.glabravar.glandulosa上部Upper23.14±3.70b41.25±6.81b48.66±4.90b4.16±0.08a5.65±0.26a中部Middle38.04±1.69a58.24±4.92ab57.04±6.48a4.20±0.08a5.93±0.23a下部Lower38.82±3.23a67.09±5.88a59.96±5.32a4.30±0.09a5.98±0.26a
1)同列中不同的小写字母表示同一指标在同种类的花序不同部位间差异显著(P<0.05) Different small letters in the same column indicate the significant difference of the same index among different positions of inflorescence of the same species (P<0.05).
3 讨论和结论
3.1位置效应对5种甘草属植物植株不同部位繁殖资源投入和繁殖成功的影响
通常情况下,植物在整个生长季节内获得的资源量是有限的,其营养生长与生殖生长之间的资源分配权衡问题一直是植物生活史研究的热点[18],资源分配模式在很大程度上反映了植物的生活史特征[19]。本研究中,光果甘草、乌拉尔甘草、黄甘草和蜜腺甘草的繁殖资源投入从植株下部到上部依次递减,表明这4种甘草属植物的繁殖资源分配明显受到资源竞争的影响,植株下部花序开花早且靠近母体资源库,在资源竞争上具有明显优势,使植株下部的花序数量和正常发育的花序数量增加,果穗干质量增大,座果率和结籽率提高;越到植株上部供给繁殖的资源越少,花序数量也趋少,植物有选择性地使植株上部或中部的花序和花败育,以减小资源竞争的压力,使部分花序和花能获取更多的母本资源并能够发育形成正常的果实和种子,因而,植株上部繁殖资源投入较低,座果率和结籽率也相对较低,这也是在繁殖资源受限时植物通过权衡繁殖资源分配比形成的差异化的繁殖资源分配格局,苜蓿(MedicagosativaLinn.)[20]、淫羊藿(EpimediumbrevicornuMaxim.)[21]和大花百子莲(Agapanthuspraecoxsubsp.orientalis‘Big Blue’)[22]等种类也具有相似结实格局。胀果甘草植株不同部位的繁殖资源投入、座果率和结籽率均无显著差异,单纯的资源竞争似乎无法解释这种资源分配格局和结实现象。作者在野外调查中观察到胀果甘草有严重的落花落果现象,可能与其生长所需光热要求较高[14]有关,在繁殖资源有限的情况下,胀果甘草选择随机败育的方式淘汰了大量“多余”的花,它们有可能被作为雄性角色来利用,也可能作为雌性角色的“备选”以增加繁殖成功率,从而形成了胀果甘草植株不同部位间无差异的繁殖资源分配格局。供试5种植物具有的这2种不同的繁殖资源分配模式对生长在荒漠干旱区的甘草属植物提高生存适合度和繁殖成功率具有重要生态学意义[23]。
3.2位置效应对5种甘草属植物花序不同部位繁殖资源投入和繁殖成功的影响
Diggle[24]认为,花序内不同部位间的资源分配存在显著的变化趋势。一般顶向发育的花序,其基部较上部具有较大的花[25],较大或较多的胚珠、种子和果实以及较低的P/O比(花粉胚珠比)等[1],即使在同一花序内,不同部位花的地位也不平等,存在显著的位置效应,导致资源分配模式也存在差异。目前解释花序内资源投入差异主要有3种假说:1)资源竞争假说(resource competition hypothesis),即花序内的子房竞争有限的资源,早发育和(或)空间位置位于资源近端的果实比迟发育和位于资源远端的果实能获得更多的资源[6,26];2)结构效应假说(architectural effect hypothesis),即花序内不同位置的花获得的资源与花序轴维管组织的发育状况及大小有关[6,27];3)花粉限制假说(pollen limitation hypothesis),即在同一花序内,由于不同部位的花所接受的花粉来源和数量不同,使不同部位的花由于自交和异交的差异而导致座果率和结籽率的变化[5,28-29]。
本研究中,光果甘草、乌拉尔甘草、黄甘草和蜜腺甘草的结实格局是从花序下部到上部果实和种子产量逐渐减小,生物量投入比也逐渐减小,其繁殖资源分配格局与结果结实特性可用上述花序内资源投入的3种假说进行解释。一方面,根据资源竞争假说,这4种甘草属植物的单花开放顺序从花序下部开始逐渐向上,最早开放的花在资源吸收和利用上占有主动权,能够获得较多的母体资源,因此花序下部果实成熟的可能性最高,使花序下部座果率和结籽率高;而花序上部的单花开花较晚,花和果实发育受到资源竞争的制约,花序上部的部分单花甚至出现萎蔫或无法正常开放的现象,导致花序上部座果率和结籽率低。另一方面,根据结构效应假说,总状花序的花序轴直径、维管数量和运输营养物质效率等均沿着花序轴逐渐伸长而明显降低[30],导致母体资源运输受到限制,花序上部的花发育受到影响,使座果率和结籽率较低。此外,甘草属植物为顶向开花的总状花序,传粉昆虫在花序上的运动方式也是自下向上[31],这样容易使花序下部的花更趋向得到异源花粉,增加杂交成功概率,提高结实率和结籽率;而花序下部的花粉往往被带到花序上部,造成花序上部的花自交而增加败育概率[32-33],因此,这也是导致这4种甘草属植物花序上部座果率和结籽率较低的原因之一。胀果甘草的繁殖资源分配格局、结果结实特性与供试另外4种甘草属植物明显不同,同一花序内,胀果甘草的座果率从花序下部到上部依次递减,而结籽率则无显著差异,说明胀果甘草的繁殖成功同样受资源竞争、结构效应和花粉限制的影响;但生物量投入比在花序中部较高,说明胀果甘草花序不同部位的繁殖资源分配模式不能仅仅用花序内资源分配的3种假说来解释,还与胀果甘草的资源分配效应和花序发育特性有关,推测在花序发育过程中可能存在某种生理调控机制。刘左军等[4]认为,植物体中生长发育越旺盛的部位其资源需求信号越强,母体可利用资源的投入也越多。这可能是胀果甘草因资源限制造成花序内位置资源分配模式和结实结籽格局与供试另外4种甘草属植物不同的直接原因,但这种生理学机制以及这种机制与环境变化的关系还有待进一步实验验证。
3.35种甘草属植物繁殖资源分配对种子大小和繁殖成功的影响
供试5种甘草属植物植株不同部位和同一花序内种子投影面积(种子大小)和种子千粒质量均无显著差异,这可能是甘草属植物在繁殖资源有限的情况下对种子数量与质量之间作出的权衡。在资源竞争、结构效应和花粉限制等综合因素的影响下,植物对花序内部分花采取落花(如胀果甘草)或对胚珠选择性败育,保证了部分果荚和种子能够正常发育,使繁殖成效达到最大化,对提高繁殖成功率具有重要作用。
综上所述,5种甘草属植物具有2种不同的繁殖资源分配模式和种子生产策略。光果甘草、乌拉尔甘草、黄甘草和蜜腺甘草在植株不同部位的繁殖资源分配模式具有明显的位置效应,受繁殖资源竞争的影响较大,繁殖资源投入和繁殖成功率均依次递减;而胀果甘草植株不同部位则采用繁殖资源平均分配的模式,且繁殖成功率无差异。同一花序内,光果甘草、乌拉尔甘草、黄甘草和蜜腺甘草的繁殖资源分配模式主要受资源竞争、结构效应和花粉限制的影响;而胀果甘草的繁殖资源分配模式不仅受资源竞争、结构效应和花粉限制的影响,还可能与花序发育过程中存在的某种生理调控机制有关。不同的繁殖资源分配模式是甘草属各种类长期适应各自生长环境的结果,代表了不同的繁殖策略,但由于本研究结果仅包含1年的观测数据,因而,这种繁殖资源分配模式是否随环境气候因子(如土壤、温度、气候等)的变化而发生改变,还有待进一步深入研究。此外,种子生产的位置效应对种子萌发率[34]及幼苗存活和建植[35]均有一定影响,甘草属植物种子生产的位置效应是否对其种子萌发、幼苗生长有影响,也有待后续研究。
[1]MAZER S J, DAWSON K A. Size-dependent sex allocation within flowers of the annual herbClarkiaunguiculata(Onagraceae): ontogenetic and among-plant variation[J]. American Journal of Botany, 2001, 88: 819-831.
[3]王怡, 黎云祥, 高辉, 等. 位置效应对巫山淫羊藿花蜜分泌和结实的影响[J]. 广西植物, 2013, 33(6): 769-773.
[4]刘左军, 杜国祯, 陈家宽. 黄帚橐吾花序结构的资源配置与环境的关系[J]. 植物生态学报, 2003, 27(3): 344-351.
[5]樊宝丽, 赵志刚, 孟金柳, 等. 露蕊乌头(AconitumgymnandrumMaxim)花序内位置依赖的性分配[J]. 生态学报, 2008, 28(6): 2909-2915.
[6]MEDRANOM, GUITINP, GUITINJ. Patternsoffruitand seed set within inflorescences ofPancratiummaritimum(Amaryllidaceae): nonuniform pollination, resource limitation, or architectural effects [J]. American Journal of Botany, 2000, 87: 493-501.
[7]HOLLAND J N, BRONSTEIN J L, DEANGELIS D L.Testing hypo-theses for excess flower production and low fruit-to-flower ratios in a pollinating seed-consuming mutualism[J]. Oikos, 2004, 105: 633-640.
[8]李学禹. 甘草属分类系统与新分类群的研究[J]. 植物研究, 1993, 13(1): 14-43.
[9]国家药典委员会.中华人民共和国药典: 2005年版(一部)[M]. 北京: 化学工业出版社, 2005: 59-60.
[10]李学斌, 陈林, 李国旗, 等. 中国甘草资源的生态分布及其繁殖技术研究[J]. 生态环境学报, 2013, 22(4): 718-722.
[11]刘芳, 肖彩虹. 野生甘草资源植冠种子库特性的初步研究[J]. 干旱区资源与环境, 2008, 22(3): 175-177.
[12]王丹, 王俊杰. 野生乌拉尔甘草结实特性及种子产量影响因子分析[J]. 内蒙古农业大学学报(自然科学版), 2015, 36(1): 59-64.
[13]方玉强. 甘草有性生殖过程及生殖败育的初步研究[D]. 北京: 北京中医药大学中药学院, 2013: 41-65.
[14]李学禹, 陆嘉惠. 甘草属(GlycyrrhizaL.)分类系统与实验生物学研究[M]. 上海: 复旦大学出版社, 2015: 15-28.
[15]马春英, 王文全, 张学静, 等. 乌拉尔甘草花部特征和开花结荚特性的研究[J]. 植物遗传资源学报, 2009, 10(2): 295-299.
[16]骆郴, 贾亚敏, 刘彤, 等. 利用普通扫描仪精确测量叶面积的技术及方法[J]. 北方园艺, 2007(5): 46-48.
[17]王琳, 刘彤, 韩志全, 等. 古尔班通古特沙漠植物种子大小变异的空间格局[J]. 生态学报, 2014, 34(23): 6797-6806.
[18]SCHMID B, WEINER J. Plastic relationships between reproductive and vegetative mass inSolidagoaltissima[J]. Evolution, 1993, 47: 61-74.
[19]任明迅, 姜新华, 张大勇. 植物繁殖生态学的若干重要问题[J]. 生物多样性, 2012, 20(3): 241-249.
[20]张爱勤, 朱进忠. 苜蓿结实格局及其影响因素的研究[J]. 新疆农业大学学报, 2005, 28(4): 15-19.
[21]杨子松, 权秋梅, 黎云祥, 等. 淫羊藿开花的时空效应对其结实的影响[J]. 西北植物学报, 2009, 29(8): 1599-1606.
[22]孙颖, 马翠青, 陈士惠, 等. 大花百子莲的结实和结籽格局及种子产量影响因素分析[J]. 植物资源与环境学报, 2014, 23(2): 48-53.
[23]王喜勇, 高蕊, 周晓青, 等. 角果藜的结实格局及地下散布单位的入土机制[J]. 草业学报, 2014, 23(2): 352-357.
[24]DIGGLE P K. Architectural effects and the interpretation of patterns of fruit and seed development[J]. Annual Review of Ecology and Systematics, 1995, 26: 531-552.
[25]ASHMAN T L, HITCHENS M S. Dissecting the causes of variation in intra-inflorescence allocation in a sexually polymorphic species,Fragariavirginiana(Rosaceae)[J]. American Journal of Botany, 2000, 87: 197-204.
[26]STEPHENSON A G. Flower and fruit abortion: proximate causes and ultimate functions[J]. Annual Review of Ecology and Systematics, 1981, 12: 253-279.
[27]BUIDE M L. Disentangling the causes of intrainflorescence variation in floral traits and fecundity in the hermaphroditeSileneacutifolia[J]. American Journal of Botany, 2008, 95: 490-497.
[28]BRUNET J. Male reproductive success and variation in fruit and seed set inAquilegiacaerulea(Ranunculaceae)[J]. Ecology, 1996, 77: 2458-2471.
[29]BROOKES R H, JESSON L K, BUR M. Reproductive investment within inflorescences ofStylidiumarmeriavaries with the strength of earlyresourcecommitment[J]. Annalsof Botany, 2010, 105: 697-705.
[30]邹四轩. 紫萼花序内花粉和胚珠数目变化的机制: 施肥系列的验证[D]. 成都: 四川农业大学林学院, 2013: 4-6.
[31]廖云海, 陆嘉惠, 张际昭, 等. 光果甘草生殖生物学特性的初步研究[J]. 西北植物学报, 2010, 30(5): 939-943.
[32]张大勇. 植物生活史进化与繁殖生态学[M]. 北京: 科学出版社, 2004: 100-113.
[33]张大勇, 姜新华. 植物交配系统的进化、资源分配对策与遗传多样性[J]. 植物生态学报, 2001, 25(2): 130-143.
[34]BUIDEML.PollinationecologyofSileneacutifolia(Caryophyllaceae): floral traits variation and pollinator attraction[J]. Annals of Botany, 2006, 97: 289-297.
[35]谢田朋, 杜国祯, 张格非, 等. 黄帚橐吾种子生产的花序位置效应及其对幼苗建植的影响[J]. 植物生态学报, 2010, 34(4): 418-426.
(责任编辑: 张明霞)
Preliminary study on position effect of inflorescence and seed production, and reproductive resource allocation model of five species in
GlycyrrhizaLinn.HAN Chuna, LU Jiahuia,b,c,①, CHEN Xiaocuia, NIU Qingdonga, SONG Fenga, CHEN Chaonana
(Shihezi University: a. College of Life Sciences, b. The Key Laboratory of Oasis Eco-Agriculture, Xinjiang Production and Construction Group, c. Institute of Licorice, Shihezi 832003, China),J.PlantResour. &Environ., 2016, 25(3): 72-79
TakingGlycyrrhizaglabraLinn.,G.inflataBatal.,G.uralensisFisch.,G.eurycarpaP. C. Li andG.glabravar.glandulosaX. Y. Li planted in Shihezi of Xinjiang as researched objects, inflorescence number, inflorescence normal development rate, single-flower number per inflorescence and ear dry weight at different positions of plant body, and biomass investment rate, fruit setting rate, seed setting rate, seed projected area and 1 000-grain weight of seed at different positions of plant body and inflorescence were determined. On this basis, reproductive resource allocation model and seed production strategy of these five species inGlycyrrhizaLinn. tested were analyzed. The results show that within a plant body, inflorescence number, inflorescence normal development rate, single-flower number per inflorescence and ear dry weight ofG.glabra,G.uralensis,G.eurycarpaandG.glabravar.glandulosagenerally decrease successively from lower part to upper part of plant body, while there is no significant difference in these four indexes among different positions of plant body ofG.inflata. Within a plant body, biomass investment rate and fruit setting rate at middle part of plant body ofG.inflataare higher, but there is no significant difference in its biomass investment rate, fruit setting rate and seed setting rate among different positions of plant body, while, biomass investment rate, fruit setting rate and seed setting rate of other four species tested generally decrease successively from lower part to upper part of plant body. Within a inflorescence, biomass investment rate at middle part of inflorescence ofG.inflatais higher than that at upper part and lower part of inflorescence, its fruit setting rate decreases successively from lower part to upper part of inflorescence, and there is no significant difference in its seed setting rate among different positions of inflorescence, while, biomass investment rate, fruit setting rate and seed setting rate of other four species tested generally decrease successively from lower part to upper part of inflorescence. There are no significant difference in seed projected area and 1 000-grain weight of seed among different positions of plant body and inflorescence of five species tested. The comprehensive research result shows that under the influences of resource competition, structure effect and pollen limitation, there are two different resource allocation models and seed production patterns of five species inGlycyrrhizatested. In which,G.glabra,G.uralensis,G.eurycarpaandG.glabravar.glandulosaensure early-development flower or fruit to earn more resources by reducing resources input in late-development flower or fruit and achieve the aim of successful reproduction, whileG.inflatareduces resource competition pressure by measure of random abortion in flower and fruit. These two reproductive resource allocation models and seed production strategies have an important effect on enhancing success rate of reproduction ofGlycyrrhizaspecies.
GlycyrrhizaLinn.; inflorescence development; reproductive resource allocation; seed production; position effect
2016-01-05
国家自然科学基金资助项目(31260042)
韩春(1991—),男,甘肃武威人,硕士研究生,主要从事药用植物资源方面的研究。
E-mail: jiahuil@shzu.edu.cn
Q944.59; S567.7+1
A
1674-7895(2016)03-0072-08
10.3969/j.issn.1674-7895.2016.03.09
