春季我国不同纬度河口浮游动物群落变化趋势
2016-11-11卢伍阳马增岭徐兆礼高倩
卢伍阳,马增岭,徐兆礼,高倩
(1.中国水产科学研究院 东海水产研究所 农业部海洋与河口渔业重点开放实验室,上海 200090;2.上海海洋大学 海洋科学学院,上海 201306;3.温州大学 生命与环境科学学院,浙江 温州 325035)
春季我国不同纬度河口浮游动物群落变化趋势
卢伍阳1,2,马增岭3,徐兆礼1*,高倩1
(1.中国水产科学研究院 东海水产研究所 农业部海洋与河口渔业重点开放实验室,上海 200090;2.上海海洋大学 海洋科学学院,上海 201306;3.温州大学 生命与环境科学学院,浙江 温州 325035)
根据2010-2012年春季由南向北河口海域:北仑河口、九龙江口、瓯江口、长江口和灌河口的浮游动物调查资料,通过比较不同纬度河口浮游动物群落的生物多样性指数(H′)、物种更替率(R)和群落相似系数(S),探讨了河口浮游动物群落特征沿着不同纬度变化的趋势。结果表明,自南向北5个河口可以划分为3个不同的浮游动物区系。北仑河口和九龙江口为南部亚热带区系,位于亚热带海域,以亚热带种为主,H′分别为1.01和2.18,两河口之间R值为65.52%,S值为51.28%;瓯江口和长江口为中部过渡区系,位于亚热带和温带过渡海域,亚热带种和暖温带种是主要出现的种类,H′分别为1.31和1.86,两河口之间R值为58.33%,S值为58.82%;灌河口单独为北部温带区系,位于温带海域,暖温带种是主要出现的种类,H′值为1.19,与其余河口物种更替率均高于80.00%,群落相似系数均低于32.00%;表明群落结构由南到北的变化非常显著。线性回归结果表明:我国不同河口之间浮游动物的物种更替率与纬度差(两河口所处纬度的差值,ΔN)呈正相关关系(R=0.026ΔN+0.608,p=0.002);群落相似系数与纬度差呈负相关关系(S=0.034ΔN+0.578,p=0.001)。春季,我国不同纬度河口浮游动物群落结构的变化,主要是因为影响各河口的水温和水团存在较大差别所致。
河口;浮游动物;群落;纬度;变化
1 引言
在我国,从南到北共有1 500多条江河注入近海[1],形成的河口,纵跨热带、亚热带和温带等气候带,分布于18°~41°N之间;南北不同纬度、不同气候带河口的海域环境有可能造成浮游动物群落形成较大差异。作为河口生态系统中重要的生物类群,浮游动物也是环境和气候变化的指示生物[2—3]。河口浮游动物群落结构的变化与海洋气候和环境具有较高的耦合性,故对不同纬度河口浮游动物的群落变化趋势进行分析就显得特别重要。
春季,我国近海由北风转向南风、温度回升[4];然而这一过程由南向北展开,不同纬度气候环境分化,使得我国从南到北不同河口海域环境呈现不同的格局,由此浮游动物群落结构形成相应的变化。目前,我国对春季河口浮游动物的研究主要局限于单一河口海域,研究内容多是浮游动物的群落结构[5],空间分布[6]和季节更替[7]等方面。对于我国近海多个河口(两个及以上)浮游动物的研究,陈剑等[8]比较了夏季东南沿海的闽江口和椒江口浮游动物群落结构的差异;张达娟等[9]分析了黄河口、长江口和珠江口的浮游动物种类数和生物量的变化趋势,但均没有研究不同纬度之间多个河口的群落结构变化。在国外,这类跨大尺度纬度的研究也不多见。Kibirige等[10]研究了南非两个相邻亚热带河口在不同时期微型浮游动物对浮游动物群落结构的影响;Sautour和Castel[11]对春季欧洲西南部相距不远的3个河口浮游动物分布进行了研究,这些研究都局限在一个较小的范围。
本文基于2010-2012年北仑河口、九龙江口、瓯江口、长江口和灌河口等5个河口浮游动物现场调查资料,对春季不同纬度河口之间浮游动物群落划分和变化进行研究;以期了解我国海域气候环境对浮游动物群落多样性的影响及河口浮游动物对环境的指示作用,并有助于认识我国不同纬度河口浮游动物群落结构大尺度的变化规律。
2 材料与方法
2.1调查地点、时间和方法
2010—2012年春季分别在北仑河口、九龙江口、瓯江口、长江口和灌河口采集浮游动物样品(表1)。样品采集和处理等均按照《海洋调查规范》进行,用浅水I型浮游生物网(口径50 cm、孔径0.505 mm,网长145 cm),自海底至海面垂直拖拽获得。采集样品均经5%福尔马林溶液固定,在体视显微镜下进行分类鉴定和计数(ind/m3)。本文的浮游动物不包括浮游幼体(虫)、水母类和原生动物。
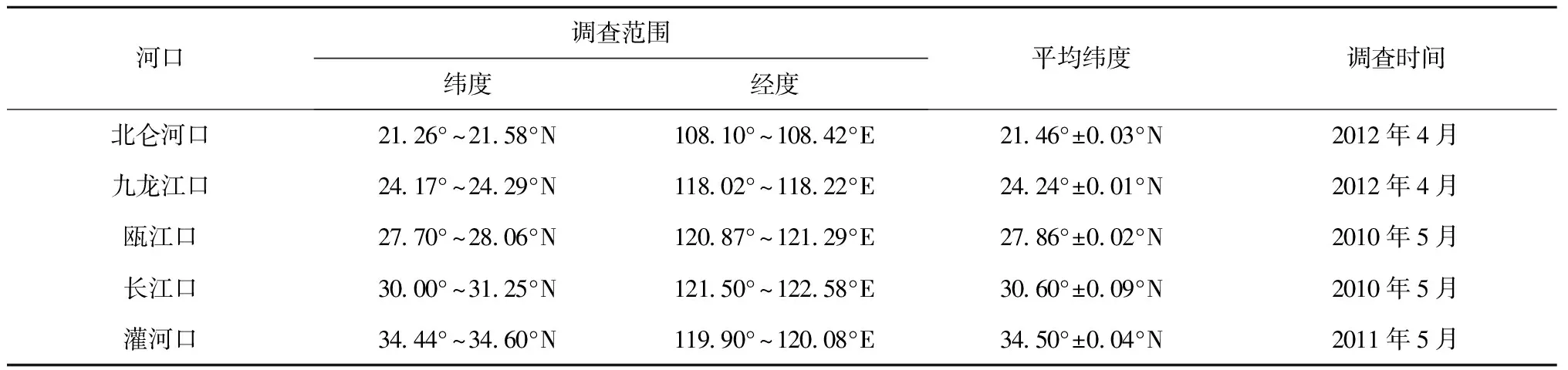
表1 调查站位情况
2.2数据处理方法
采用Shannon-Weaver多样性指数H′,均匀度指数J′,物种更替率(R)和群落相似性系数(S)对不同纬度河口浮游动物群落进行分析[12—15],计算方法为:
(1)
(2)
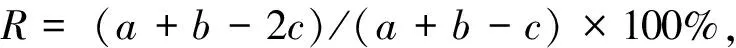
(3)
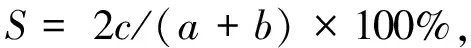
(4)
式中,s为种类数,Pi为第i种的个体数占该群落总个体数的比例,a和b分别为不同河口浮游动物的总种数;c为两个河口站位中共有种类数。
2.30-1型变量聚类分析计算方法
为了分析河口浮游动物群落种类的变化和分布的差异,本文用0-1型变量聚类分析方法来探究我国不同河口浮游动物群落的区系特征。建立“河口”ד浮游动物种类是否出现”的二元生物数据矩阵,其中用“1”表示种类在该河口出现,“0”代表种类在该河口不出现。然后进行系统聚类分析,聚类距离采用Jaccard系数,聚类方法选择类平均法,经分析就可得到不同河口之间浮游动物群落的种类分布相似程度[16]。
3 结果与分析
3.1不同河口浮游动物群落多样性的变化
不同河口浮游动物群落的种类数(s)、多样性指数(H′)和均匀度指数(J′)平均值分别为21±3.18、1.51±0.22和0.53±0.06。从表2可以看出,春季我国各河口浮游动物的多样性不高,均匀度也较低。长江口的种类数最多,九龙江口的多样性指数和均匀度均是最高的;灌河口的种类数最少,北仑河口的多样性指数和均匀度均是最低的。

表2 不同河口浮游动物群落多样性的分布
3.2不同河口之间总种类数和共有种类数的变化
从不同河口之间总种类数和共有种类数的变化(表3)可以看出,两河口间总种类数变化范围为29~54种,共有种类均在3~15种之间变化。其中北仑河口与长江口的总种类数最多,达到54种;九龙江口与灌河口的总种类数最少,仅有29种。瓯江口与长江口之间共有种类数最多,为15种;次之为北仑河口和九龙江口之间,共有种类数为10种,灌河口与北仑河口之间共有种类数最少,为3种。

表3 不同河口之间浮游动物总种类数和共有种类数
注:左下半部分为共有种类数c;右上半部分为总种类数a+b。
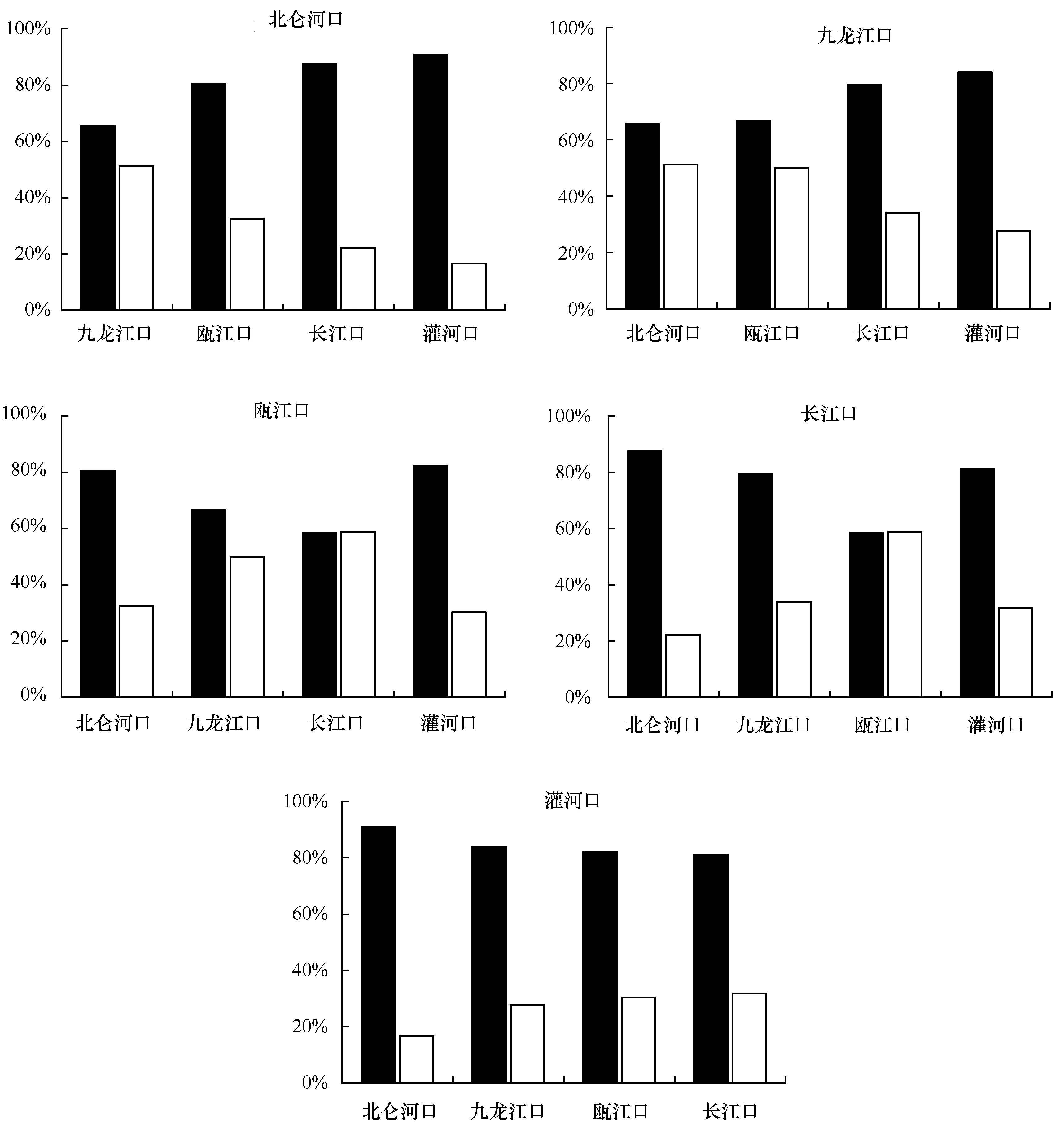
图1 各河口分别与其他4个河口之间的物种更替率(R)和群落相似系数(S)的变化Fig.1 Variations of turnover rate and similarity coefficient of zooplankton between different estuaries ■:物种更替率,□:群落相似系数■: turnover rate, □:similarity coefficient
3.3不同河口之间物种更替率和群落相似系数的变化趋势
两河口之间物种更替率(R)和群落相似系数(S)的平均值分别为77.62%±0.11和35.53%±0.14。北仑河口和九龙江口之间的R值较低,为65.52%,S值较高,为51.28%;瓯江口和长江口之间的R值最低,为58.33%,S值最高,为58.82%;灌河口与其他4个河口之间的R值均较高,S值均较低。
采用物种更替率(R)和群落相似性系数(S)来研究不同纬度河口之间浮游动物群落的变化趋势。通过分析物种更替率和相似系数与两河口所处纬度的差值(两河口调查站位纬度平均值之间的差值即为纬度差,ΔN)的关系发现(表4),在我国近海,两河口之间的纬度差与物种更替率呈正相关,线性回归方程为:R=0.026ΔN+0.608;与群落相似系数呈负相关,线性回归方程为:S=-0.034ΔN+0.578。也就是说两河口之间纬度差越大,则物种更替率(R)越大,群落相似性系数(S)越小,河口浮游动物群落越不相似;相反,两河口之间纬度差越小,则物种更替率(R)越小,群落相似性系数(S)越大,河口浮游动物群落越相似。
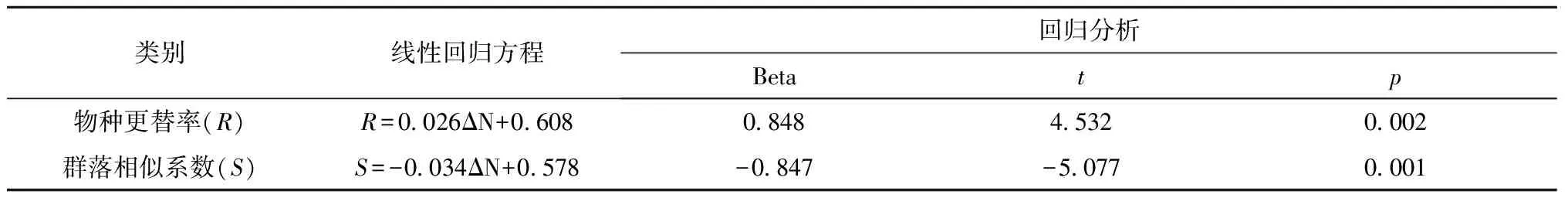
表4 不同河口之间浮游动物更替率和相似系数与纬度差的关系
注:ΔN为两河口调查站位纬度平均值之间的差值,即为纬度差。
3.4不同河口浮游动物区系的划分
根据图2的结果发现,北仑河口和九龙江口在0.73距离水平上聚为一类,表明这两个河口浮游动物种类组成较相似;在0.64距离系数水平上,瓯江口与长江口聚为一类,表明这两个河口浮游动物种类组成非常相似;在0.86距离系数水平上,南部的北仑河口、九龙江口、瓯江口和长江口聚为一大类,表明这4个河口在浮游动物种类组成上有一定的相似性;灌河口独自成为一类,在较大的距离系数上与其他4个河口聚为一类,说明灌河口与其他4个河口浮游动物的种类组成差异较明显。结果表明我国的5个河口自南向北划分成3个浮游动物区系:北仑河口和九龙江口为一个区系,瓯江口与长江口为一个区系,灌河口单独构成一个区系。
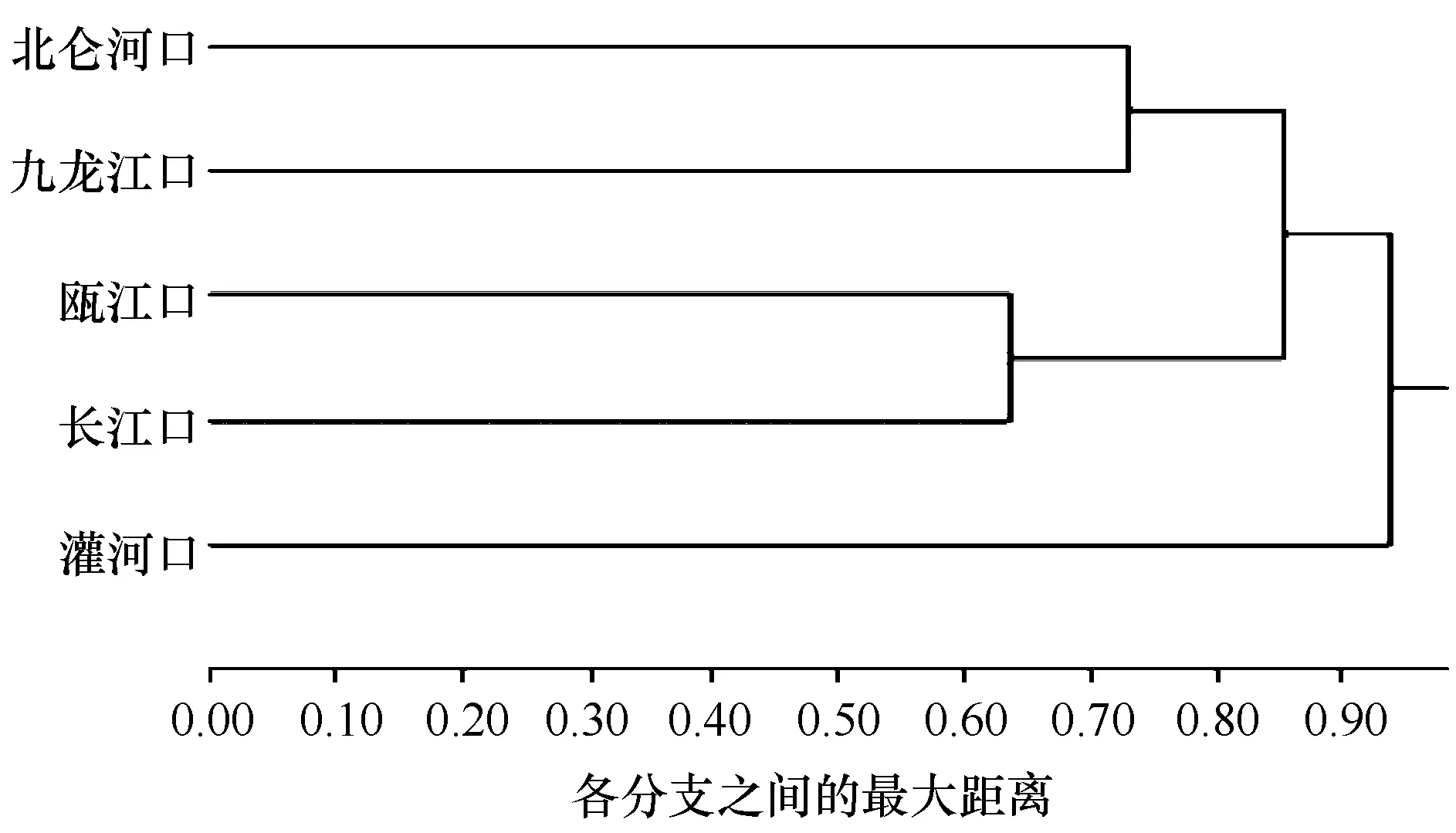
图2 不同河口之间浮游动物的0-1聚类结果Fig.2 0-1 cluster analysis of zooplankton between different estuaries
3.5不同河口浮游动物主要种类的出现频率和丰度百分比
本文取浮游动物在河口出现频率高于20.00%的为该河口主要种类,则自南向北不同河口的主要种类分别为12种、10种、9种、12种和8种。中华哲水蚤(Calanussinicus)是5个河口的共有主要种类,出现频率和丰度百分比均较高;纳嘎带箭虫(Zonosagittanagae)、真刺唇角水蚤(Labidoceraeuchaeta)和中华假磷虾(Pseudeuphausiasinica)等在不同河口的出现频率也较高;软拟海樽(Doliolettagegenbauri)、虫肢歪水蚤(Tortanus(Eutortanus)vermiculus)和强壮滨箭虫(Aidanosagittacrassa)等则是特定河口的主要种类。
3.6不同河口浮游动物主要种类的生态类群
春季我国不同河口浮游动物的主要种类生态类群以亚热带种和暖温带为主(表5)。
北仑河口:主要种类以软拟海樽、肥胖软箭虫、鸟喙尖头溞和锥形宽水蚤等亚热带种为主,暖温带种只有中华哲水蚤、小拟哲水蚤、近缘大眼水蚤和纳嘎带箭虫等4种。
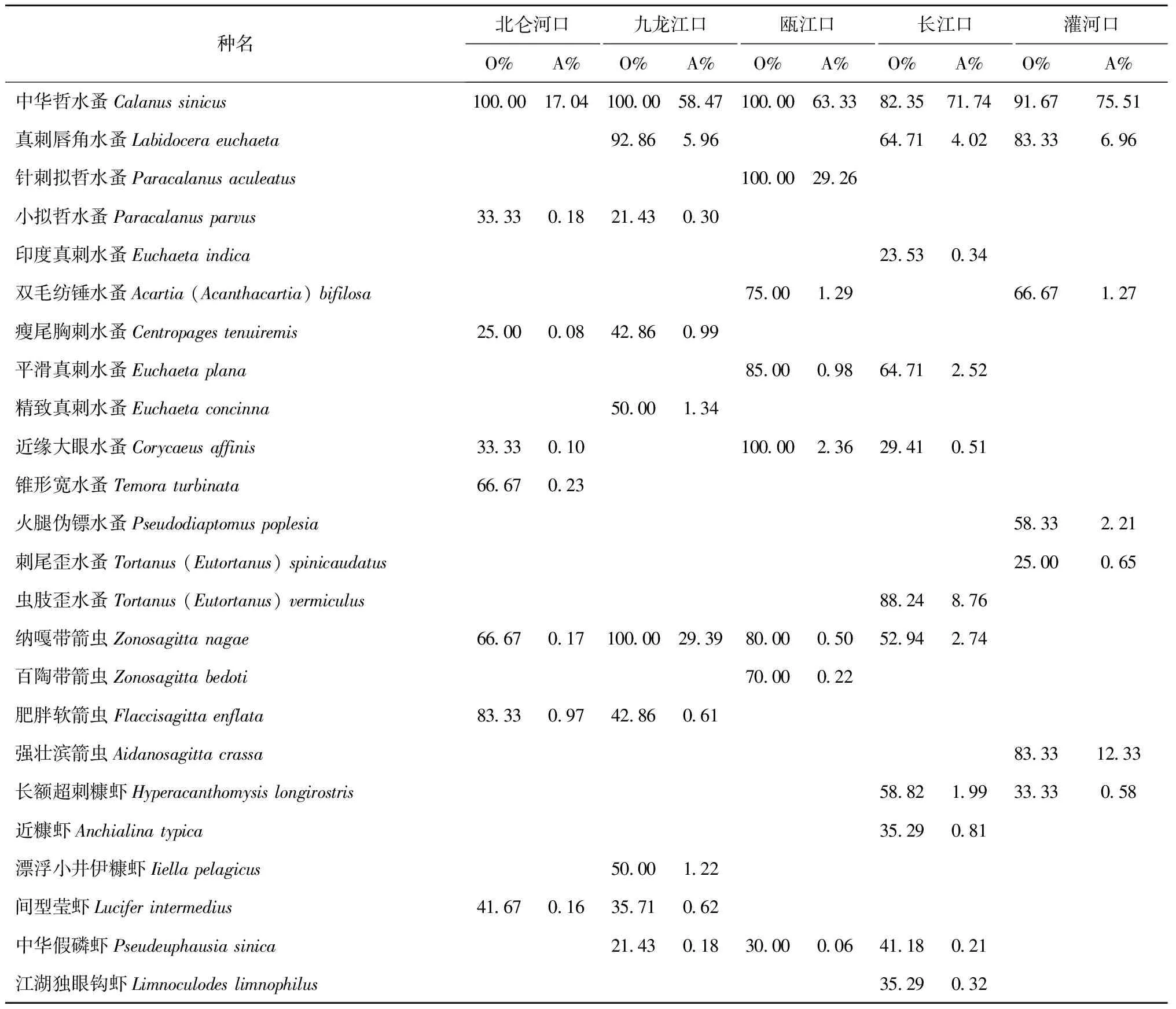
表5 不同河口浮游动物主要种的出现频率和丰度百分比
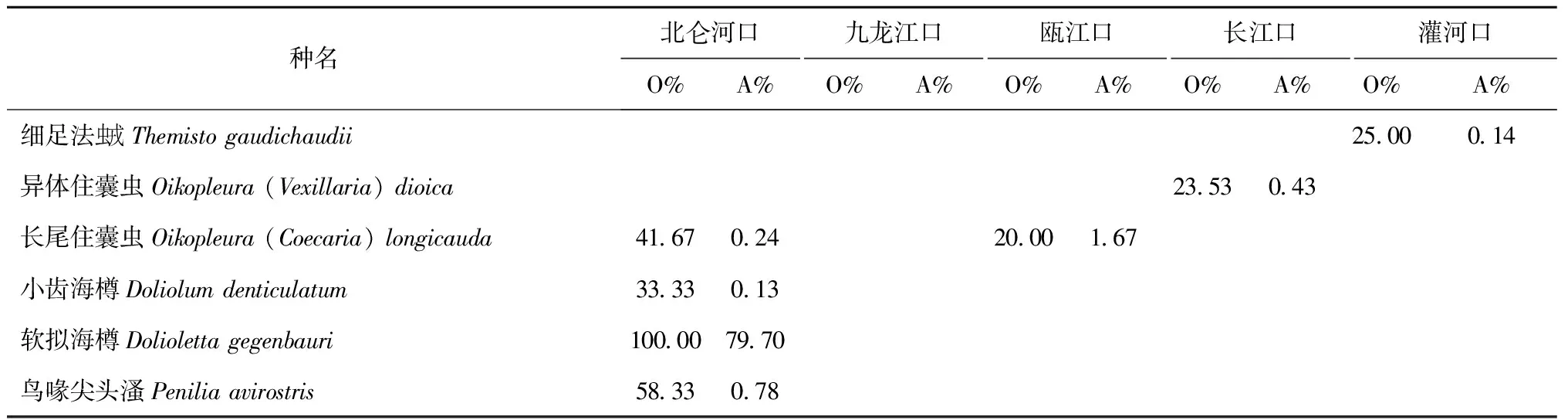
续表5
注:O%为出现频率的百分比,A%为丰度百分比。
九龙江口:主要种类中亚热带种略多,包括精致真刺水蚤、瘦尾胸刺水蚤、肥胖软箭虫和间型莹虾等;暖温带种有中华哲水蚤、纳嘎带箭虫和真刺唇角水蚤等。
瓯江口:亚热带种和暖温带种种类数相近,分别有针刺拟哲水蚤、百陶带箭虫、中华假磷虾和中华哲水蚤、平滑真刺水蚤、双毛纺锤水蚤等。
长江口:亚热带种有虫肢歪水蚤、中华假磷虾和近糠虾等;暖温带种包括中华哲水蚤、真刺唇角水蚤和长额超刺糠虾等;暖温带种略多。
灌河口:主要种类中暖温带种最多,有中华哲水蚤、真刺唇角水蚤、双毛纺锤水蚤和强壮滨箭虫等;亚热带种仅有火腿伪镖水蚤一种。
4 讨论
4.1水温对不同纬度河口浮游动物群落的影响
春季,我国近海因南北纬度跨度大,使得自南向北海水温度变化明显。由此浮游动物群落结构形成相应的变化,水温成为影响近海浮游动物群落的重要环境因子[17—18]。加上不同浮游动物对温度的耐受能力不同,从而导致近海河口海域浮游动物的分布和丰度发生差异。
因受太阳辐射和海流的影响,春季我国近海等温线大致呈纬向分布,近海沿岸海域水温变化范围为10~26℃[4]。也就是说我国近海海域水温随着纬度的升高而自南向北逐渐降低。近海独特的水温变化趋势,会对不同纬度河口浮游动物的群落产生显著影响。丁峰元等[19]指出春季东海区近海,水温随着纬度增加而逐渐降低,群落间差异极显著的水温会影响浮游动物的分布。正是因为不同河口海域水温存在差异,从而致使软拟海樽、精致真刺水蚤和强壮滨箭虫等仅成为特定河口的主要种类。中华哲水蚤随着水温的不断降低,其丰度百分比不断升高,对各河口浮游动物的总丰度贡献逐渐增大。偏高温的亚热带种在南部的北仑河口和九龙江口丰度百分比增大,种类较多;特别是北仑河口,亚热带种软拟海樽的丰度百分比超过中华哲水蚤,成为最主要的种类。偏低温的暖温带种,则在北部的河口大量出现,亚热带种则出现较少。
这是因为温度会直接影响浮游动物的新陈代谢、生长和繁殖发育等方面,从而对浮游动物的分布和丰度产生影响[20—21]。软拟海樽是狭温的亚热带种,北仑河口适宜的温度使其出现率和丰度百分比均是最高的;在其他河口可能因水温不适宜而未出现。徐兆礼等[22]在研究东海海樽类分布时发现:春季软拟海樽高丰度区在温度较高的暖流影响海域,而在其他海域丰度较低。暖温带种强壮滨箭虫是渤海和黄海的主要种类,较喜低温;其最适温度为10~15℃[23]。春季,灌河口海域的水温较低,适宜其生长,所以其大量出现。而在南部近海沿岸海域,水温多在20℃左右,较高的温度限制了强壮滨箭虫的出现。水温成为强壮滨箭虫向南扩布的限制因子。中华哲水蚤作为暖温带广布种,适温范围是1~27℃[24];较宽的适温使其自南海北部的北仑河口到南黄海的灌河口均有分布,且出现率均高于80.00%。但由于中华哲水蚤繁殖最盛期的温度为10~18℃,上限大约是23℃[25—26];这就导致其在不同河口海域的丰度存在巨大差异。春季,北仑河口海域水温接近中华哲水蚤的繁殖上限,抑制了其生长和繁殖,故丰度百分比仅为17.04%。而在灌河口,海水温度适宜中华哲水蚤的生长和产卵,故丰度百分比高达75.51%。
自南向北,受我国近海水温变化的影响,河口海域暖温带种逐渐增多,亚热带种逐渐减少,中华哲水蚤的丰度百分比逐渐升高。不同河口海域浮游动物分布的差异对我国海洋气候和环境变化产生了明显的响应。
4.2水团对不同纬度河口浮游动物群落的影响
水团对河口浮游动物群落的影响主要在两个方面,一是水团的携带作用,影响浮游动物的种类组成;二是改变了河口海域的水温,从而影响了浮游动物的生态类型。
在冬春季,中国近岸的沿岸流几乎连城一片,使得沿岸流水团可以裹挟着东海沿岸的中华哲水蚤逐步带入台湾海峡南部及以南海区[27]。导致春季九龙江口和北仑河口海域有中华哲水蚤出现,并且出现率高达100.00%。在北仑河口,沿岸流携带着中华哲水蚤进入河口海域的同时,可能也一并带入了小拟哲水蚤、近缘大眼水蚤和纳嘎带箭虫等暖温带种。郑白雯等[28]就认为北部湾北部春季出现的暖温带种类应是由沿粤西沿岸流带入。
春季,东南沿海东北季风减弱,但长江冲淡水仍流向南方,与浙江沿岸流混合,形成混合水团,影响着浙江近岸河口海域浮游动物的种类[29—30]。因此,瓯江口海域受北下低温沿岸水团的影响,暖温带种较多。但此时台湾暖流逐步增强,带来的外海高温水团,使得该河口海域的水温升高。受此影响,偏高温的亚热带种百陶带箭虫和针刺拟哲水蚤成为该河口特有的主要种类。
长江口在春季主要受冲淡水和外海水团的影响[31]。长江巨大的冲淡水在河口海域形成了广阔的河口羽状锋,致使河口低盐种虫肢歪水蚤大量出现,成为长江口的主要种类。外海水团的侵入,使得长江口虽然纬度偏北,但主要种类种中仍出现了较多亚热带种。不同水团携带着不同适应性物种在长江口海域汇集,致使长江口浮游动物种类数高达31种,主要种类中暖温带种和亚热带种的数量都远高于邻近河口。纪焕红和叶属峰[32]研究表明长江口海域因不同水团交汇,导致浮游动物种类的组成和生态类型混杂,群落结构呈现多种结构复合的特征。
4.3不同纬度河口浮游动物群落相似性大尺度变化趋势
在春季,我国不同河口之间所处纬度相差越大,浮游动物群落越不相似;反之,则越相似。反映在物种更替率(R)和群落相似性系数(S)上就是两河口之间纬度差越大,R值越大,S值越小;相反,则R值越小,S值越大。例如,长江口和瓯江口之间纬度差最小,所以两河口之间R值最低,为58.33%,S值最高,为58.82%,浮游动物群落非常相似。灌河口位于最北边,与南部河口之间的纬度差非常大,故与其他河口之间R值均在80.00%以上,S值均维持在32.00%以下;故与其他河口浮游动物群落相似性差异较大。
不同河口之间浮游动物群落相似性的这种变化趋势,与各河口间的水温变化密切相关。在我国近海,水温随着所处纬度的升高不断降低;同样,河口海域也有相同的情况。春季,这种独特的河口海域水温变化趋势,影响了不同适温浮游动物在该河口的分布。邻近河口之间,因水温相近,相似适温的浮游动物较多,从而导致两河口间浮游动物群落较相似。北仑河口自南向北,与其余河口之间的浮游动物共有种分别为10、7、6和3种;与九龙江口因同处南部海域,水温相近,浮游动物群落就表现出极高的相似性,R值为65.52%,S值高达51.28%。灌河口由于具有明显的暖温带特征,海域水温较低,故与其他河口浮游动物群落相似性较小。特别是与南部亚热带气候特征明显的北仑河口,因两河口之间温度相差较大,因而两河口之间的R值最高(90.91%),S值最低(16.67%)。
4.4不同纬度河口浮游动物群落多样性大尺度变化趋势
春季,我国不同纬度河口浮游动物群落多样性差异明显;自南向北,不同河口浮游动物的多样性呈现降低的趋势。例如,九龙江口多样性指数为2.18,瓯江口为1.31,灌河口仅1.19。在5个河口中,出现了长江口(1.86)和北仑河口(1.01)两个例外情况,前者多样性高于邻近河口浮游动物的多样性,北仑河口浮游动物多样性最低。之所以发生这一情况,是与不同河口海域多样性形成的机制有关。
一般来说,种类越丰富、种类数量分布越均匀,多样性指数就越高[33]。这就是说多样性指数决定因素除了种类数,还有总丰度在不同种类之间分配的均匀性。春季,中华哲水蚤是各河口最主要的出现种类。中华哲水蚤作为暖温带种,从南到北在不同纬度河口的丰度百分比逐渐升高,对浮游动物群落结构的影响越来越大,严重影响了不同种类之间丰度的均匀性。中华哲水蚤的这种影响也存在于椒江口和舟山渔场海域[30,34],并且使春季浮游动物的多样性指数远低于其他季节。在南部河口,中华哲水蚤的丰度百分比较低,使得浮游动物群落的均匀性较高。同时,南部河口亚热带气候特征逐渐变得明显,导致亚热带种的种类数开始增多,河口浮游动物的种类数高于北部河口。以上两个因素的共同作用,导致我国从南到北不同河口浮游动物多样性指数有逐渐升高的趋势。
但我国不同纬度河口径流差异较大,气候变化明显,河口水域生态环境复杂;使得部分河口浮游动物群落的多样性出现波动。其中长江口作为我国最大的河口,入海径流高达9.2亿立方米[4],巨大的径流使河口水域盐度梯度变化面积较大,河口羽状锋的分布范围广阔。盐度成为影响长江口浮游动物群落结构的主要因素[35—36]。有研究表明[31,37]在长江口咸淡水过渡带,中华华哲水蚤(Sinocalanussinensis)、虫肢歪水蚤和真刺唇角水蚤等低盐种是主要种类,而中华哲水蚤在口外海域才是主要种类。不同盐度适应性的浮游动物大量出现,使得种类数大量增加,均匀度也变高,进而多样性指数提高。杨宇峰等[18]指出盐度是决定河口浮游动物群落结构变化的关键性非生物因子,影响浮游动物的种类和丰度的分布。对于北仑河口,中华哲水蚤已经不是最主要的出现种类,取而代之的是典型的亚热带种软拟海樽。适应于偏高温高盐环境的海樽类大量出现,致使河口浮游动物群落结构单一,均匀度非常低,使得多样性指数最低。最主要种类浮游动物生态类群的转换,使得北仑河口浮游动物群落多样性发生了巨大变化。
由此可见,在我国沿海河口海域,当暖温种的丰度百分比较高时,从南到北不同河口浮游动物多样性指数呈逐渐升高的趋势;但到了北仑河口这样典型的亚热带河口,因终年亚热带中均可以大量生长,导致亚热带种类的丰度百分比较高,使得浮游动物多样性指数较低。
4.5不同纬度河口浮游动物的区系划分及其生态特征
基于历年海洋水文气候要素,我国近海海域以“全夏线”和“无冬线”为界,可以划分为热带海域(“全夏线”以南)、亚热带海域(“全夏线”和 “无冬线”之间)和温带海域(“无冬线”以北),其中 “无冬线”以北至长江口外海域是亚热带和温带型海域的过渡区域[38—39]。通过0-1型变量聚类对5个不同纬度河口浮游动物群落进行区系划分,发现划分的3个浮游动物区系与我国近海海洋水文气候区划范围高度吻合。因此,对3个河口浮游动物区系作如下的命名:北仑河口和九龙江口组成的区系命名为南部亚热带区系,位于亚热带海域;瓯江口和长江口构成的区系命名为中部过渡区系,位于亚热带和温带的过渡海域;灌河口独自构成的区系命名为北部温带区系,位于温带海域。
南部亚热带区系位于我国亚热带海域,全年无冬,春季较短,海水温度较高,亚热带种成为主要种类;暖温带种只有中华哲水蚤、小拟哲水蚤和纳嘎带箭虫等4种,丰度百分比也较低。加之南海次表面水和海峡暖水给河口海域带来了许多外海适应高温的亚热带种,使得亚热带种大量出现,导致北仑河口和九龙江口之间群落结构较相似,物种更替率只有65.52%,群落相似系数高达51.28%。
中部过渡区系在我国近海中部,是亚热带和温带的过渡海域;水温较低,温带海洋环境特征仍较强,中华哲水蚤、近缘大眼水蚤和平滑真刺水蚤等暖温带种类丰度百分比较高,例如中华哲水蚤的丰度在两河口的比例分别为63.33%和71.74%。但随着季风减弱,台湾暖流由南往北逐步增强,外海暖水团的加强,使该浮游动物区系也出现了较多亚热带种。此时,暖温带种虽然丰度仍较高,但主要种类种中亚热带种比例也较高,中部区系成为亚热带种和暖温带种浮游动物过渡区系。
北部温带区系在温带海域,温带海洋特征非常明显,浮游动物群落结构与南部河口之间差别较大,与其他河口之间的共有种类非常少,群落相似性较低。主要出现的浮游动物多是暖温带种,亚热带种仅有两种。暖温带种中华哲水蚤的丰度百分比高达75.51%,暖温带喜冷广布种强壮滨箭虫也大量出现。
5 结论
我国近海独特的水温和水团的变化,影响了河口海域浮游动物的分布和丰度。自南向北,软拟海樽、精致真刺水蚤和强壮滨箭虫等成为不同河口的特有主要种类,中华哲水蚤在不同河口的丰度百分比逐渐增加;主要种类中暖温带种类数和丰度逐渐增加,亚热带种类数和丰度逐渐减少,形成了亚热带种和暖温带种动态变化的河口浮游动物群落结构。
自南向北,我国不同纬度河口浮游动物的多样性呈现降低的趋势;不同河口之间纬度相差越大,浮游动物群落越不相似;反之,浮游动物群落越相似。不同河口之间浮游动物的物种更替率与纬度差(两河口所处纬度的差值,ΔN)呈正相关关系(R=0.026ΔN+0.608,p=0.002);群落相似系数与纬度差呈负相关关系(S=0.034ΔN+0.578,p=0.001)。
春季,我国近海5个河口可以划分为3个不同的浮游动物区系:南部亚热带区系,由北仑河口和九龙江口构成,以亚热带种为主,位于亚热带海域;中部过渡区系,由瓯江口和长江口组成,是亚热带种和暖温带种共同作用的过渡区系,在亚热带和温带的过渡海域, 亚热带种和暖温带种是主要出现的种类;灌河口单独构成北部温带区系,位于温带海域,以暖温带种为主。
致谢:感谢参与海上样品采集和样品鉴定的课题组成员,特别感谢沈晓民老师在论文写作过程中提供的帮助。
[1]廖克. 中华人民共和国国家自然地图集[M]. 北京: 中国地图出版社, 1999.
Liao Ke. National Natural Atlas of China[M]. Beijing: SinoMaps Press, 1999.
[2]李云, 徐兆礼, 高倩. 长江口强壮箭虫和肥胖箭虫的丰度变化对环境变暖的响应[J]. 生态学报, 2009, 29(9): 4773-4780.
Li Yun, Xu Zhaoli, Gao Qian. Effects of global warming onSagittacrassaandSagittaenflata(Chaetognatha) in the Changjiang Estuary during different years[J]. Acta Ecological Sinica, 2009, 29(9): 4773-4780.
[3]Hays G C, Richardson A J, Robinson C. Climate change and marine plankton[J]. Trends in Ecology & Evolution, 2005, 20(6): 337-344.
[4]苏纪兰. 中国近海水文[M]. 北京: 海洋出版社, 2005: 4-164.
Su Jilan. Hydrographic Properties of the Coastal Seas of China[M]. Beijing: China Ocean Press, 2005: 4-164.
[5]巩俊霞, 杨秀兰, 段登选, 等. 黄河入海口水域春季浮游动物群落特征研究[J]. 广东海洋大学学报, 2010, 30(6): 1-6.
Gong Junxia, Yang Xiulan, Duan Dengxuan, et al. A study on zooplankton community characteristics of Yellow River Estuary waters in spring[J]. Journal of Guangdong Ocean University, 2010, 30(6): 1-6.
[6]朱延忠, 刘录三, 郑丙辉, 等. 春季长江口及毗邻海域浮游动物空间分布及与环境因子的关系[J]. 海洋科学, 2011, 35(1): 59-65.
Zhu Yanzhong, Liu Lusan, Zheng Binghui, et al. Relationship between spatial distribution of zooplankton and environmental factors in the Changjiang Estuary and its adjacent waters in spring[J]. Marine Sciences, 2011, 35(1): 59-65.
[7]孙鲁峰, 孙岳, 徐兆礼. 椒江口海域浮游动物群落季节变化特征[J]. 上海海洋大学学报, 2014, 23(1): 131-138.
Sun Lufeng, Sun Yue, Xu Zhaoli. The study on the seasonal variations of zooplankton community in Jiaojiang coastal waters[J]. Journal of Shanghai Ocean University, 2014, 23(1): 131-138.
[8]陈剑, 张宇, 徐兆礼, 等. 夏季闽江口和椒江口浮游动物群落结构的比较[J]. 海洋学报, 2015, 37(2): 111-119.
Chen Jian, Zhang Yu, Xu Zhaoli, et al. Comparison of the community structure of zooplankton between Minjiang River Estuary and Jiaojiang River Estuary during summer[J]. Haiyang Xuebao, 2015, 37(2): 111-119.
[9]张达娟, 闫启仑, 王真良. 典型河口浮游动物种类数及生物量变化趋势的研究[J]. 海洋与湖沼, 2008, 39(5): 536-540.
Zhang Dajuan, Yan Qilun, Wang Zhenliang. Variation in species number and biomass of zooplankton in typical estuaries of China[J]. Oceanologia et Limnologia Sinica, 2008, 39(5): 536-540.
[10]Kibirige I, Perissinott R, Thwala X. A comparative study of zooplankton dynamics in two subtropical temporarily open/closed estuaries, South Africa[J]. Marine Biology, 2006, 148(6): 1307-1324.
[11]Sautour B, Castel J. Comparative spring distribution of zooplankton in three macro tidal European estuaries[J]. Hydrobiologia, 1995, 311(1/3): 139-151.
[12]Shannon C E. The Mathematical Theory of Communication[M]. Urbana: University of Illinois Press, 1949: 3-55.
[13]Pielou E C. The measurement of diversity in different types of biological collections[J]. Journal of Theoretical Biology, 1966, 13: 131-144.
[14]赵志模, 周新远. 生态学引论[M]. 重庆: 科学技术文献出版社重庆分社, 1984: 108-119.
Zhao Zhimo, Zhou Xinyuan. Introduction to Ecology[M]. Chongqing: Science and Technology Literature Publishing House, Chongqing Branch, 1984: 108-119.
[15]沈国英, 黄凌风, 郭丰, 等. 海洋生态学[M]. 3版. 北京: 科学出版社, 2010: 212-219.
Shen Guoying, Huang Lingfeng, Guo Feng, et al. Marine Ecology[M]. 3rd ed. Beijing: Science Press, 2010: 212-219.
[16]唐启义. 实用统计分析及其DPS数据处理系统[M]. 北京: 科学出版社, 2002: 734-737.
Tang Qiyi. DPS Data Processing System for Practical Statistics[M]. Beijing: Science Press, 2002: 734-737.
[17]杜明敏, 刘镇盛, 王春生, 等. 中国近海浮游动物群落结构及季节变化[J]. 生态学报, 2013, 33(17): 5407-5418.
Du Mingmin, Liu Zhensheng, Wang Chunsheng, et al. The seasonal variation and community structure of zooplankton in China Sea[J]. Acta Ecologica Sinica, 2013, 33(17): 5407-5418.
[18]杨宇峰, 王庆, 陈菊芳, 等. 河口浮游动物生态学研究进展[J]. 生态学报, 2006, 26(2): 576-585.
Yang Yufeng, Wang Qing, Chen Jufang, et al. Research advance in estuarine zooplankton ecology[J]. Acta Ecologica Sinica, 2006, 26(2): 576-585.
[19]丁峰元, 李圣法, 董婧, 等. 春季东海区近海浮游动物群落结构及其影响因子[J]. 海洋渔业, 2005, 27(1): 26-32.
Ding Fengyuan, Li Shengfa, Dong Jing, et al. Preliminary study on the zooplankton community structure and its influential factors in the offshore waters of the East China Sea in spring[J]. Marine Fisheries, 2005, 27(1): 26-32.
[20]Devreker D, Souissi S, Seuront L. Effects of chlorophyll concentration and temperature variation on the reproduction and survival ofTemoralongicornis(Copepoda, Calanoida) in the Eastern English Channel[J]. Journal of Experimental Marine Biology and Ecology, 2005, 318(2): 145-162.
[21]Lenz P H, Hower A E, Hartline D K. Temperature compensation in the escape response of a marine copepod,Calanusfinmarchicus(Crustacea)[J]. Biological Bulletin, 2005, 209(1): 75-85.
[22]徐兆礼, 林茂, 张金标. 东海海樽类优势种的数量变化[J]. 动物学报, 2006, 52(1): 53-62.
Xu Zhaoli, Lin Mao, Zhang Jinbiao. Changes in dominant species of Thaliacea in the East China Sea[J]. Acta Zoologica Sinica, 2006, 52(1): 53-62.
[23]刘青, 曲晗, 张硕. 强壮箭虫对温度、盐度的耐受性研究[J]. 海洋湖沼通报, 2007(1): 111-116.
Liu Qing, Qu Han, Zhang Shuo. Preliminary studies on the tolerance to temperature and salinity inSagittacrassa[J]. Transactions of Oceanology and Limnology, 2007(1): 111-116.
[24]Wang Rong, Zuo Tao, Wang Ke. The Yellow Sea cold bottom water-an oversummering site forCalanussinicus(Copepoda, Crustacea)[J]. Journal of Plankton Research, 2003, 25(2): 169-183.
[25]Huang C, Uye S, Onbé T. Geographic distribution, seasonal life cycle, biomass and production of a planktonic copepodCalanussinicusin the Inland Sea of Japan and its neighboring Pacific Ocean[J]. Journal of Plankton Research, 1993, 15(11): 1229-1246.
[26]陈清潮. 中华哲水蚤的繁殖、性比率和个体大小的研究[J]. 海洋与湖沼, 1964, 6(3): 272-288.
Chen Qingchao. A study of the breeding periods, variation in sex ratio and in size ofCalanussinicusBrodsky[J]. Oceanologia et Limnologia Sinica, 1964, 6(3): 272-288.
[27]尹健强, 黄良民, 李开枝, 等. 南海西北部陆架区沿岸流和上升流对中华哲水蚤分布的影响[J]. 海洋学报, 2013, 35(2): 143-153.
Yin Jianqiang, Huang Liangmin, Li Kaizhi, et al. Effects of coastal current and upwelling on the distributions ofCalanussinicuson the northwest continental shelf of the South China Sea[J]. Haiyang Xuebao, 2013, 35(2): 143-153.
[28]郑白雯, 曹文清, 林元烧, 等. 北部湾北部生态系统结构与功能研究 Ⅰ. 浮游动物种类组成及其时空变化[J]. 海洋学报, 2013, 35(6): 154-161.
Zheng Baiwen, Cao Wenqing, Lin Yuanshao, et al. Ecosystem structure and function in northern Beibu Gulf Ⅰ. The temporal and spatial variation of species composition on zooplankton[J]. Haiyang Xuebao, 2013, 35(6), 154-161.
[29]董开兴, 徐兆礼. 飞云江口附近海域浮游动物群落特征[J]. 生态学杂志, 2015, 34(3): 744-752.
Dong Kaixing, Xu Zhaoli. Characteristics of zooplankton communities off the Feiyun River estuary[J]. Chinese Journal of Ecology, 2015, 34(3): 744-752.
[30]孙鲁峰, 柯昶, 徐兆礼, 等. 上升流和水团对浙江中部近海浮游动物生态类群分布的影响[J]. 生态学报, 2013, 33(6): 1811-1821.
Sun Lufeng, Ke Chang, Xu Zhaoli, et al. The influence of upwelling and water mass on the ecological group distribution of zooplankton in Zhejiang coastal waters[J]. Acta Ecologica Sinica, 2013, 33(6): 1811-1821.
[31]殷晓龙, 徐兆礼. 长江口不同水域浮游动物群落结构比较[J]. 应用与环境生物学报, 2015, 21(1): 88-95.
Yin Xiaolong, Xu Zhaoli. Comparison of zooplankton community of different areas in Changjiang River Estuary[J]. Chinese Journal of Applied and Environmental Biology, 2015, 21(1): 88-95.
[32]纪焕红, 叶属峰. 长江口浮游动物生态分布特征及其与环境的关系[J]. 海洋科学, 2006, 30(6): 23-30.
Ji Huanhong, Ye Shufeng. Ecological distribution characteristics of zooplankton and its relationship with environmental factors in the Changjiang River Estuary[J]. Marine Sciences, 2006, 30(6): 23-30.
[33]孙军, 刘东艳. 多样性指数在海洋浮游植物研究中的应用[J]. 海洋学报, 2004, 26(1): 62-75.
Sun Jun, Liu Dongyan. The application of diversity indices in marine phytoplankton studies[J]. Haiyang Xuebao, 2004, 26(1): 62-75.
[34]俞存根, 陈小庆, 胡颢琰, 等. 舟山渔场及邻近海域浮游动物种类组成及群落结构特征[J]. 水生生物学报, 2011, 35(1): 183-193.
Yu Cungen, Chen Xiaoqing, Hu Haoyan, et al. Species composition and community structure characteristics of zooplankton in the Zhoushan fishing ground and its adjacent area[J]. Acta Hydrobiologica Sinica, 2011, 35(1): 183-193.
[35]徐兆礼. 长江口邻近水域浮游动物群落特征及变动趋势[J]. 生态学杂志, 2005, 24(7): 780-784.
Xu Zhaoli. Character of zooplankton community and its variation in the water near the Yangtze River estuary[J]. Chinese Journal of Ecology, 2005, 24(7): 780-784.
[36]Marques S C, Azeiteiro U M, Marques J C, et al. Zooplankton and ichthyoplankton communities in a temperate estuary: spatial and temporal patterns[J]. Journal of Plankton Research, 2006, 28(3): 297-312.
[37]汤新武, 蔡德所, 陈求稳, 等. 三峡工程三期蓄水初期长江口水域春季浮游动物群落特征及其与环境的关系[J]. 环境科学学报, 2015, 35(4): 1082-1088.
Tang Xinwu, Cai Desuo, Chen Qiuwen, et al. Spring character of zooplankton community and its relation to environmental factors in Yangtze estuary right after the third-stage impoundment of Three Gorges Reservoir[J]. Acta Scientiae Circumstantiae, 2015, 35(4): 1082-1088.
[38]陈上及, 何维焕, 姚湜予, 等. 中国近海海洋水文气候季节的划分[J]. 海洋学报, 1992, 14(6): 1-11.
Chen Shangji, He Weihuan, Yao Shiyu, et al. Division of Seasons for the ocean hydrology and climate in the coastal waters of China[J]. Haiyang Xuebao, 1992, 14(6): 1-11.
[39]陈上及, 姚湜予. 中国近海海洋水文气候区划: Ⅱ. 聚类分析和模糊聚类软划分[J]. 海洋学报, 1995, 17(3): 1-8.
Chen Shangji, Yao Shiyu. Division of the ocean hydrology and climate by using cluster analysis and ISODATA of fuzzy cluster in the Coastal Waters of China[J]. Haiyang Xuebao, 1995, 17(3): 1-8.
Variation of zooplankton communities in Chinese estuaries along latitudes in spring
Lu Wuyang1,2,Ma Zengling3, Xu Zhaoli1, Gao Qian1
(1. Ministry of Agriculture Key Laboratory of East China Sea and Oceanic Fishery Resources Exploitation, East China Sea Fisheries Research Institute, Chinese Academy of Fishery Sciences, Shanghai 200090, China; 2. College of Marine Sciences, Shanghai Ocean University, Shanghai 201306, China;3.College of Life and Environmental Science, Wenzhou University, Wenzhou 325035, China)
Based on the zooplankton samples from stations in Beilun Estuary, Jiulongjiang Estuary, Oujiang Estuary, Changjiang Estuary (Yangtze River) and Guanhe Estuary(covering 14 latitudes) in April or May of 2010 to 2012, the community features of Diversity index(H′), replacement rate(R) and similarity coefficient(S) were compared. This paper also analyzed the variation and reasons of zooplankton community in different estuaries. The results indicated that the zooplankton community of the five estuaries were divided into three different zooplankton fauna, which were the southern subtropical, central transitional and northern temperate from the south to the north. The southern subtropical zooplankton fauna included Beilun Estuary and Jiulong Estuary. In this fauna, the subtropical species was the mainly and a different diversity index (H′) (1.01 and 2.18) were recorded, located in the subtropical sea. There placement rate(R) and similarity coefficient(S) were respectively 65.52% and 51.28% between Beilun Estuary and Jiulong Estuary. In central transitional zooplankton fauna, constituted by Oujiang Estuary and Changjiang Estuary, it was exclusively of the subtropical species and warm temperate species, which had an alike diversity index (H′) (1.31 and 1.86, respectively), situated in the transition waters between subtropical and temperate. Between the two estuaries, the replacement rate(R) were lowest (58.33%) and the similarity coefficient(S) were highest (58.82%). At high latitude, only the Guanhe Estuary belongs to the northern temperate, the warm temperate species was dominated, which had a lower diversity index (H′) (1.19) and located in the temperate sea. The replacement rate(R) and similarity coefficient(S) of Guanhe Estuary were significantly different from the other estuaries. From south to north, along with the increase of latitude, the structure of estuarine zooplankton community has changed significantly. Linear regression figured that replacement rate(R) was positively correlated with the difference of longitude between the two estuaries in Chinese (ΔN),R=0.026ΔN+0.608(p=0.002). Similarity coefficient(S) was negatively correlated with the difference of longitude (ΔN),S=-0.034ΔN+0.578(p=0.001). In spring, the main reasons which caused the variation of zooplankton community between different estuaries was due to the difference of the influence of temperature and water masses.
estuary; zooplankton; community; latitude; variation
10.3969/j.issn.0253-4193.2016.10.009
2016-03-16;
2016-05-11。
浙江省科技计划项目(2015C33246);海洋公益性行业科研专项经费项目(201305027-8);国家自然科学基金项目(41176131)。
卢伍阳(1992—),男,安徽省合肥市人,主要从事河口生态学研究。E-mail:1097140197@qq.com
徐兆礼,研究员,主要从事海洋浮游动物和海洋生态学研究。E-mail:xiaomin1957@126.com
Q178.53
A
0253-4193(2016)10-0083-11
卢伍阳,马嶒岭,徐兆礼,等. 春季我国不同纬度河口浮游动物群落变化趋势[J].海洋学报,2016,38(10):83—93,
Lu Wuyang,Ma Zengling, Xu Zhaoli,et al. Variation of zooplankton communities in Chinese estuaries along latitudes in spring[J]. Haiyang Xuebao,2016,38(10):83—93, doi:10.3969/j.issn.0253-4193.2016.10.009
