夏季南极长城湾海流变化特征及其对浮游植物生物量的影响
2016-11-11罗光富何剑锋张芳蔡明红林凌蓝木盛
罗光富,何剑锋,2*,张芳,蔡明红,林凌,蓝木盛
(1. 中国极地研究中心 国家海洋局极地科学重点实验室,上海 200136;2. 同济大学 环境科学与工程学院,上海 200092)
夏季南极长城湾海流变化特征及其对浮游植物生物量的影响
罗光富1,何剑锋1,2*,张芳1,蔡明红1,林凌1,蓝木盛1
(1. 中国极地研究中心 国家海洋局极地科学重点实验室,上海 200136;2. 同济大学 环境科学与工程学院,上海 200092)
通过对2010/2011年夏季南极长城湾叶绿素a浓度和海流等环境参数的传感器连续观测,研究其时空变化特征及海流对浮游植物生物量的影响。结果表明,12.9 m水层叶绿素a浓度从12月中旬开始增长,直至2月上旬开始维持相对稳定,期间出现两个高值点(2.74 μg/L 和3.76 μg/L)。叶绿素a浓度表现出了24 h周期变化特征,每日叶绿素a浓度最高值出现在正午前后时段的概率要高于其他时段。表层海水受西北风影响,流速较大,约为60~100 cm/s;中层和底层水体流速较小,约为10 cm/s。表层的风海流使海水在长城湾发生辐散,下层海水向上涌升,驱动了水体的垂直交换。长城湾与麦克斯韦尔湾存在水体交换,水体主要从中层进入湾内,从表层输出。长城湾的叶绿素a浓度与流速呈显著负相关,表明低流速环境有利于长城湾浮游植物的生长。
叶绿素a浓度;海流;浮游植物;长城湾;南极
1 引言
全球变暖引发了南大洋与南极大陆一系列的生态环境问题,如中低纬度浮游植物种类向极地海域入侵[1—2]、部分大型海洋生物数量下降[3]、冰盖融化引起全球海平面上升[4—5]等。南极长城站所在的南极半岛地区是全球升温最快的区域之一[6—7],长城湾是我国依托考察站的南极重点研究海域。浮游植物是海洋食物链的基础,在海洋生态系统能量流动和全球碳循环中扮演着重要角色,其初级生产量约占整个生物圈净初级生产量的一半[1,8—11]。1984年我国开始南极科学考察以来,关于长城湾浮游植物种类组成[12]、叶绿素a浓度变化[13—15]等的研究已有众多报道。相关研究结果表明长城湾浮游植物主要以冷水性和近岸广温性种类为主,优势种随季节变化而出现演替[12];叶绿素a浓度年际变化和次表层极大值现象显著[13—15]。
对于相关环境因子的观测则主要集中在温度、盐度[16]、营养盐[17]、潮汐[18]、光合有效辐射[19]等,然而有关重要环境因子之一海流的时空变化则一直缺少相对系统的研究。有研究表明,海流对水体中浮游生物幼体的播散和种群的维持起着重要作用[20]。浮游植物生长所需的营养盐可以通过海流输运到特定区域或水层,如上升流可以将底层营养盐输送到表层[21]。流速对浮游植物群落结构[22—23]、生长繁殖[24—26]及生物量[27]等均有影响。本文通过对长城湾连续100 d(27次南极科学考察期间,2010年12月13日—2011年3月22日)的叶绿素a浓度和海流观测数据进行分析,研究它们的时空变化特征,探究长城湾海流变化对浮游植物生物量的潜在影响。
2 研究区域与方法
2.1研究区域概况
长城湾位于菲尔德斯半岛中国南极长城站东南侧,为一个半封闭型内湾,面积约3 km2,距湾顶约200 m处有一深水区,最大水深约35 m[16]。湾内存在许多明、暗礁石,在湾口的中部有一小岛,通过麦克斯韦尔湾与布兰斯菲尔德海峡相通。湾顶的东北部,在涨潮时与阿德雷湾相连,落潮时形成一条砾石小堤,将两湾隔断(图1)。长城湾的潮汐属于不规则半日潮,年平均潮差为1.243 m[18]。长城站常年受到亚南极海洋性气团和极地气旋的影响,具有阴天多、湿度大、大风和暴风雪多的气候特点,夏季(12—2月)月平均温度集中在-1.2~2.9℃之间,冬季(6—8月)在-13.4~-1.0℃之间变化。1985年至2008年长城站站区的气温平均每10年升高0.27℃,呈明显的升温趋势[28]。站区冬半年平均风速9 m/s,夏半年7.7 m/s[18]。风力大小对长城湾平均水位有一定的影响,夏季长城湾风力相对较弱,只能引起短暂的增水或减水现象,平均水位的变幅较小[18]。长城湾位于南极半岛以北,与南极其他海域相比,由于纬度低,海水较浅,海冰具有独特的特征。海冰覆盖时间短,稳定性差,湾内海冰并非年年形成。文献资料[29—30]表明长城湾海冰一般形成于4—5月,7—10月冰情相对稳定,11月海冰开始慢慢融化,至12月海冰已经基本消失殆尽。
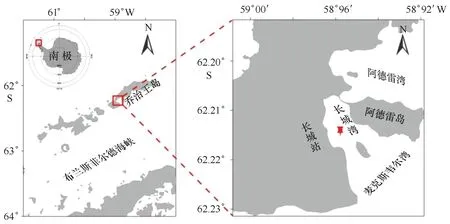
图1 长城湾监测站点示意图Fig.1 Location of the monitoring site in Great Wall Bay
2.2研究方法
中国第26次和27次南极考察期间,在长城湾布放了一套“极地近岸海海洋环境监测系统”[31](图2)。系统采用坐底式ADCP测量与剖面生态要素测量模式相结合,首次实现了对南极长城湾海域海流特征及海洋生态要素的长期连续监测[19,32]。剖面上层(12.9 m)和中层(19.6 m)配备SEB16 CTD传感器,可以测量温度、盐度、水深、pH、PAR及叶绿素荧光等参数;底层(29.7 m)配备SEB37 CTD传感器,可以测量温度、盐度、水深等参数;坐底平台配备TRDI Workhorse监测型直读式600 kHz ADCP,测量水体流速、流向,数据间隔1 m。系统每10 min采集1组数据,连续监测100 d(2010年12月13日—2011年3月22日),共采集约1.4万组数据(每组数据均包括:温度、盐度、叶绿素a、流速、流向等参数)。
测得的海流数据中含有3种成分:(1)潮流,由引潮力引起的确定周期性运动;(2)余流,由海面风及海上密度场的变化等非引潮力引起的海水运动;(3)湍流等随机扰动有关的随机波动[33]。ADCP获取的数据中包括了东西u(t)、南北v(t)、垂直w(t)3个方向的分量,对任一分量,如u(t)可表示成以下形式:

(1)
式中,a(t)为潮流成分,b(t)表示余流,c(t)为随机扰动,通常将c(t)视为噪声项。
由于观测期间存在一定的不稳定因素,需要在尊重实测数据的前提下进行海流数据质量控制。首先,对原始声学信号特征分析,辨别出错误数据并剔除;然后,通过正态分布规律,剔除均方相对误差超过3倍的测量值,因质量控制缺失的数据,采用线性插值的方法替代;最后,将东西矢量流速u(t)和南北矢量流速v(t)合成,得到水平流速大小和流向。
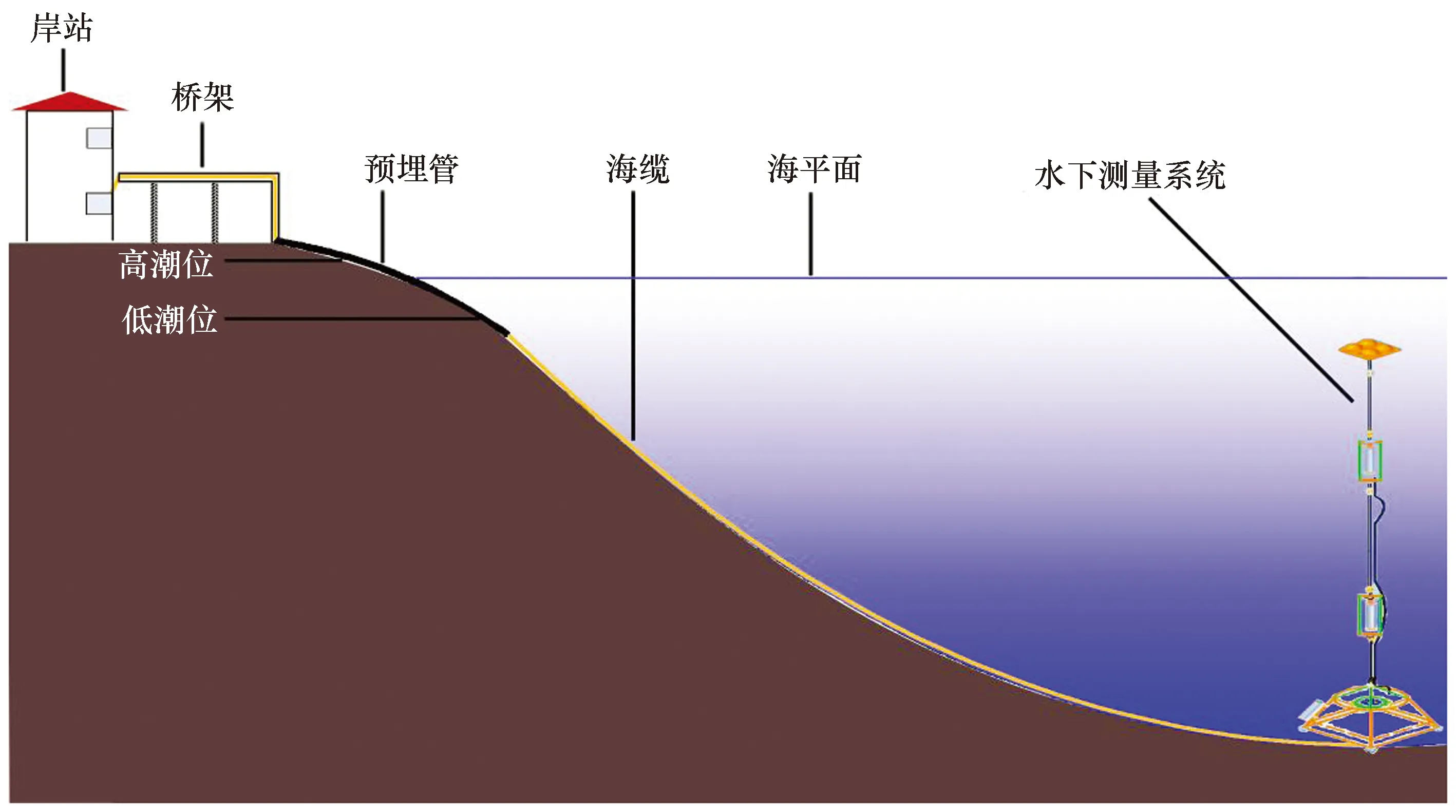
图2 极地近岸海洋环境监测系统示意图Fig.2 The schematic diagram of the polar coastal marine environment monitoring system
3 研究结果
3.1温度、盐度时空变化特征
整个观测期间,随着南半球夏季光照加强、气温升高,3个水层水温均平缓上升(图3)。其中12.9 m水层由开始的0.48℃上升至2011年1月18日的2.52℃,之后在1.5~2.0℃之间波动。19.6 m和29.7 m水层的变化趋势与表层一致,均在2011年1月18日有最大值,分别为2.15℃和2.05℃。3个水层的盐度变化趋势也较为一致,呈现波动递减的趋势,主要集中在33.86~34.19之间(图3),盐度的波动可能与降雨、陆源淡水输入有关。在垂直分布上,温度和盐度均表现出明显的分层现象。
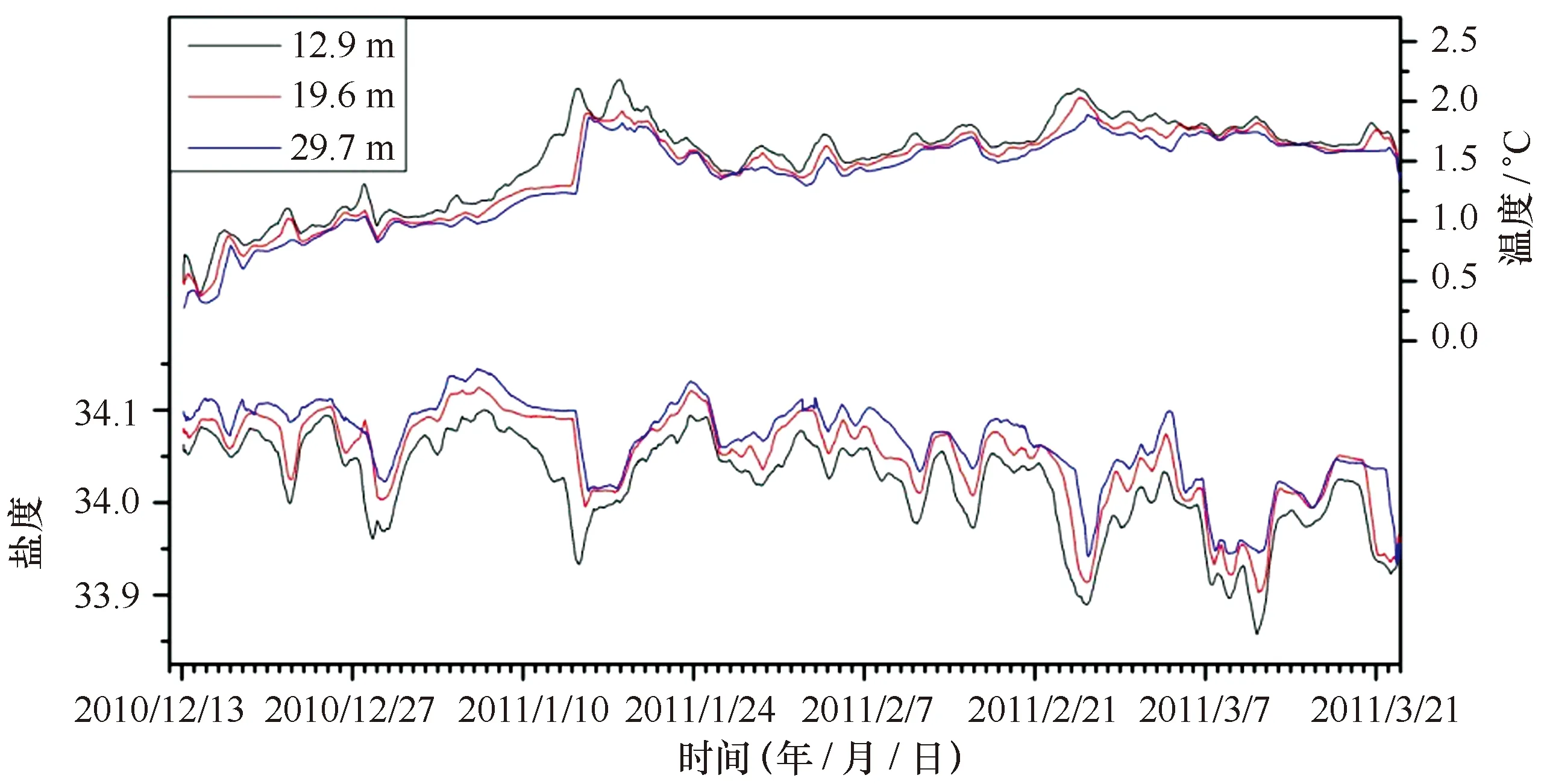
图3 观测期间不同水层水体温度和盐度随时间的变化Fig.3 Seasonal variation of water temperature and salinity in Great Wall Bay 24 h平滑数据 With the data of daily average
3.2叶绿素a浓度变化特征
叶绿素a是海洋浮游植物光合作用的主要色素,其浓度通常被用于表征水体中浮游植物的生物量。在上层水体(水深12.9 m)中,叶绿素a浓度介于0.15~3.76 μg/L之间(图4A),平均为2.25 μg/L。叶绿素a浓度从12月中旬开始增长,直至2月上旬开始维持相对稳定,期间出现两个高值点,分别为2.74 μg/L (2010年12月27日)和3.76 μg/L(2011年1月14日)。中层水体(水深19.6 m)叶绿素a浓度相对较小,介于0.1~1.57 μg/L之间,平均为0.59 μg/L,大部分时段维持在1 μg/L以下,峰值出现在2011年1月16日。
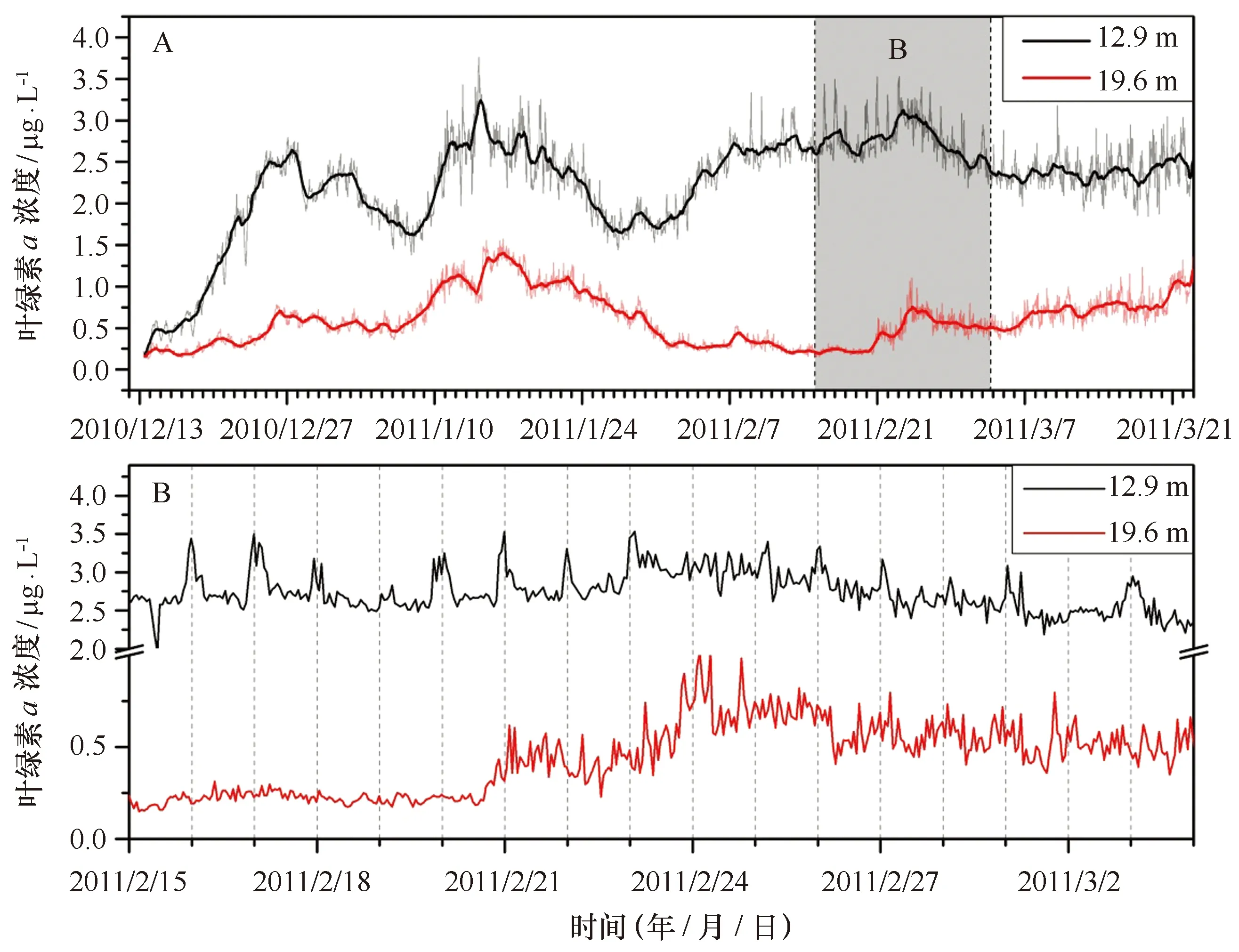
图4 夏季长城湾叶绿素a浓度随时间变化Fig.4 Seasonal and diurnal variations of chlorophyll a concentrations in Great Wall Bay A.24 h平滑数据;B. 2011/2/15至2011/3/3期间1 h间隔数据A.With the data of daily average;B. with the data of hour average from 2011/2/15 to 2011/3/3
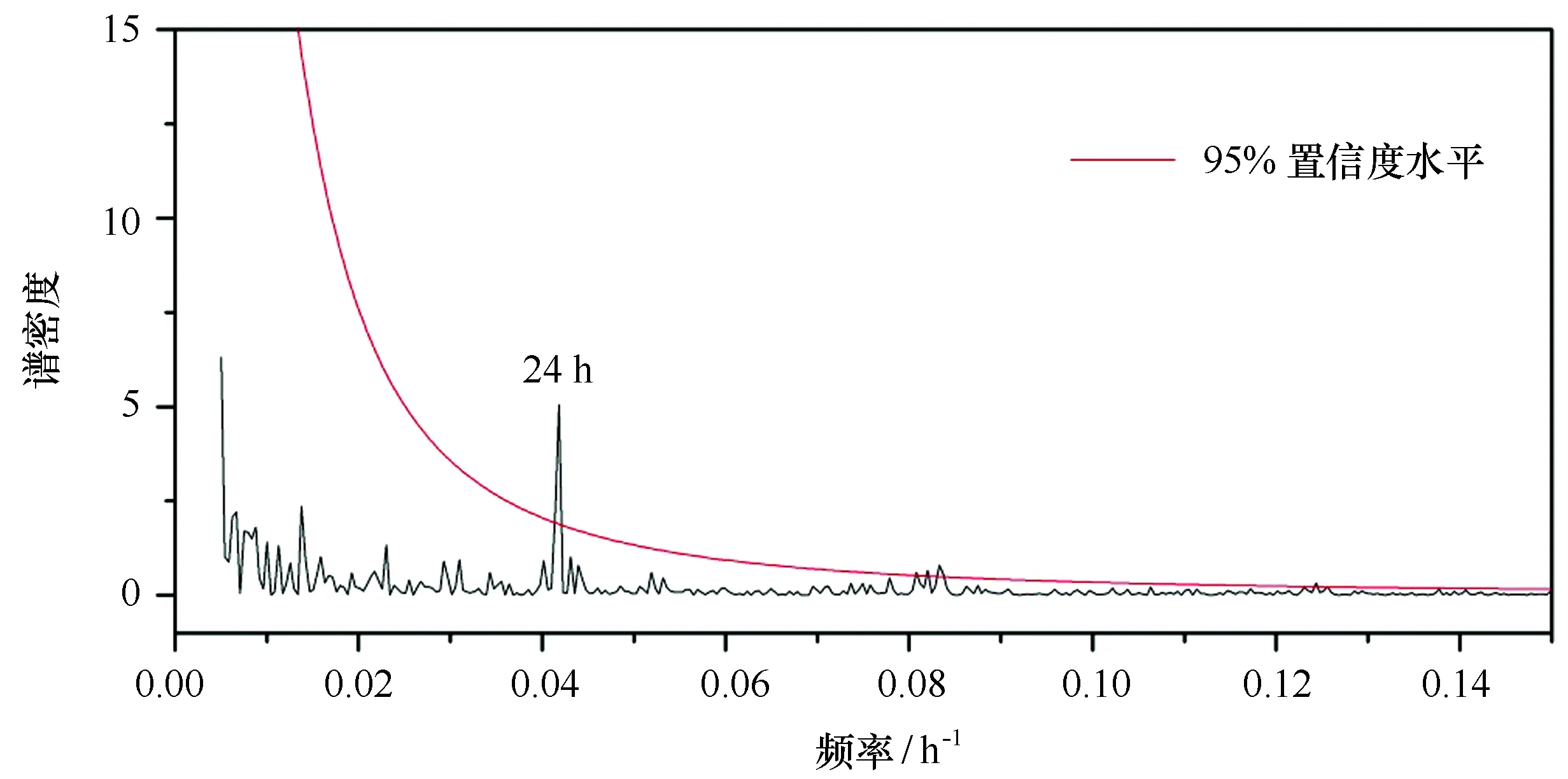
图5 夏季长城湾12.9 m水层叶绿素a浓度频谱分析Fig.5 Power spectra for chlorophyll a concentrations in Great Wall Bay (12.9 m depth)
从叶绿素a浓度随时间分布图(图4B)中可以看出,12.9 m水层的叶绿素a浓度存在明显的周期性变化,而在19.6 m水层中这一现象不明显(本数据记录为北京时间,长城站地区与北京时间相差12 h,标尺刻度为当地时间正午12点位置)。对12.9 m水层的叶绿素a浓度进行频谱分析(图5),结果表明该水层的叶绿素a浓度存在显著的24 h变化周期(通过95%置信度检验)。进一步对观测期间(100 h)叶绿素a浓度每天的最大值和最小值出现的时间段进行统计分析(图6),结果表明每天的叶绿素a浓度最高值出现在中午11:00—12:59(当地时间)时间段内的百分比最高,共有24 d,占总观测天数的24%;其次是15:00—16:59时间段,包含13 d,占总观测天数的13%;在9:00—16:59时间段的8 h中,出现叶绿素a浓度最高值的天数占总观测天数的61%。由此可见,叶绿素a浓度存在24 h周期变化,但是每天变化规律各不相同。每日叶绿素a浓度峰值出现在白天的概率高于夜晚,且出现在正午前后时段的概率要高于其他时段,说明浮游植物的生长、繁殖对光照强度敏感,白天光照强浮游植物的生物量也相对较高;反之,夜晚的生物量相对较低。
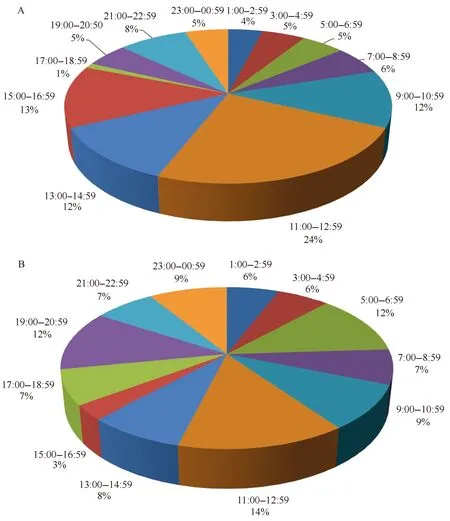
图6 观测期间每日叶绿素a浓度最大值(A)和最小值(B)在各时间段出现的百分比Fig.6 The percentage of daily chlorophyll a maximum (A) and minimum (B) value during different time period
3.3长城湾海流变化特征
观测期间,表层水体的流速相对较大,多数集中在60~100 cm/s之间(图7);底层水体的流速相对较小,约为10 cm/s。时间序列流速剖面分布呈明显的分层特征,其中2010年12月的分界线处在离底15 m处(图7A); 2011年1月的分界线有两段,1月上旬在离底15 m处, 1月中下旬在离底26 m处(图7B) ;2月的分界线也有两段,2月上旬位于离底25 m处,2月中下旬在离底20 m处(图7C);3月的分界线则主要在离底26 m处(图7D)。
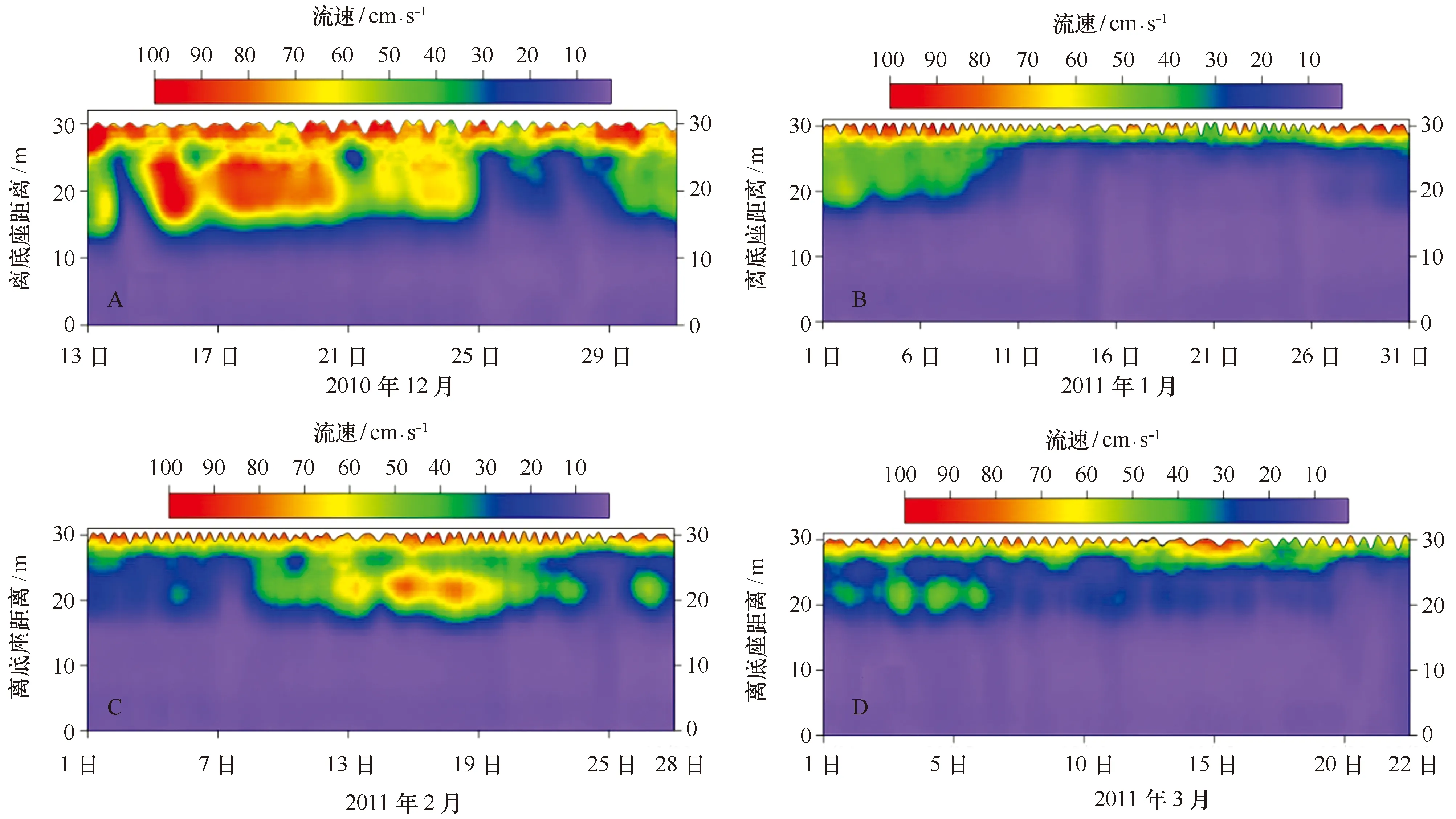
图7 不同月份长城湾水平流速大小随时间的变化Fig.7 Time series of current velocity in Great Wall Bay
对12.9 m水层的流速进行频谱分析(图8),结果表明该水层的流速存在显著的12 h和24 h变化周期(通过95%置信度检验)。实测的海流数据中主要包括潮流、余流和湍流等成分,在近岸海域,潮流是实测海流最重要的成分之一,长城湾的潮汐属于不规则半日潮[18],因此流速存在12 h和24 h变化周期。由于长城湾的风速较大[28],水深较浅,海流受风的影响显著,因此在流速的时间序列图中(图7)周期变化不明显。
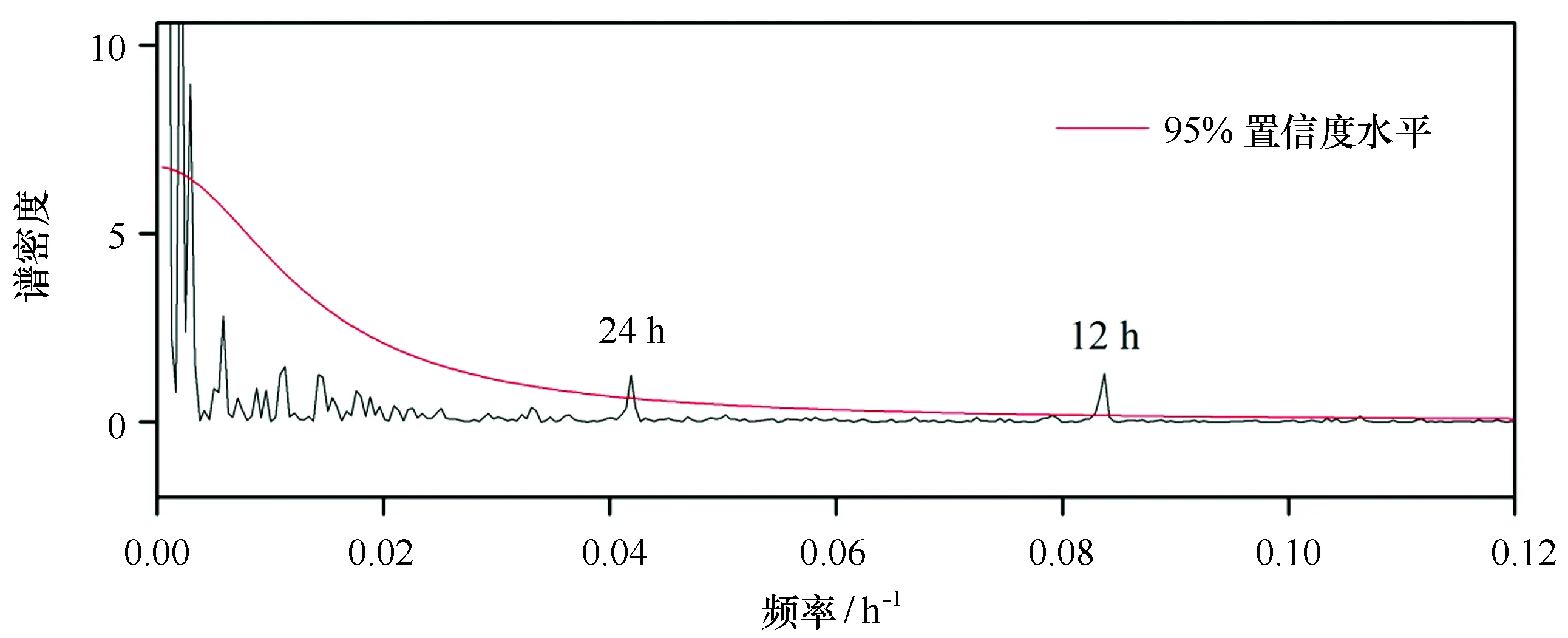
图8 夏季长城湾12.9 m水层流速频谱分析Fig.8 Power spectra for current velocity in Great Wall Bay (12.9 m depth)
3.4长城湾叶绿素a浓度与流速的关系分析
根据观测期间上层(12.9 m)叶绿素a浓度的变化情况,分解成10个阶段(表1),分别与水平流速做相关性分析。从表中可以看出,整个采样期间叶绿素a浓度与水平流速(R2=-0.260,n=14 400,P<0.01)之间存在明显的负相关,且前半个时段的相关性要强于后半个时段。进一步细化分析可以发现,在叶绿素a浓度处于增加的时段相关性强于其他时段。如:2010年12月15—26日,叶绿素a浓度从最初的0.05 μg/L迅速增加至2.74 μg/L,达到第一个峰值,与水平流速表现出显著负相关(R2=-0.595,n=1 584,P<0.01)。这一阶段随着流速的增大,叶绿素a浓度由2.5 μg/L减小至约0.5 μg/L(此时流速约为1 m/s,图9A)。同样,2011年1月7日至1月14日,叶绿素a浓度从1.51 μg/L迅速增加至3.76 μg/L,达到第二个峰值,与水平流速表现出显著负相关(R2=-0.615,n=1 152,P<0.01)。这一阶段水平流速小于0.1 m/s时,叶绿素a浓度集中在2~4 μg/L,且流速越小浓度越高;而水平流速大于0.2 m/s时,叶绿素a浓度维持在1.6 μg/L(图9B)。由此可见,流速在一定程度上会影响长城湾浮游植物的生物量。
表1不同采样时段叶绿素a浓度与流速的相关性分析
Tab.1Correlation between chlorophyllaconcentrations and current velocities within upper water column during different timescales
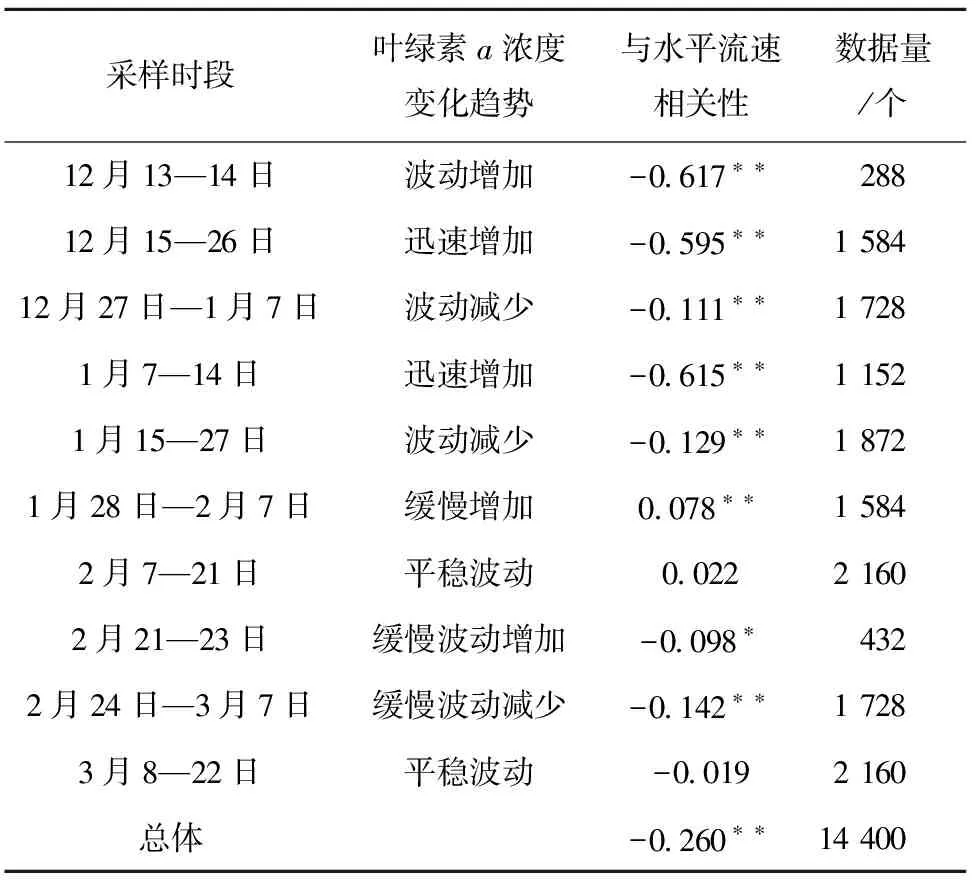
注:**表示极显著相关(P<0.01); *表示显著相关(P<0.05)。
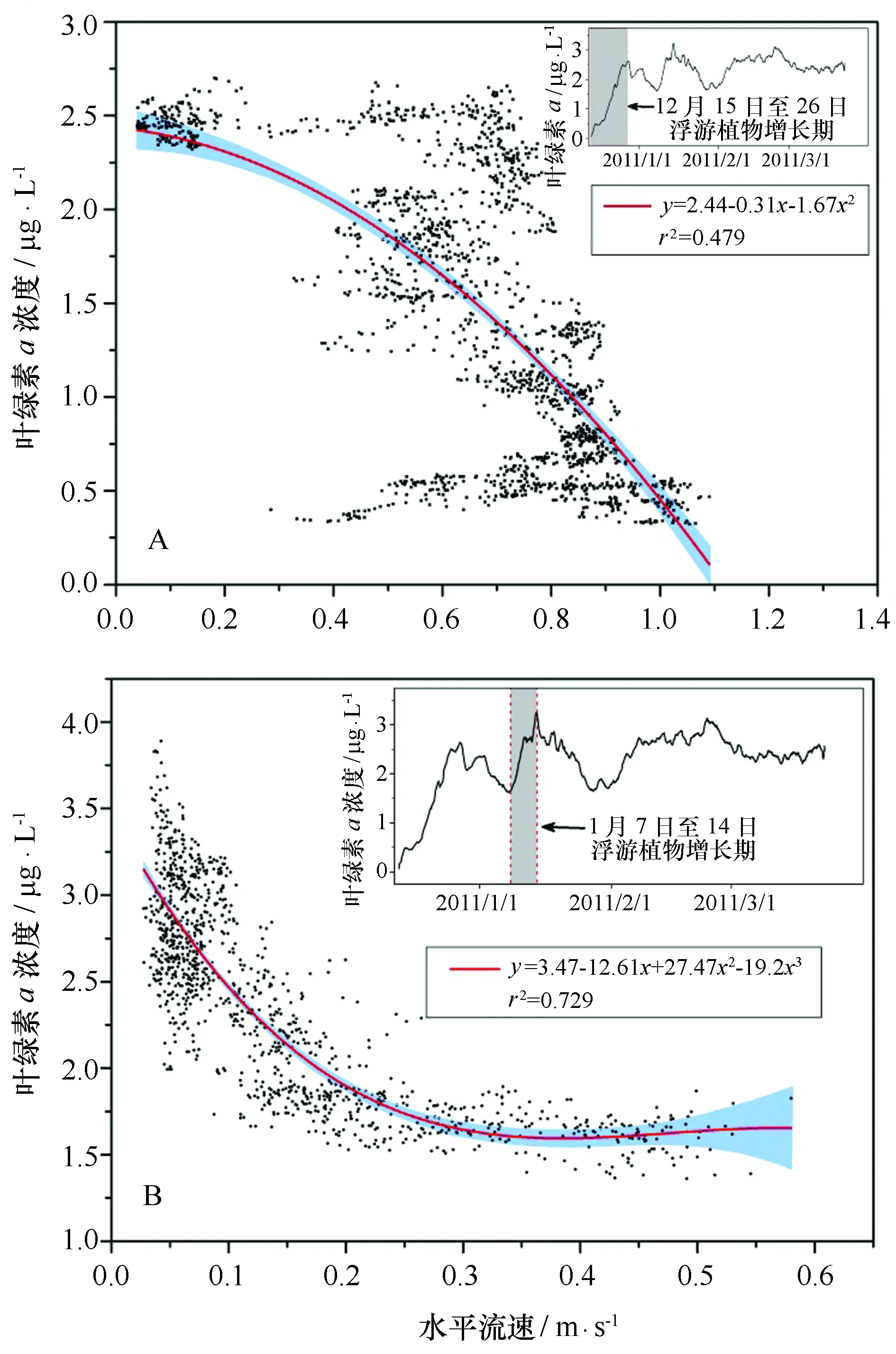
图9 浮游植物增长期叶绿素a浓度与水平流速相关性Fig.9 Fit of chlorophyll a in relation to current velocity蓝色阴影部分为95%置信区间。A为2010年12月15—26日; B为2011年1月7—14日The shaded areas of blue represent 95% confidence bands. A. from 2010/12/15 to 12/26, B. from 2011/1/7 to 1/14
4 讨论
4.1长城湾的水交换特征
长城湾所处区域盛行西风[28],表层海流受短时风况的影响显著(图10),在风的作用下表层流向随风向而改变。从12月至次年3月,长城站附近主要以西北风为主,平均风速达7 m/s,期间出现短暂几天的东南风(12月15、25和27日,1月13—20日,2月7、13和21日,3月9、20—22日)。表层流向与风向表现出显著的同步变化规律,西北风出现时表层海水流向湾外,东南风出现时湾外海水涌入长城湾。表层海水受风的影响流速较大,而底层靠近海底,摩擦力作用会导致流速减缓[21]。
从长城湾海流的流向分布可以看出: 12月(图10)表层(离底25~30 m层)流向约为135°,水流从长城湾流向麦克斯韦尔湾;而中层(离底10~25 m层)流向分布在315°~360°,水流从麦克斯韦尔湾流进长城湾;底层(离底0~10 m层)水体的流向分布在135°~180°,长城湾底层水体缓慢地从湾内流向麦克斯韦尔湾。1月份(图10),由外部流向长城湾的水流变得很薄(5~8 m),随着夏季温度的升高,降雨、陆源水等输入长城湾,补给了湾内的水量,加强了长城湾的水体向外输送。2月和3月(图10),外部流入水体由薄变厚,说明夏末气温趋于平稳,长城湾的降雨、陆源水输入减少,湾外海水又重新进入长城湾。将各月海流中的潮流成分剥离后得到各月的余流分布(图11)。长城湾表层余流与中、底层余流呈反向分布。其中表层余流方向为东南向(约为135°),中层则为西北向(约为315°)。从12月至3月东南方向的表层余流逐渐变厚(5 m增加到10 m),速度也增大,从12月的约20 cm/s增加至3月的约30 cm/s。中、底层余流的大小与方向在整个夏季的变化不大,以西北向为主,速度约为15 cm/s。由此可见,长城湾与麦克斯韦尔湾可以发生水体交换,水体主要从中层进入湾内,由表层流出。
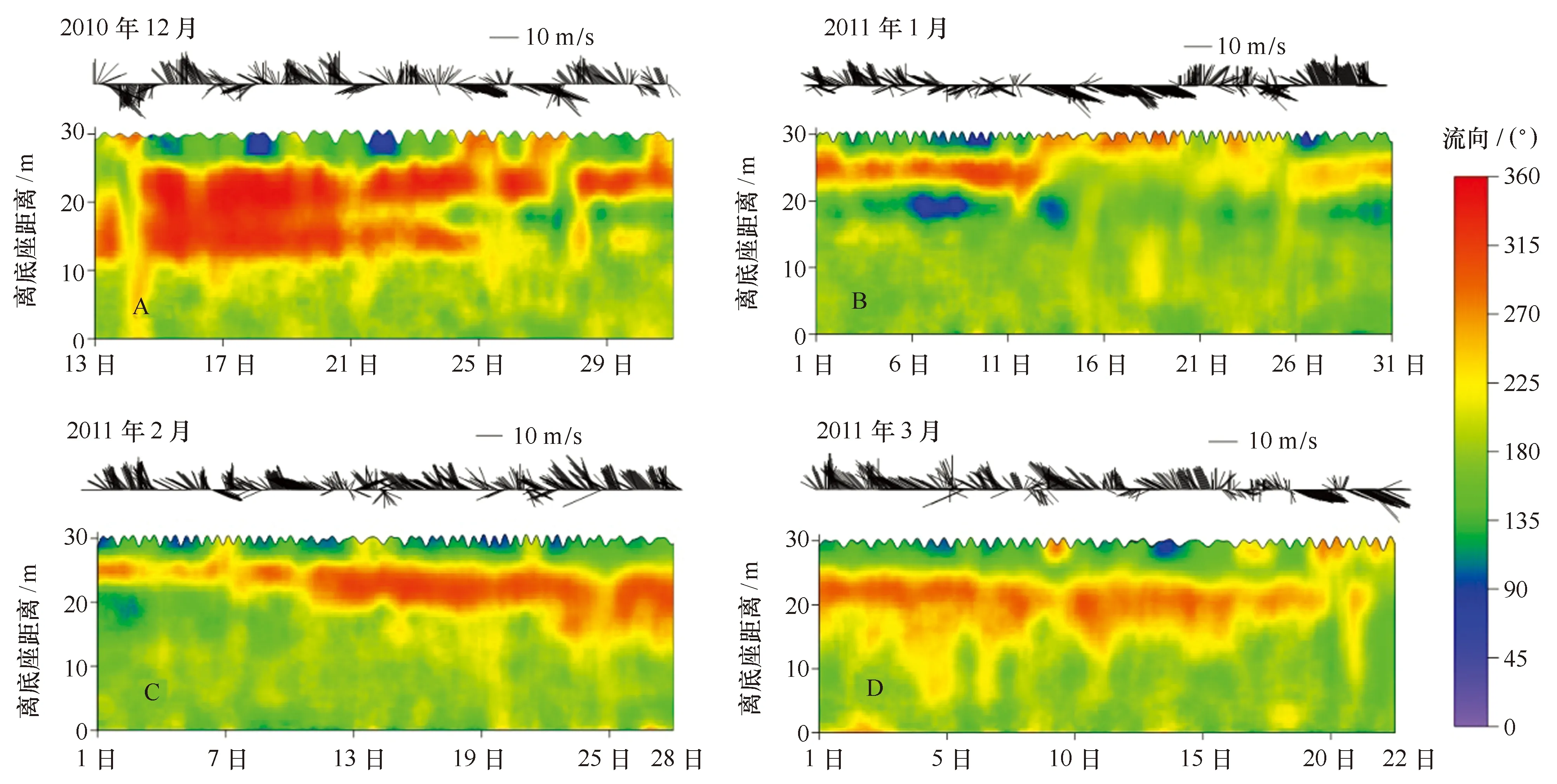
图10 不同月份长城湾风场及海流流向分布图Fig.10 Time series of wind and current direction in Great Wall Bay
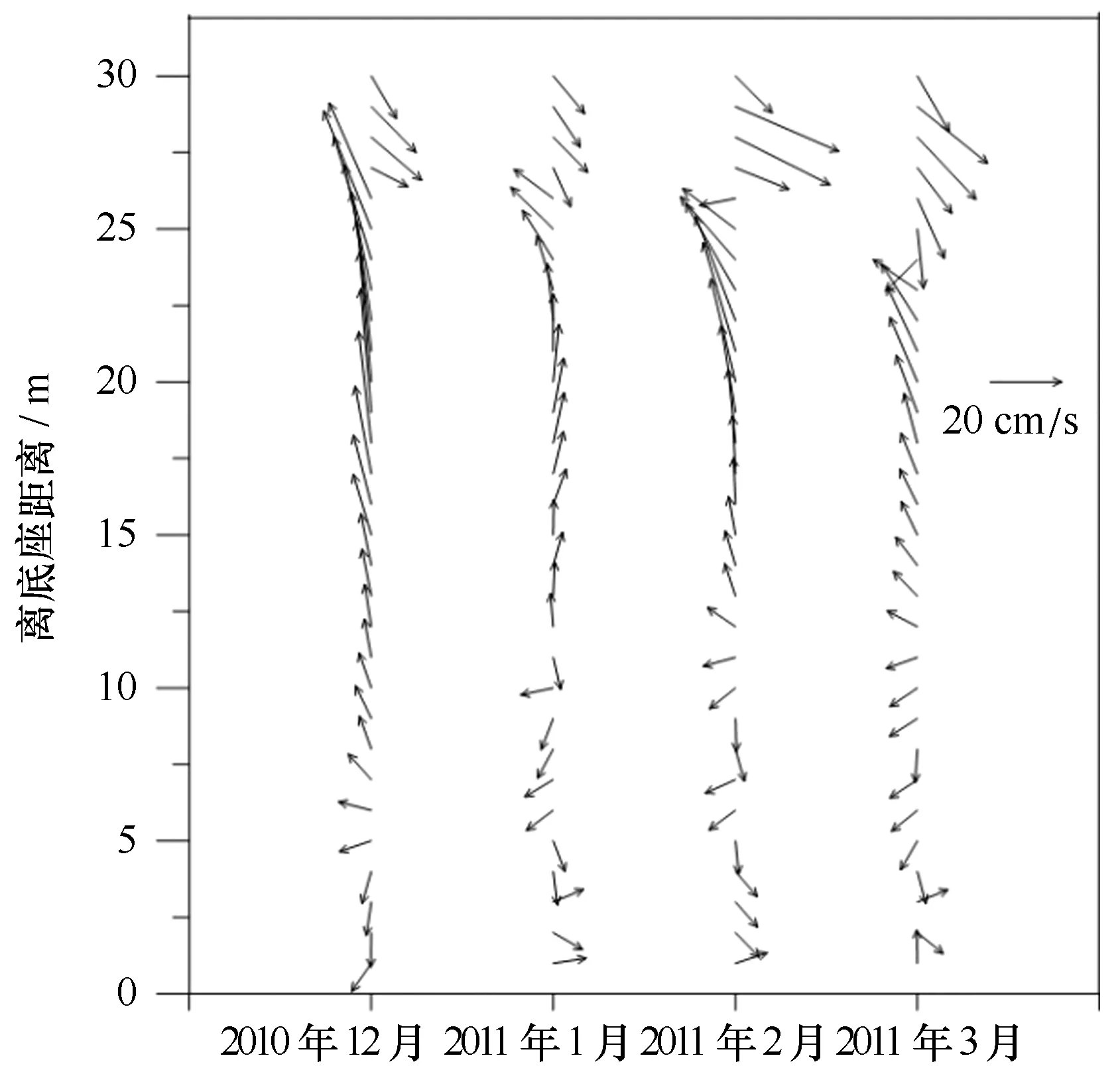
图11 夏季长城湾月平均余流分布Fig.11 Distributions of residual current in summer
此外,风海流的体积运输会导致海水在海湾发生辐散或者辐聚,由于连续运输又引起海水产生上升或下沉运动[21]。24年的长城站风向资料显示,长城湾北到偏西方向(270°~360°)的风向频率在75%以上[28],说明大部分时间风向与长城湾的两岸平行,使表层海水产生离岸的运动(即辐散),因此下层海水涌升到上层,驱动了长城湾水体的垂直交换。
综上所述,长城湾水体的来源主要包括麦克斯韦尔湾输入、陆地径流输入及少量浮冰融水和降雪降雨输入,输出途径主要为表层输出及少量的蒸发和下渗作用(图12)。麦克斯韦尔湾水主要从湾口断面的中层进入长城湾,由于辐散作用,水体在进入过程中有较弱的上升运动,表层水体可以通过风生海流作用向湾外输送。因此,夏季长城湾水体可以与湾外水体持续保持交换。
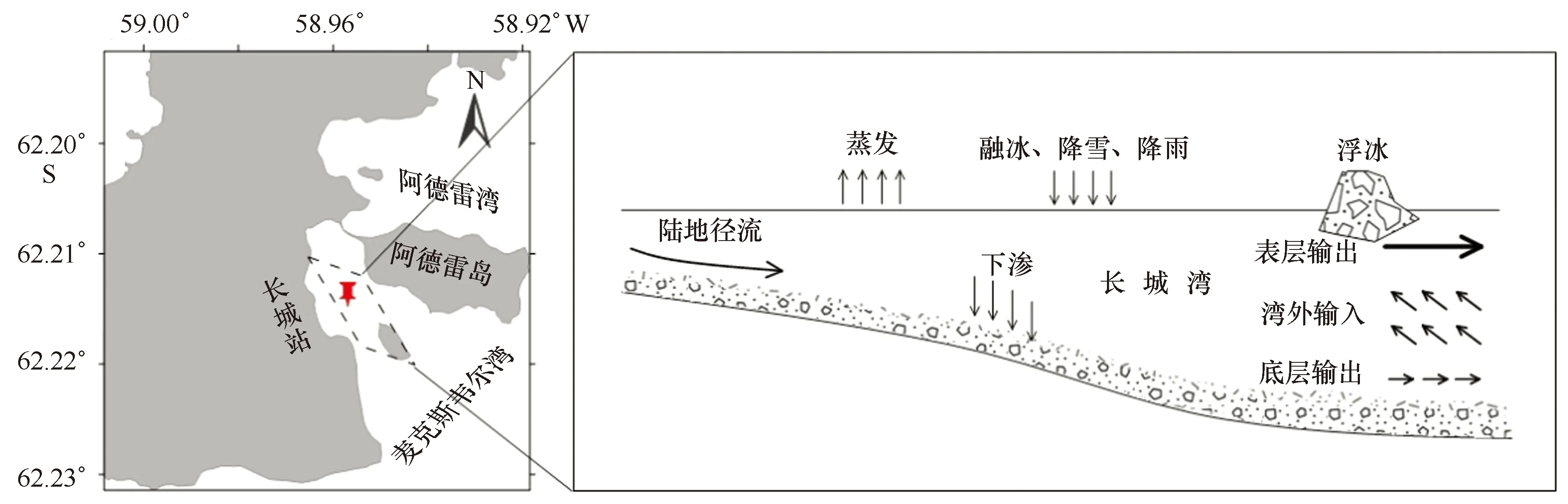
图12 长城湾水交换简易模式图Fig.12 The simplified pattern of water exchange in Great Wall Bay
4.2海流对浮游植物的影响
海流对浮游生物最重要、最直接的影响在于海流播散和维持生物群的作用[20]。海流的运输作用可以将开放海域或邻近海域的浮游植物类群引入长城湾中,从而影响浮游植物的群落结构和生物量。李瑞香等[17]的研究表明,聚生角刺藻、无刺翼根管藻、脆杆藻等是长城湾历年夏季的优势种,但是在湾口也常出现扁面角刺藻等一些重现性较差的种类,这可能与湾外海流有关。长城湾的东南方向是麦克斯韦尔湾,而麦克斯韦尔湾与布兰斯菲尔德海峡相连,受湾外水流的影响,长城湾湾口的叶绿素a浓度低于湾内水体[34]。在风浪的作用下,表层海水很不稳定,不利于浮游植物的生存和停留,造成长城湾中层和底层水体中的叶绿素a含量高于表层[13]。各种海流交汇的海洋锋引起海水的辐散、辐聚和混合,这些过程控制着初级生产的营养盐供应,进而对海水中浮游植物的种类组成和分布产生影响[20]。Elliott和Defew[22]发现,流速增加可以带来更多的营养盐,促进一些种类浮游植物的增长,同时也对其他一些种类的浮游植物增长有负面影响。海流带来的水体扰动和沉积物再悬浮也能使浮游植物的群落结构发生改变,例如在南极阿德默勒尔蒂湾(Admiralty Bay),夏初水体混合引起沉积物再悬浮,使底栖羽纹硅藻成为优势种;夏末则受湾外水体的影响,中心硅藻变成优势种[35]。
流速会影响浮游植物的生物量。本研究中长城湾叶绿素a浓度的第二个峰值期间(2011年1月7—14日),水平流速小于0.1 m/s时,叶绿素a浓度集中在2~4 μg/L,且流速越小浓度越高;而水平流速大于0.2 m/s时,叶绿素a浓度维持在1.6 μg/L。由长城湾叶绿素a浓度与流速的相关性分析可知(表1,图9),叶绿素a浓度与流速呈显著负相关,说明低流速环境有利于长城湾的浮游植物生长,反之高流速环境则不利于浮游植物生长。在加利福尼亚北部沿岸海域,Hood等[36]研究了20 m水深处的叶绿素a浓度与流速关系,发现二者显著相关,流速急的海区叶绿素a浓度低,流速缓慢的海区叶绿素a浓度高。D’Amour和Scheibling[26]的水槽实验表明,不同水流作用(0.3~0.5 m/s)产生的拖拽力对绿藻的生长有影响,0.5 m/s是其临界流速。Long等[25]建立生态动力学模型分析流速对藻类增长的影响,结果显示大约0.04 m/s流速是浮游藻类生长的最佳流速。由此可见,不同的浮游植物类群对流速的耐受限度也不同,符合谢尔福德耐受性定律[20],即当流速处于某一范围时,对有些种类浮游植物生长有利,而对另一些种类浮游植物却有抑制作用。对某一种浮游植物而言,流速太高或者太低都会影响其生存与繁殖。
5 结论
通过分析长城湾夏季水体中叶绿素a浓度和海流变化数据,探讨了长城湾水交换特征及海流对浮游植物的影响,得到以下几点结论:
(1)夏季长城湾水体中叶绿素a浓度上层(12.9 m)高于下层(19.6 m),二者浓度范围分别为0.05~3.76 μg/L和0.13~1.57 μg/L。上层水体叶绿素a浓度从12月中旬开始增长,直至2月上旬开始维持相对稳定,期间出现两个高值点(2.74 μg/L 和3.76 μg/L);下层水体叶绿素a浓度也从12月中旬开始增长,峰值出现在1月中旬,为1.57 μg/L,随后从2月初开始维持相对稳定,约为1 μg/L。叶绿素a浓度存在24 h周期变化,但是每天变化规律各不相同,正午前后时段出现叶绿素a峰值的概率高于其他时段,峰值出现在白天的概率高于夜晚。
(2)观测期间,各月的表层水平流速总体较大,主要集中在60~100 cm/s之间;中层和底层水体的流速都较小,约为10 cm/s。表层海流受短时风况的影响显著,使长城湾海水发生辐散,下层海水向上涌升,驱动了长城湾水体的垂直交换。长城湾与麦克斯韦尔湾存在水体交换,水体主要从湾口断面的中层进入湾内,由表层流出。
(3)长城湾的叶绿素a浓度与流速呈显著负相关,表明不同的浮游植物类群对流速的耐受限度也不同,低流速环境有利于长城湾浮游植物类群的生长。海流可以将其他海域的浮游植物种类引入长城湾,也可以将长城湾的浮游植物往湾外搬运,使浮游植物群落结构发生改变。在今后对长城湾浮游植物群落的研究中需要考虑海流对其的潜在影响。
致谢:对所有参与监测系统回收与布放的极地考察队员表示衷心感谢!同时感谢审稿专家提出的宝贵意见!
[1]Thomas M K, Kremer C T, Klausmeier C A, et al. A global pattern of thermal adaptation in marine phytoplankton[J]. Science, 2012, 338(6110): 1085-1088.
[2]Burrows M T, Schoeman D S, Buckley L B, et al. The pace of shifting climate in marine and terrestrial ecosystems[J]. Science, 2011, 334(6056): 652-655.
[3]Forcada J, Hoffman J I. Climate change selects for heterozygosity in a declining fur seal population[J]. Nature, 2014, 511(7510): 462-465.
[4]Abram N J, Mulvaney R, Wolff E W, et al. Acceleration of snow melt in an Antarctic Peninsula ice core during the twentieth century[J]. Nature Geoscience, 2013, 6(5): 404-411.
[5]Chen J L, Wilson C R, Tapley B D. Contribution of ice sheet and mountain glacier melt to recent sea level rise[J]. Nature Geoscience, 2013, 6(7): 549-552.
[6]Shevenell A E, Ingalls A E, Domack E W, et al. Holocene Southern Ocean surface temperature variability west of the Antarctic Peninsula[J]. Nature, 2011, 470(7333): 250-254.
[7]Mulvaney R, Abram N J, Hindmarsh R C A, et al. Recent Antarctic Peninsula warming relative to Holocene climate and ice-shelf history[J]. Nature, 2012, 489(7414): 141-144.
[8]Boyce D G, Dowd M, Lewis M R, et al. Estimating global chlorophyll changes over the past century[J]. Progress in Oceanography, 2014, 122: 163-173.
[9]McQuatters-Gollop A, Reid P C, Edwards M, et al. Is there a decline in marine phytoplankton?[J]. Nature, 2011, 472(7342): E6-E7.
[10]Boyce D G, Lewis M R, Worm B. Global phytoplankton decline over the past century[J]. Nature, 2010, 466(7306): 591-596.
[11]Field C B, Behrenfeld M J, Randerson J T, et al. Primary production of the biosphere: integrating terrestrial and oceanic components[J]. Science, 1998, 281(5374): 237-240.
[12]俞建銮, 李瑞香, 黄凤鹏. 南极长城湾浮游植物生态的初步研究[J]. 南极研究, 1992, 4(4): 34-39.
Yu Jianluan, Li Ruixiang, Huan Fengpeng. A preliminary study on the ecology of the phytoplankton in Great Wall Bay, Antarctica[J]. Antarctic Research, 1992, 4(4): 34-39.
[13]朱明远, 李宝华, 黄凤鹏, 等. 南极长城湾夏季叶绿素a变化的研究[J]. 极地研究, 1999, 11(2): 113-121.
Zhu Mingyuan, Li Baohua, Huang Fengpeng, et al. A study on cholorophyllacontent and its variation in Great Wall Bay, Antarctica during the austral summers from 1992 to 1995[J]. Chinese Journal of Polar Research, 1999, 11(2): 113-121.
[14]李宝华. 南极长城站码头及临近海域夏季叶绿素a含量及变化[J]. 极地研究, 2004, 16(4): 332-337.
Li Baohua. Variation of chlorophyllacontents at the wharf of the Great Wall Station, Antarctica[J]. Chinese Journal of Polar Research, 2004, 16(4): 332-337.
[15]李宝华. 南极长城站及邻近海域叶绿素a次表层极大值的研究[J]. 极地研究, 2004, 16(2): 127-134.
Li Baohua. The subaltern of chlorophyllamaxima in the Great Wall Bay and adjacent sea, Antarctica[J]. Chinese Journal of Polar Research, 2004, 16(2): 127-134.
[16]何剑锋, 张芳, 蔡明红, 等. 基于极地近岸海洋环境监测系统的南极长城湾生态环境变化初析[J]. 极地研究, 2011, 23(4): 269-274.
He Jianfeng, Zhang Fang, Cai Minghong, et al. Basic analysis of the seasonal environmental variation at Great Wall Bay, Antarctica, using a nearshore marine enviromental monitoring system[J]. Chinese Journal of Polar Research, 2011, 23(4): 269-274.
[17]李瑞香, 朱明远, 洪旭光. 南极长城湾夏季浮游植物数量与环境的关系[J]. 黄渤海海洋, 2001, 19(3): 71-75.
Li Ruixiang, Zhu Mingyuan, Hong Xuguang. Relationship between the phytoplankton abundance and environment in the Great Wall Bay, Antarctica[J]. Journal of Oceangraphy of Huanghai & Bohai Seas, 2001, 19(3): 71-75.
[18]孙洪亮. 南极长城站潮汐变化特征及水文气象因素对平均水位的影响[J]. 海洋学报, 1992, 14(4): 112-123.
Sun Hongliang. Variation of the tide at the Great Wall Station and the effect of hydrometeorological factor on mean water level[J]. Haiyang Xuebao, 1992, 14(4): 112-123.
[19]Ma Yuxin, Zhang Fang, Yang Haizhen, et al. Detection of phytoplankton blooms in Antarctic coastal water with an online mooring system during summer 2010/11[J]. Antarctic Science, 2014, 26(3): 231-238.
[20]沈国英, 黄凌风, 郭丰, 等. 海洋生态学[M]. 3版. 北京: 科学出版社, 2010.
Shen Guoying, Huang Lingfeng, Guo Feng, et al. Marine Ecology[M]. 3rd ed. Beijing: Science Press, 2010.
[21]冯士筰, 李凤岐, 李少菁. 海洋科学导论[M]. 北京: 高等教育出版社, 1999.
Feng Shizuo, Li Fengqi, Li Shaojing. An Introduction to Marine Science[M]. Beijing: Higher Education Press, 1999.
[22]Elliott J A, Defew L. Modelling the response of phytoplankton in a shallow lake (Loch Leven, UK) to changes in lake retention time and water temperature[J]. Hydrobiologia, 2012, 681(1): 105-116.
[23]Lehman J T, Platte R A, Ferris J A. Role of hydrology in development of a vernal clear water phase in an urban impoundment[J]. Freshwater Biology, 2007, 52(9): 1773-1781.
[24]Mitrovic S M, Oliver R L, Rees C, et al. Critical flow velocities for the growth and dominance ofAnabaenacircinalisin some turbid freshwater rivers[J]. Freshwater Biology, 2003, 48(1): 164-174.
[25]Long Tianyu, Wu Lei, Meng Guohu, et al. Numerical simulation for impacts of hydrodynamic conditions on algae growth in Chongqing Section of Jialing River, China[J]. Ecological Modelling, 2011, 222(1): 112-119.
[26]D’Amours O, Scheibling R E. Effect of wave exposure on morphology, attachment strength and survival of the invasive green algaCodiumfragilessp.tomentosoides[J]. Journal of Experimental Marine Biology and Ecology, 2007, 351(1/2): 129-142.
[27]Li Feipeng, Zhang Haiping, Zhu Yiping, et al. Effect of flow velocity on phytoplankton biomass and composition in a freshwater lake[J]. Science of the Total Environment, 2013, 447: 64-71.
[28]卞林根, 马永锋, 逯昌贵, 等. 南极长城站(1985-2008)和中山站(1989-2008)地面温度变化[J]. 极地研究, 2010, 22(1): 1-9.
Bian Lin’gen, Ma Yongfeng, Lu Changgui, et al. Temperature variations at the Great Wall Station (1985-2008) and Zhongshan Station (1989-2008), Antarctic[J]. Chinese Journal of Polar Research, 2010, 22(1): 1-9.
[29]秦大河. 南极洲乔治王岛长城湾一年生海冰的发育特征和物理性质[J]. 冰川冻土, 1991, 13(2): 115-130.
Qin Dahe. Developing and physical characteristics of first-year sea ice in Great Wall Bay and its adjacent area by King George Island, Antarctica[J]. Journal of Glaciology and Geocryology, 1991, 13(2): 115-130.
[30]杨清华, 张本正, 李明, 等. 2012年南极长城站气象和海冰特征分析[J]. 极地研究, 2013, 25(3): 268-277.
Yang Qinghua, Zhang Benzheng, Li Ming, et al. Analysis of weather and sea ice at the Antarctic Great Wall Station in 2012[J]. Chinese Journal of Polar Research, 2013, 25(3): 268-277.
[31]何剑锋, 张芳, 林凌. 我国极地海底观测系统的发展与展望[J]. 地球科学进展, 2013, 28(5): 566-571.
He Jianfeng, Zhang Fang, Lin Ling. Development and prospect of Chinese polar seabed observation systems[J]. Advances in Earth Science, 2013, 28(5): 566-571.
[32]He Jianfeng, Zhang Fang, Lin Ling, et al. Effects of the 2010 Chile and 2011 Japan tsunamis on the Antarctic coastal waters as detected via online mooring system[J]. Antarctic Science, 2012, 24(6): 665-671.
[33]王辉武, 陈红霞, 吕连港, 等. 楚科奇海夏季潮流和余流观测研究[J]. 海洋学报, 2011, 33(6): 1-8.
Wang Huiwu, Chen Hongxia, Lv Lian’gang, et al. Study of tide and residual current observations in Chukchi Sea in the summer 2008[J]. Haiyang Xuebao, 2011, 33(6): 1-8.
[34]吕培顶, 渡边研太郎. 1988/1989年夏季南极长城海湾生态环境观测[J]. 南极研究, 1994, 6(3): 62-73.
Lü Peiding, Watanabe K. Observations on ecological environmental condition of the Great Wall Bay, King George Island, Antarctica, from November 1988 to March 1989[J]. Antarctic Research, 1994, 6(3): 62-73.
[35]Lange P K, Tenenbaum D R, de Santis Braga E, et al. Microphytoplankton assemblages in shallow waters at Admiralty Bay (King George Island, Antarctica) during the summer 2002-2003[J]. Polar Biology, 2007, 30(11): 1483-1492.
[36]Hood R R, Abbott M R, Huyer A, et al. Surface patterns in temperature, flow, phytoplankton biomass, and species composition in the coastal transition zone off Northern California[J]. Journal of Geophysical Research: Oceans, 1990, 95(C10): 18081-18094.
Effects of current velocity on phytoplankton biomass in Great Wall Bay, Antarctica during summer 2010/2011
Luo Guangfu1, He Jianfeng1,2, Zhang Fang1, Cai Minghong1, Lin Ling1, Lan Musheng1
(1. The Key Laboratory for Polar Science of State Ocean Administration, Polar Research Institute of China, Shanghai 200136, China;2. College of Environmental Science and Engineering, Tongji University, Shanghai 200092, China)
Understanding the mechanism by which environmental factors affects phytoplankton growth in summer is essential for determining polar marine ecosystem responses to global climate change. Here we deployed an online mooring system in Great Wall Bay to determine how current velocity and local physical forcing affect the temporal and spatial distribution of phytoplankton biomass in Antarctic coastal waters. The results show that chlorophylla(Chla)concentrations accumulated from mid-December and two significant blooms developed in January (2.74 μg/L and 3.76 μg/L) and were then maintained at a relatively high level. The diurnal variations of cholorophyllaconcentrations were observed in both upper and middle layers during the monitoring period. The surface current velocity reached approximately (60-100)cm/s, while the middle and bottom current velocity was limited to about 10 cm/s. We conclude that summer winds played an important role in the surface current velocity and direction. The monitoring site appears to exchange with adjacent oceanic waters, such an intrusion of water would bring in varied phytoplankton species. The results of Pearson correlation analysis showed the significant and positive influence of low current velocity on chlorophyllaconcentrations, especially during phytoplankton blooms, whereas the increased current velocity showed the significant and negative correlations with phytoplankton biomass.
chlorophyllaconcentrations; current; phytoplankton biomass; Great Wall Bay
10.3969/j.issn.0253-4193.2016.10.012
2016-01-11;
2016-05-05。
南北极环境综合考察与评估专项(CHINARE2016-01-05);国家高技术研究发展计划(2007AA09Z121)。
罗光富(1989—),男,福建省宁化县人,从事极地海洋生态研究。E-mail:luoguangfu@pric.org.cn
何剑锋,男,研究员,从事极地海洋生态研究。E-mail:hejianfeng@pric.org.cn
Q958.8
A
0253-4193(2016)10-0113-12
罗光富,何剑锋,张芳,等. 夏季南极长城湾海流变化特征及其对浮游植物生物量的影响[J].海洋学报,2016,38(10):113-124,
Luo Guangfu, He Jianfeng, Zhang Fang, et al. Effects of current velocity on phytoplankton biomass in Great Wall Bay, Antarctica during summer 2010/2011[J]. Haiyang Xuebao,2016,38(10):113-124, doi:10.3969/j.issn.0253-4193.2016.10.012
