基于形态和叶表皮微形态特征试论国产赖草属属下类群间的系统关系
2016-11-10李兆孟周勇辉刘玉萍
李兆孟 周勇辉 刘玉萍 苏 旭
(青海师范大学生命与地理科学学院,青藏高原药用动植物资源重点实验室,青海省自然地理与环境过程重点实验室,青藏高原环境与资源教育部重点实验室,西宁 810008)
基于形态和叶表皮微形态特征试论国产赖草属属下类群间的系统关系
李兆孟 周勇辉 刘玉萍 苏 旭*
(青海师范大学生命与地理科学学院,青藏高原药用动植物资源重点实验室,青海省自然地理与环境过程重点实验室,青藏高原环境与资源教育部重点实验室,西宁 810008)
通过对先前学者报道的国产赖草属24个物种、72个代表居群形态和叶表皮微形态性状特征的观测和研究,结果发现:(1)原国产赖草属物种在旗叶长宽、叶片被毛状况、花序直曲和长宽、穗轴每节小穗数、每小穗小花数、外稃和内稃长度等15个外部形态学性状上均存在不同程度的差异,可将参试物种区分为3个不同的组;大赖草、粗穗赖草、硕花赖草和柴达木赖草多穗组植物的穗状花序粗壮呈圆锥状、直立、密集,穗轴同节着生3-多枚小穗,每小穗常含3~10小花,颖线状披针形等,致使其处于该属最原始的地位;皮山赖草、若羌赖草和格尔木赖草单穗组物种的穗状花序细弱呈线状、直立、疏松,穗轴每节仅着生1枚小穗,每小穗往往含2~5小花,颖线状披针形或锥形,这些较高级的外部形态特征使得该组植物隶属于赖草属中较高级的类群;而少穗组植物的外部形态性状通常介于前两组植物的中间过渡状态,因而它的系统地位自然也应处于多穗组和单穗组植物之间。同时,依据外部形态性状的递变趋势分析显示,3组植物具有直接的派生渊源。(2)国产赖草属植物的叶表皮皆由长细胞、短细胞、气孔器细胞和刺毛所组成,表现为典型的狐茅型;它们除在长细胞类型和壁的厚薄、气孔器保卫细胞的体积和类型等性状上具有明显重叠而显示相似外,其它多数性状如长细胞的长度和壁的波曲程度、短细胞的分布式样、副卫细胞的形状及刺毛的类型等性状上具有明显差异,其可将赖草属植物鉴分为与形态学界定结果完全相同的3个组。同时,根据3组植物及组内物种叶表皮性状的演化趋势,对各组和组内物种的演化关系和系统位置分析表明,多穗组植物最原始,少穗组植物较进化,单穗组植物最高级;多穗组可能直接派生了较进化的少穗组,并在少穗组的基础上进而产生了最高级的单穗组。赖草属属下类群的这一系统关系与利用外部形态特征所获得的系统与进化关系基本一致。
禾本科;赖草属;形态性状;叶表皮微形态;系统关系
赖草属(LeymusHochst)是禾本科(Poaceae)、小麦族(Triticeae)中一个具有重要经济和生态价值的多年生属[1]。现知,全世界共约有53种,从北极寒温带到南亚热带均有其分布;我国共有33种、4亚种和3变种[2],其中仅有24种、4亚种和2变种被《Flora of China》[3]所记载,主要分布于西北、华北、东北及西南地区,通常生长于海拔500~5 000 m左右的干旱半干旱山坡、草地、林缘、路旁、河滩、墙脚等地,尤其是它们对干旱、寒冷、盐碱等逆境条件具有高度的适应性。同时,赖草属中多数物种为优良牧草,具有较高的饲用价值。另外,绝大多数赖草属植物还具有穗大、粒多、粒大、抗病虫、高光效等优良特性[4],是农牧业上育种的重要基因资源。
赖草属系由Hochstetter[5]于1958年基于欧滨麦(Leymusarenarius(L.) Hochst.)所建立。自该属建立以来,国内外诸多学者对赖草属的范围、形态特征、生境、地理分布和细胞学进行了一系列的研究[6~14],取得了令人称羡的成绩。尽管前人的研究功不可没,但它们却具有一定的局限性,如有的学者仅采用单一的外部形态性状来建立新物种,也有的学者仅采用单一的研究手段来确定物种间的系统发育关系,这种局部的、不完整的研究资料是不能代表整个国产赖草属系统进化关系的;同时,加之赖草属是一个多型性、多变异的属,属内部分近缘物种间性状差异较小,种间界限模糊不清,往往给野外鉴定造成困难或导致室内鉴定的张冠李戴。尤其是先前学者[15~16]依据少量代表种的叶表皮微形态特征确立的国产赖草属的组群划分和系统进化关系究竟是否正确?目前,国内外均未见学者做过任何验证性研究报道。据此,本文以先前学者发表的24个国产赖草属物种作为研究对象,以叶表皮微形态特征为依据,并结合外部形态特征,试图探讨国产赖草属属下类群间的系统进化关系,这不仅可为先前确立的国产赖草属系统进化关系再次进行合理性验证,而且也可为整个小麦族植物系统与进化关系的研究提供一些基础性资料。
1 材料和方法
1.1 实验材料的选择
供试材料全部取自馆藏副份腊叶标本。整个叶表皮解剖和外部形态特征观测材料共涉及168份,尽量选取不同纬度、不同海拔的材料进行实验。由于实验类群叶片上表皮的形状差异较小,实验效果不大,故本研究实验的重点放在叶片下表皮上。同时,根据类群分布范围的大小对各个代表物种的具体份额再次进行分配,即硕花赖草(L.pluriflorus)、大赖草(L.racemosus)、皮山赖草(L.pishanica)、青海赖草(L.qinghaicus)、伊吾赖草(L.yiunensis)、胎生赖草(L.viviparus)、刺状赖草(L.spiniformis)和光洁赖草(L.mundus)分布范围狭窄,选择全部居群的材料进行形态学和叶表皮微形态特征的观察和测量;而羊草(L.chinensis)和赖草(L.secalinus)的分布范围广泛,主要分布于青藏高原及其毗邻地区,故对其仅选择分布于青海、西藏、新疆和甘肃等省区的共16个居群材料进行实验研究。每个代表物种的典型凭证标本详见表1。

表1 实验材料名称及来源
续表1Continuedtable1

种名Species采集地Locality凭证标本∗Voucher实验用途Experimentused天山赖草L.tianschanicus新疆乌鲁木齐Urumqi,Xinjiang新疆队(XinjiangExp.)80MO、LE新疆喀什Kashi,Xinjiang新疆队(XinjiangExp.)1719MO、LE新疆布尔津Buerjin,Xinjiang新疆队(XinjiangExp.)1838MO、LE新疆哈巴河Habahe,Xinjiang新疆队(XinjiangExp.)2244MO、LE胎生赖草L.viviparus新疆叶城Yecheng,Xinjiang黄荣福(R.F.Huang)CG⁃86⁃058MO、LE西藏朗县NangCounty,Xizang西藏科考队(XizangExp.)3464MO、LE新疆赛力木湖Sailimuhu,Xinjiang新疆队(XinjiangExp.)399MO、LE青海赖草L.qinghaicus青海西宁Xining,Qinghai张志和等(Z.H.Zhang,etal.)2957MO、LE宽穂赖草L.ovatus西藏阿里Ali,Xizang西藏科考队(XizangExp.)3759MO、LE西藏晋南Jinnan,Xizang西藏科考队(XizangExp.)3983MO、LE青海玛曲Maqu,Qinghai陈世龙等(S.L.Chen,etal.)1068MO、LE青海西宁Xining,Qinghai植被地理组(VegetationGeographyExp.)997MO、LE新疆叶城Yecheng,Xinjiang吴玉虎(Y.H.Wu)871407MO、LE窄颖赖草L.angustus青海循化Xunhua,Qinghai王为义(W.Y.Wang)5MO、LE新疆若羌Ruoqiang,Xinjiang黄荣福(R.F.Huang)3895MO、LE河北阳原Yangyuan,Hebei黄河队(HuangheExp.)47MO、LE青海玛多Maduo,Qinghai吴玉虎(Y.H.Wu)913MO、LE羊草L.chinensis新疆沙湾Shawan,Xinjiang关可俭(K.J.Guan)778MO、LE青海兴海Xinghai,Qinghai郭本兆(P.C.Kuo)523MO、LE河北保定Baoding,Hebei采集人不详(Anonymous)4016MO、LE山西太原Taiyuan,Shanxi李华锡(H.X.Li)89MO、LE黑龙江嘉荫Jiayin,Heilongjiang王光正(G.Z.Wang)64MO、LE陕西延安Yan’an,Shanxi梁一民(Y.M.Liang)804MO、LE纤细赖草L.tenuis西藏吉隆Jilong,Xizang青藏队(Qinghai⁃XizangExp.)6250MO、LE西藏扎达Zhada,Xizang青藏队(Qinghai⁃XizangExp.)76⁃7856MO、LE西藏多玛Doma,Xizang青藏队(Qinghai⁃XizangExp.)76⁃9068MO、LE西藏朗县NangCounty,Xizang西藏科考队(XizangExp.)3627MO、LE赖草L.secalinus西藏仲巴Zhongba,Xizang青藏队(Qinghai⁃XizangExp.)6504MO、LE甘肃天水Tianshui,Gansu郭本兆(P.C.Kuo)3188MO、LE青海共和GongHe,Qinghai郭本兆,王生新(P.C.Kuo&S.X.Wang)6466MO、LE青海西宁Xining,Qinghai吴玉虎(Y.H.Wu)1923MO、LE细弱赖草L.leptostachyus青海海北Haibei,Qinghai杨福囤和赵瑞林(F.T.Yang&R.L.Zhao)69MO、LE新疆乌鲁木齐Urumqi,Xinjiang李安仁等(A.R.Li,etal.)4128MO、LE青海西宁Xining,Qinghai植被地理组(VegetationGeographyExp.)92MO、LE褐穗赖草L.bruneostachyus山西五寨Wuzhai,Shanxi杨金祥(J.X.Yang.)3678MO、LE河北清河Qinhhe,Hebei采集人不详(Anonymous)329MO、LE青海格尔木Geermu,Qinghai采集人不详(Anonymous)45MO、LE西藏双湖Shuanghu,Xinjiang青藏队(Qinghai⁃XizangExp.)9920MO、LE刺状赖草L.spiniformis新疆玛纳斯Manasi,Xinjiang关可俭(K.J.Guan)938MO、LE光洁赖草L.mundus西藏扎达Zhada,Xizang青藏队(Qinghai⁃XizangExp.)7767MO、LE
注:*凭证标本存放于中国科学院西北高原生物研究所青藏高原生物标本馆(QTPMB)。 MO.用于形态学观察;LE.用于叶表皮观察
Note:*Vouchers are conserved in QTPMB. MO.Used for morphological observation; LE.Used for leaf epidermal observation
1.2 研究方法
1.2.1 外部形态观察
对国产赖草属24个物种、不同来源的72个代表居群的168份馆藏腊叶标本分别进行了旗叶长度、旗叶宽度、叶片被毛状况、穗状花序直曲、花序长度、花序宽度、小穗数、小穗长度、每小穗小花数、颖体形状、颖体长度、颖脉数、第一外稃长度、第一外稃脉数、第一内稃长度等15个外部形态性状的观察和测量。每个代表居群取4~5株,个别居群只选取1~2株观察的平均数,再将每个物种不同代表居群的观察资料平均,以对各个物种进行相互比较;同时,对于分类价值较大的外部形态性状矩阵,应用SPSS软件进行主成分分析(principal component analysis,PCA)。
1.2.2 叶表皮微形态观察
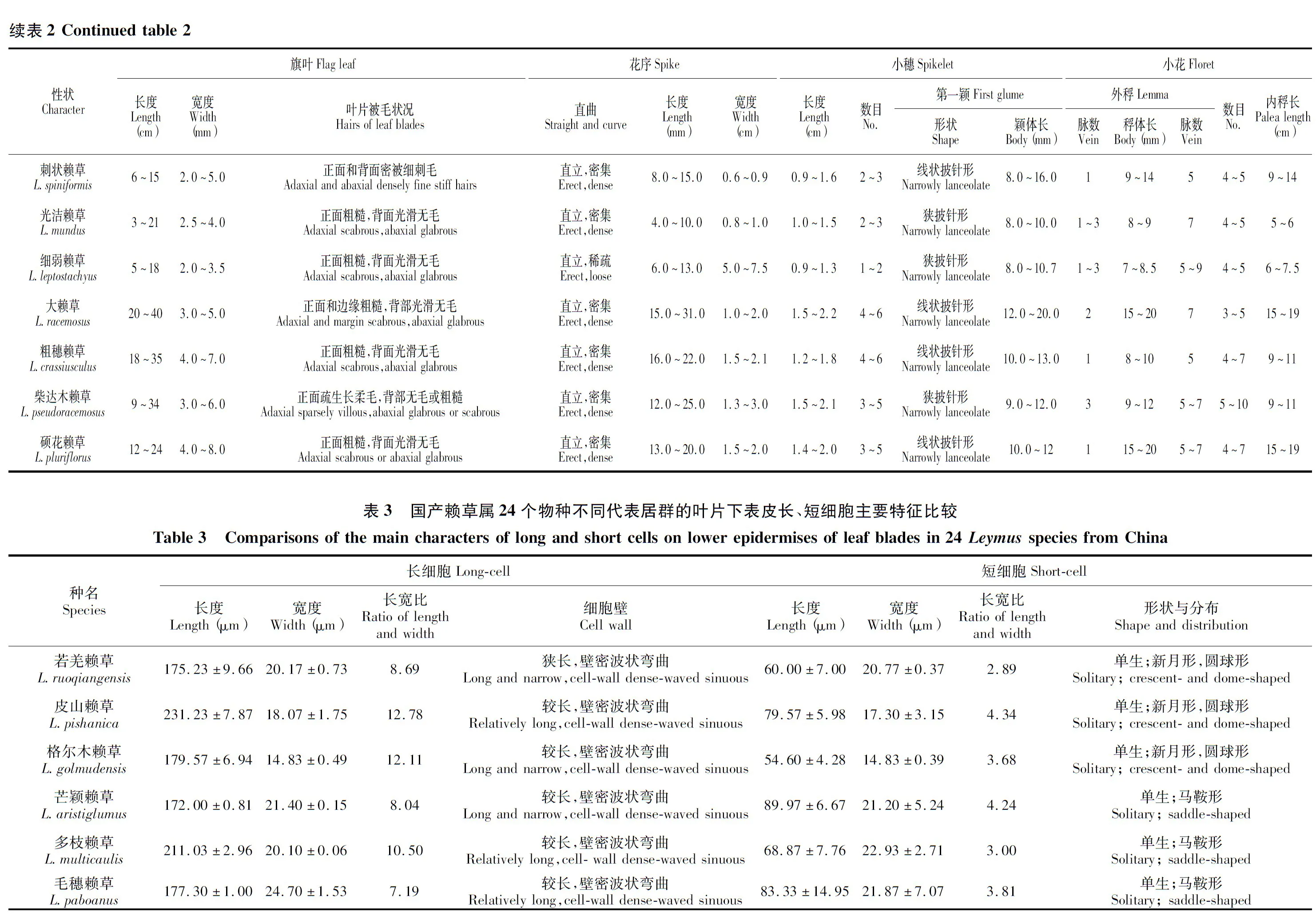
用于叶表皮微形态观察的材料均取自花序下第2叶片的中间部段,共24个物种72个代表居群。按常规叶表皮解剖法操作,即先将叶片用沸水软化15~20 min,材料软化后置于塑料板上用单面刀片轻轻刮削,使其保留叶片上、下表皮薄膜层,然后清水洗净薄膜,用1%的番红染色1 min,再置于光镜下观察,同时进行描述、绘图、拍照等工作,最后以常规永久制片法脱水封固保存。长细胞测其绝对长度,每个代表居群取3个视野,每个视野取10个细胞,取平均值;气孔器大小测其长宽比,每个代表居群取3个视野,每个视野取5个气孔器,取平均值。叶表皮特征用Metcalfe[17]的微形态术语进行描述。
2 结果与分析
2.1 外部形态性状差异分析
国产赖草属24个物种、72个代表居群15个性状的观测、统计结果详见表2。从表2可以明显看出,国产赖草属24个物种的各项统计值基本符合中国一些植物志书上记载的该类植物各对应性状的数值,即使是毛穗赖草、褐穗赖草及格尔木赖草与原始描述相比,也仅在植株高矮、花序长短、两颖片长度等少数性状上有微小差异,这可能与观察标本的数量、区域、生境有关,但并未超出该类群的性状变异范围。显然,这些外部形态性状特征均在前人描述的小麦族范畴内,隶属于小麦族类群无疑。同时,在表2所列的性状数据中,有些性状如旗叶长度和宽度、叶片被毛状况、颖脉数在参试物种间具有一定差异,但在组间则表现重叠,差异皆不明显;而其它绝大多数性状在国产赖草属的组间存在明显差异,至少可将先前界定的3个组群的外部形态间隔大小显示出来,表明国产赖草属划分为3个不同组群的合理性和正确性。此外,我们还对花序长度、花序直曲、小穗数、每小穗小花数、第一颖颖体长度、第一外稃长度等15个具有较大分类价值形态性状特征构成的矩阵进行了主成分分析。依据主成分做二维散点图,结果显示国产赖草属所有参试居群呈现3个明显不同的分布区,它们可以将不同物种的代表居群明显区分开来,与先前界定的3个组群完全相符,再次表明国产赖草属隶属于3个独立的组群(图1)。

图1 主要外部形态性状主成分的散点图 若羌赖草;皮山赖草;格尔木赖草;芒颖赖草;多枝赖草;毛穗赖草;伊吾赖草;弯曲赖草;天山赖草;胎生赖草;青海赖草;宽穂赖草;窄颖赖草;羊草;纤细赖草;赖草;褐穗赖草;刺状赖草;光洁赖草;细弱赖草;大赖草;粗穗赖草;柴达木赖草;硕花赖草Fig.1 2D PCA scatter plot of principal component in main external morphological characters L.ruoqiangensis; L.pishanica; L.golmudensis; L.aristiglumus; L.multicaulis; L.paboanus; L.yiunensis; L.flexus; L.tianschanicus; L.viviparus; L.qinghaicus; L.ovatus; L.angustus; L.chinensis; L.tenuis; L.secalinus; L.bruneostachyus; L.spiniformis; L.mundus; L.leptostachyus; L.racemosus; L.crassiusculus; L.pseudoracemosus; L.pluriflorus
故下面将3组植物的外部形态特征概总如下:
多穗组(Sect.Racemosus):穗状花序常直立,密集;小穗通常3-多枚生于穗轴每节,每小穗含3~12小花;第一颖颖体狭长,介于9~20 mm,呈披针形或狭披针形;外稃先端渐尖,具5~7脉;植株一般高80~110 cm。
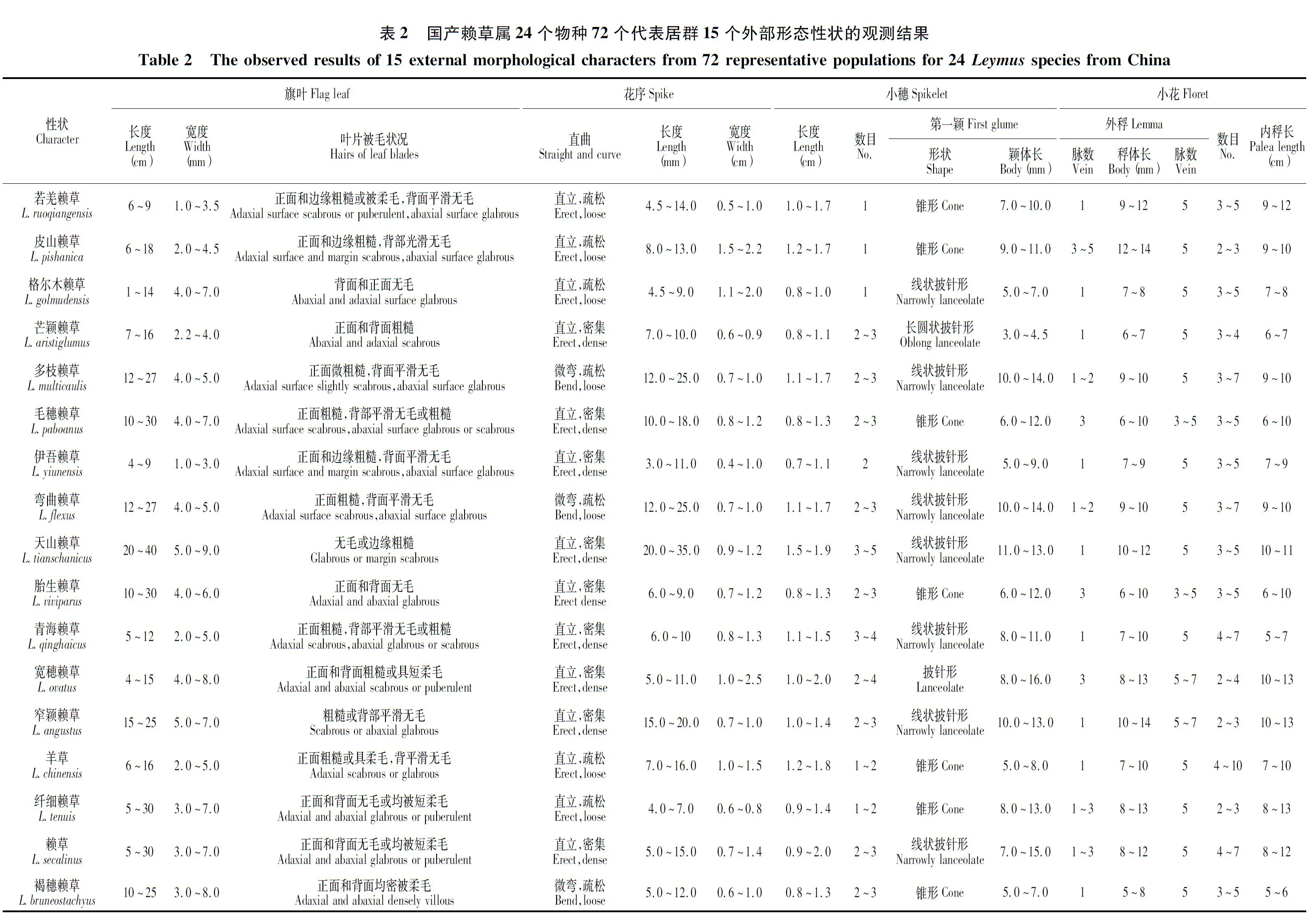



少穗组(Sect.Leymus):穗状花序直立,稀弯曲或下垂;小穗常2~3枚生于穗轴每节,每小穗含3~7小花;第一颖颖体较长,长度介于3~16 mm,呈狭披针形或锥刺形;外稃无芒或具2~7 mm长的短芒,常具5脉;植株一般高20~100 cm。
单穗组(Sect.Anisopyrum):穗状花序直立,疏松;小穗通常单生(偶双生)于穗轴每节,每小穗含3~5小花;第一颖颖体稍长,介于5~11 mm,呈线状披针形或锥刺形;外稃无芒或具短尖头,常具5脉;植株一般高10~70 cm。
2.2 叶表皮微形态性状差异分析
光镜下观察了国产赖草属24个物种、72个代表居群的叶表皮微观特征,发现它们的叶表皮皆由长细胞、气孔器细胞、短细胞和刺毛4类结构细胞所组成(图版Ⅰ~Ⅳ)。长细胞普遍分布于各个参试物种中,呈长方形纵向相接成行,大致水平排列于脉间和脉上,通常脉上着生1~5列,脉间着生4~27列,长度为75~600 μm,宽度为16~45 μm,细胞壁常厚且呈波状弯曲;短细胞孪生或单生,稀不着生,与长细胞相间排列,呈马鞍形、狭马鞍形、新月形、圆球形;气孔器细胞为椭圆形,常带状分布于表皮区域或1~2列生于脉间,长45~238 μm,宽28~70 μm,保卫细胞哑铃形,副卫细胞呈高圆顶形、低圆顶形或平边形;刺毛主要着生于脉上,少数生于脉间,且脉上多刺,脉间多钩。

图版Ⅰ 国产赖草属24个物种的叶片下表皮光学显微照片 1.大赖草;2.柴达木赖草;3.粗穗赖草;4.硕花赖草;5.赖草;6.疏节赖草PlateⅠ Light microscopic micrographs on the lower epidermises of leaf blades of 24 Leymus species from China 1.L.racemosus; 2.L.pseudoracemosus; 3.L.crassiusculus; 4.L.pluriflorus; 5.L.secalinus; 6.L.laxinodis

图版Ⅱ 国产赖草属24个物种的叶片下表皮光学显微照片 1.毛穗赖草;2.窄颖赖草;3.天山赖草;4.宽穗赖草;5.羊草;6.多枝赖草Plate Ⅱ Light microscopic micrographs on the lower epidermises of leaf blades of 24 Leymus species from China 1.L.paboanus; 2.L.angustus; 3.L.tianschanicus; 4.L.ovatus; 5.L.chinensis; 6.L.multicaulis

图版Ⅲ 国产赖草属24个物种的叶片下表皮光学显微照片 1.伊吾赖草;2.芒颖赖草;3.褐穗赖草;4.青海赖草;5.纤细赖草;6.细弱赖草Plate Ⅲ Light microscopic micrographs on the lower epidermises of leaf blades of 24 Leymus species from China 1.L.yiunensis; 2.L.aristiglumus; 3.L.bruneostachyus; 4.L.qinghaicus; 5.L.tenuis; 6.L.leptostachyus

图版Ⅳ 国产赖草属24个物种的叶片下表皮光学显微照片 1.皮山赖草;2.格尔木赖草;3.若羌赖草;4.弯曲赖草;5.胎生赖草;6.光洁赖草Plate Ⅳ Light microscopic micrographs on the lower epidermises of leaf blades of 24 Leymus species from China 1.L.pishanica; 2.L.golmudensis; 3.L.ruoqiangensis; 4.L.flexus; 5.L.viviparus; 6.L.mundus
然而,在国产赖草属24个物种中,上述这几类结构细胞的形态、数量和叶面的组合式样是不尽相同的,尤其是长细胞的长短和宽窄,气孔器的大小与分布、副卫细胞的形状,以及短细胞的丰缺和着生方式上具有较为明显的差异(表3~4和图版Ⅰ~Ⅳ)。这些差异不仅可以作为属下次级类群划分的参考依据,而且还可以作为本文推断属下类群间系统进化关系的重要依据。若就先前发表的国产赖草属3个组而言,多穗组植物通常长细胞短阔、细胞壁薄且平直,短细胞多呈孪生、马鞍形和圆球形,气孔器细胞体积小、副卫细胞多呈高圆顶形偶呈低圆顶形,刺毛主要分布于脉间、多呈钩,脉带接近脉间带;少穗组植物长细胞较长、细胞壁厚并呈疏波状弯曲,短细胞单生、常呈马鞍形偶呈圆球形,气孔器体积大、副卫细胞多呈低圆顶形或趋于平边形,刺毛多分布于脉上和脉间,脉带明显窄于脉间带;而单穗组植物的长细胞狭长、细胞壁厚多呈密波状弯曲,短细胞单生呈新月形和圆球形,气孔器细胞体积小、副卫细胞多呈平边形,无刺毛着生,脉带明显窄于脉间带。这些差异特征充分表明,先前界定的赖草属3个组群间具有明显的间断,理应隶属于不同的类群。同理,国产赖草属24个参试物种在上述这4类结构细胞上也存在着一定程度的差异,它们同样也具有类群划分和系统进化关系推断的作用和意义。
3 讨论
3.1 国产赖草属属内次级类群鉴分
就国产赖草属24个物种、72个居群外部形态特征的研究结果看,生殖器官的多数性状在类群间可以形成诸多间断,它们不仅造就了赖草属不同组的划分,而且还将该属分隔成了不同的物种。现综观国产赖草属的各种间断,我们发现较大的间断主要体现在花序直曲、小穗数、每小穗小花数等,这些性状具有较为重要的分类价值,完全可以将国产赖草属植物鉴分为不同的组群;相应,其它性状如花序长度、第一颖颖体长度、第一外稃长度及营养器官的一些性状,只能用于赖草属种级类群的区分。正是这样,本文仍然将国产赖草属划分为3组、24种,这与先前学者[2]报道的研究结果是一致的。
从国产赖草属参试物种叶表皮微形态的实验结果看,长细胞的长度、细胞壁的厚度和波曲程度,气孔器的体积和形状,以及刺毛的数量在参试类群间具有较大差异存在,并且这些性状在类群间所形成的最大间断与前面依据外部形态特征形成的类群间断极其相似,可以直接将国产赖草属区分为3个不同的组群;同样,国产赖草属所拥有的其它一些性状如长细胞细胞壁的厚度、短细胞的分布式样及气孔器的列数等,差异较小,与外部形态上的花序长度、第一颖颖体长度、第一外稃长度、叶片被毛状况等相似,只可用于种级水平类群的鉴分,据此本文对国产赖草属种级阶元划分的结果依然不变,与形态学研究结果完全相同。
3.2 国产赖草属属下类群演化关系
3.2.1 国产赖草属3组植物间的演化关系
在外部形态上,尽管能用于国产赖草属类群划分的性状较多,但具有演化意义且能较易确定演化趋势方向的性状却较少。我们发现,唯有花序的形状,穗轴同节小穗的数目、颖和外稃的形状及叶片的宽度上呈现一定的演化趋势,可以反映类群间的系统进化关系。本研究发现,国产赖草属多穗组植物穗状花序粗壮、外观呈锥形,穗轴同节常着生3-多枚小穗,颖呈宽披针形或披针形,外稃细狭、无芒或具芒尖;少穗组植物穗状花序细狭呈条状,穗轴同节着生2~3枚小穗,颖多呈窄披针形,外稃常宽阔、具芒尖或短芒;单穗组植物穗状花序细弱呈线状,穗轴各节仅着生1枚小穗,颖通常锥刺形,外稃略宽、常具短芒。因此,若按照小麦族植物穗状花序可以由繁到简演变的理论[18],则不难看出多穗组应是国产赖草属中最原始的组群,少穗组应是多穗组之上较高级的组群,而单穗组应是属中最高级的组群。其次,在类群亲缘上,多穗组与少穗组物种的穗状花序均较密集,穗轴同节着生2枚以上小穗,颖均显披针形等,示意两者具有较近的亲缘关系,尤其是它们的穗状花序较粗这一独有特征暗示两者可能具有直接的派生渊源;相应,少穗组和单穗组植物的穗状花序较狭窄、穗轴先端和基部均着生1枚小穗等特征,也同样表明两者可能存在直接的渊源关系;相反,多穗组和单穗组植物则具有较远的亲缘关系,原因在于其除了在外稃先端芒长、颖片和外稃脉数上类似外,其余形态特征相差较远。此外,我们认为少穗组可能处于性状演化的中间状态,充当了多穗组和单穗组演化的过渡桥梁。
国产赖草属3植物的这一系统进化关系若是印证在叶表皮微形态上,其关系也是十分清楚。先前学者[16,19]研究曾认为,赖草属中壁薄、平直、短锉的长细胞是初生类型,壁厚、波曲、展长的长细胞是次生类型;初生类群短细胞通常成排联生,演化趋势为马鞍形→新月形→圆球形;气孔器副卫细胞按照高圆顶副卫细胞→低圆顶副卫细胞→平边形副卫细胞方向而演化;刺是刺毛中的初生类型,钩是刺毛中的较高级类型,钩状毛是刺毛中的最高级类型。印证在类群演化上,长细胞短阔、细胞壁平直,短细胞孪生、呈马鞍形和圆球形,副卫细胞常为高圆顶形,脉间刺毛多为钩的多穗组原始;长细胞较长、细胞壁疏波状弯曲,短细胞单生、常呈马鞍形,副卫细胞为低圆顶形或趋于平边形,表皮具刺毛少穗组应是多穗组之上较进化的组群;而长细胞狭长、细胞壁密波状弯曲,短细胞单生、呈新月形和圆球形,副卫细胞多呈平边形,表皮无刺毛的单穗组最进化。在类群亲缘上,演化居中的少穗组在长细胞延伸、短细胞类型和着生方式以及脉带与脉间带相对宽度上均与单穗组相近,特别是短细胞单生这一独有特征暗示两者间可能具有直接的派生渊源;同时,其与多穗组仅在长细胞细胞壁的波曲、副卫细胞形状上具有某些相似,表明两者间可能具有相对较近的亲缘关系,尤其是多穗组长细胞细胞壁平直,少穗组植物长细胞细胞壁往往微弯,也进而暗示两者可能存在直接的派生关系。这与上述利用外部形态特征获得的演化结论基本一致。
3.2.2 国产赖草属部分物种间的演化关系
我们的研究还发现,上述国产赖草属外部形态和叶表皮微形态特征还可进一步推证属下参试物种间的系统进化关系,尤其是对物种演化级次上的确认是较为容易的。对实验过的物种而言,多穗组中大赖草穗轴同节着生4~6枚小穗,每小穗含3~5小花,颖线状披针形,长细胞短阔、细胞壁平直,短细胞呈狭马鞍形和圆球形,显然以性状的演化趋势来判断,大赖草应是该组最原始的类群;相反,柴达木赖草穗轴同节着生3~5枚小穗,每小穗含5~10小花,颖狭披针形,长细胞稍长、细胞壁稍弯曲,短细胞呈马鞍形或圆球形,可能是该组最高级的类群;而多穗组中硕花赖草和粗穗赖草穗轴同节常着生3~6枚小穗,每小穗常含4~7小花,颖线状披针形,长细胞细胞壁平直或微弯,短细胞呈狭马鞍形或圆球形,这些性状多少处于初生与次生的中间状态,其演化水平自然应介于前两者之间。在类群亲缘上,柴达木赖草和粗穗赖草除仅在外稃长度,外稃脉数目,长细胞长宽和气孔器体积上稍微差异外,其余特征基本相似或类同,足以表明两者的亲缘关系可能较近;而大赖草和粗穗赖草的主要区别也仅仅体现在第一颖颖体长短,长细胞长宽等极少数性状特征上,这也同样揭示了它们可能具有极为密切的亲缘关系。
国产赖草属少穗组种数较多,性状差异较大,演化水平参差不齐。其中,青海赖草穗轴同节着生3~4枚小穗,每小穗含4~7小花,颖线状披针形,长细胞稍短、壁波状弯曲,副卫细胞高圆顶形,应是该组最原始的类群;相反,纤细赖草穗轴同节着生2~3枚小穗,每小穗含2~3小花,颖锥形,长细胞稍长、细胞壁密波状弯曲,副卫细胞平边形,则可能是该组演化层次最高的类群;而另外其它类群如芒颖赖草、多枝赖草、褐穗赖草、青海赖草、赖草、羊草、宽穂赖草、窄颖赖草、光洁赖草及刺状赖草等,它们的性状多处于初生与次生的两极之间,应是该组演化层次居中的类群。在亲缘关系上,从类群相互间所具的性状来看,一些类群亲缘关系较近,如多枝赖草和弯曲赖草同具微弯、疏松的穗状花序,穗轴同节着生2~3枚小穗,长细胞较长、细胞壁密波状弯曲,副卫细胞平边形等,说明两者具有较近的亲缘关系;毛穗赖草和胎生赖草的花序直立、密集,穗轴同节着生2~3枚小穗,颖呈锥状,长细胞明显延长、壁波状弯曲,短细胞单生,也表明两者的亲缘关系可能较近;同样,窄颖赖草与芒颖赖草、细弱赖草与纤细赖草、天山赖草与胎生赖草、光洁赖草与细弱赖草等各对类群间,形态和叶表皮微形态性状极为相似,暗示这些类群间理应具有相对较近的亲缘关系。
尽管单穗组仅含3种国产赖草属植物,但这3个物种的性状并非完全相同,从而致使它们的演化水平各有上下。其中,格尔木赖草每小穗常含3小花,颖体和稃体相对较短,长细胞较短、细胞壁波状弯曲,脉带窄于或等于脉间带,可能是该组中最原始的类群;皮山赖草每小穗含2~3小花,颖体和稃体相对较长,长细胞最长、细胞壁密波状弯曲,脉带显著窄于脉间带,应是该组中最进化的类群;若羌赖草所具性状,多少处于上述两者之间,它的演化水平自然居中。在亲缘关系上,格尔木赖草和若羌赖草的穗状花序直立、疏松,穗轴同节仅着生1枚小穗,每小穗含3~5小花,长细胞较长、细胞壁密波状弯曲,副卫细胞平边形,说明两者的亲缘关系可能较近;同样,若羌赖草和皮山赖草具有直立、疏松的穗状花序,穗轴同节仅着生1枚小穗,颖常呈锥形,内稃近等于外稃,叶片正面和边缘粗糙或被柔毛、背面平滑无毛,长细胞较长、细胞壁密波状弯曲,短细胞单生、新月形和圆球形,副卫细胞平边形,脉带窄于脉间带,预示两者在该组内可能具有相对较近的亲缘关系;相反,格尔木赖草和皮山赖草所拥有的外部形态和叶表皮微形态的共同性状特征相对较少,仅在穗状花序的直曲,穗轴同节小穗的数目,长细胞的长度与细胞壁的弯曲程度等少数性状上相同,其余多数性状特征存在着某种差异,这进而表明它们之间的亲缘关系相对较远。
1.杨瑞武.赖草属植物的系统与进化研究[D].成都:四川农业大学,2003.
2.蔡联炳,苏旭.国产赖草属的分类修订[J].植物研究,2007,27(26):652-660.
3.陈守良,李德铢,朱光华,等.《Flora of China》[M].Beijing:Science Press,2006,22:387-394.
4.杨瑞武,周永红,郑有良,等.赖草属三个八倍体和两个十二倍体物种的核型研究[J].草业学报,2004,13(2):99-105.
5.Hoehstetter C F.Nachtraglicher Commentar zu meiner Abhandlung:“Aufbau der Graspflanze etc.”[J].Flora,1848,7:105-118.
6.Nevski S A.Agrostologische Studien IV.Uber das System der Hordeeae Benth.[J].Trudy Bot Inst Akad Nauk SSSR,Ser 1,FI Sist Vyss Rast,1933,1:9-32.
7.Pilger R.Addimenta Agrostologica.I.Triticeae(Hordeae)[J].Botanisehe Jahrbticher,1949,74:1-27.
8.Tzvelev N N.Tribe 3.Triticeae Dumort[M].// Fedorov A A.Poaceae URSS.Leningrad:Navka Publishing House,1976:176-189.
9.Löve A.Conspectus of the Triticeae[J].Feddes Repertorium,1984,95(7):425-521.
10.Dewey D R.The Genomic System of Classification as a Guide to Intergeneric Hybridization with the Perennial Triticeae[M].//Gustafson J P.Gene Manipulation in Plant Improvement.New York:Plenum Press,1984:209-279.
11.耿以礼.中国主要植物图说——禾本科[M].北京:科学出版社,1959:429-434.
12.郭本兆.中国植物志:第9卷,第3分册[M].北京:科学出版社,1987:15-22.
13.刘尚武.青海植物志:第4卷[M].西宁:青海人民出版社,1999:103-108.
14.颜济,杨俊良.中国赖草属新植物[J].云南植物研究,1983,5(3):275-276.
15.蔡联炳,张梅纽.国产赖草属的叶表皮特征与组群划分[J].植物研究,2005,25(4):400-405.
16.刘玉萍,苏旭,何懿涵,等.国产赖草属3组植物叶表皮解剖特征及其系统关系[J].植物研究,2013,33(6):644-652.
17.Metcalfe C R.Anatomy of the Monocotyledons Vol.1 Gramineae [M].London:Oxford Clarendon Press,1960:1-713.
18.郭本兆,王世金.我国小麦族的花序形态演化及其属间亲缘关系的探讨[J].西北植物研究,1981,1(1):12-19.
19.智丽.赖草属(禾本科)的分类、演化及地理分布的研究[D].西宁:中国科学院研究生院,2000.
BiosystematicRelationshipsamongTaxabelowLeymus(Poaceae)fromChinaBasedontheExternalMorphologyandMicromorphologicalCharacteristicsofLeafEpidermis
LI Zhao-Meng ZHOU Yong-Hui LIU Yu-Ping SU Xu*
(Key Laboratory of Medicinal Animal and Plant Resource in the Qinghai-Tibetan Plateau,Key Laboratory of Physical Geography and Environmental Process in Qinghai Province,Key Laboratory of Education Ministry on Environments and Resources in the Qinghai-Tibetan Plateau,School of Geography and Life Science,Qinghai Normal University,Xining 810008)
We analyzed the main character differences of seventy-two reprehensive populations from twenty fourLeymusspecies previously reported by scholars by morphological observation and anatomical analysis of leaf epidermises. There are different degrees of difference in fifteen morphological characters including length and width of flag leaves, hairs of leaf blades, straight/curve and length/width of inflorescence, spikelet No. of each inflorescence, floret No. of each spikelet, length of lemma and palea. The tested species can be divided into three sections according to them. The sect. Racemosus species ofL.racemosus,L.crassiusculus,L.pluriflorusandL.pseudoracemosus, with sturdy, erect and thick spike, 3-number of spikelets per rachis node, 3-10 florets each spikelet, linear lanceolate glume, etc., is the most primitive one in genusLeymus. Due to the sect. Anisopyrum species includingL.pishanica,L.golmudensisandL.ruoqiangensis, having thin, erect and loose spike, only one spikelet per rachis node, usually 2-5 florets each spikelet, linear lanceolate or cone glume, etc., so it is the most advanced of them. Meanwhile, because the external morphological characters of the sect.Leymusspecies usually lie in the intermediate transition state between the first two groups, naturally its systematics position should also be between the sect. Racemosus and sect. Anisopyrum species. At the same time, based on the evolutionary trends of the external morphological characters of three above sections, we deduced that the sect. Racemosus might produce immediately the sect.Leymus, whereas the sect. Anisopyrum might derive immediately form the sect.Leymus. The leaf epidermis ofLeymusfrom China consists of long cell, stomatal cell, short cell and prickle hair. It belongs to the typical festucoid type. Except that some characters, such as the type of long-cells and thick/thin of cell walls, the volume and type of guard cells from stomatal apparatus, have obvious overlap and show similar, other characters, including the length of long-cells and curve extent of cell walls, distribution pattern of short-cells, shape of subsidiary cells, and type of prickle-hairs, have obvious differences. They can delimitateLeymusspecies into three sections identical with the morphology. According to the evolutionary trends of leaf epidermal characteristics of three sections and species within each section, their evolutionary positions and relationships were inferred. The results show that the sect. Racemosus is the most primitive one among three sections, the sect.Leymusis slightly more advanced than the former, and the sect. Anisopyrum is the most advanced of them. The sect. Racemosus might produce immediately the sect.Leymus, whereas the sect. Anisopyrum might derive immediately form the sect.Leymus. The biosystematic relationships of taxa below the genus are corroborated by the evolutionary trend of external morphology.
Poaceae;Leymus;morphological character;morphological of leaf epidermis;biosystematic relationship
国家自然科学基金项目(31260052);青海省自然科学基金项目(2014-ZJ-947Q);青藏高原药用动植物资源重点实验室(2015-Z-Y0b)资助
李兆孟(1988—),男,硕士研究生,主要从事植物系统进化与分子生态学研究。
* 通信作者:E-mail:xusu8527972@126.com
2015-09-10
Q949.71
A
10.7525/j.issn.1673-5102.2016.01.003
