IL-6跨信号转导作用与其相关疾病的研究进展
2016-11-06侯小飞综述高方友审校贵州医科大学贵州贵阳550004贵州省人民医院贵州贵阳55000
侯小飞 综述 高方友 审校(.贵州医科大学,贵州贵阳550004;.贵州省人民医院,贵州贵阳55000)
·综述·
IL-6跨信号转导作用与其相关疾病的研究进展
侯小飞1综述 高方友2审校
(1.贵州医科大学,贵州贵阳550004;2.贵州省人民医院,贵州贵阳550002)
IL-6在机体内具有重要作用,其信号转导主要通过两种途径即经典信号转导和跨信号转导途径进行。经典信号转导主要与IL-6膜特异性受体结合激活下游信号,跨信号转导是IL-6与游离的sIL-6R结合激活下游信号。由于经典信号转导途径IL-6受体的局限性,其作用并不广泛,本文主要就IL-6跨信号转导途径在疾病中的作用做一简单综述。
白细胞介素6;跨信号转导;经典信号转导;相关疾病;进展
白细胞介素6(IL-6)是一种含有184个氨基酸的螺旋蛋白,1986年由Kishimoto等首先从T细胞中获取cDNA克隆。它主要由抗原递呈细胞如B细胞、巨噬细胞、树突状细胞分泌,另外某些非专业性抗原递呈细胞如神经胶质细胞、上皮细胞、内皮细胞、某些肿瘤细胞等也可进行分泌。IL-6在机体许多活动中充当重要角色,不仅活跃在调控免疫系统和神经系统中,而且在肝细胞再生、成骨细胞生长和新陈代谢等方面都有参与。目前已知,IL-6信号转导途径主要有两种形式,分别为经典信号转导和跨信号转导途径。由于受ILv6受体局限性的影响,经典信号转导途径作用并不广泛,本文主要就ILv6跨信号转导途径在疾病中的作用做一简单综述。
1 经典信号转导途径及其受体
1.1 IL-6Ra IL-6Ra是IL-6的膜特异性结合受体,由468个氨基酸组成,分子量为80 kDa。在经典信号转导过程中,IL-6需与两种不同结构的糖蛋白膜受体结合,其中一种为IL-6Ra(另一种为IL-6Rβ,即gp130)。IL-6Ra分布较为局限,主要存在于肝细胞及造血细胞膜表面[1-2],因此导致IL-6的经典信号转导途径较为局限。
1.2 gp130 gp130在组织内的表达无处不在,分子量是130 000。gp130又称为IL-6Rβ,它并不单独与IL-6或IL-6Ra结合,只有在IL-6激活细胞膜上的IL-6Ra受体形成复合物后才能与gp130结合来引起下游信号的传导。此外,gp130除与IL-6具有高亲和力外,它也是IL-11、IL-27、白血病抑制因子(LIF)、睫状神经营养因子(CNTF)等受体所共用的亚单位。
1.3 信号转导过程IL-6与细胞膜上的IL-6Ra结合后引起构象改变,形成活化的IL-6R复合物,这一复合物使同来结合的两个gp130二聚体化。gp130的二聚体化可以激活细胞内两种信号的传导路径:JAK途径与Ras-Raf信号路径,细胞内信号JAK途径磷酸化信号转导和转录活化蛋白(STAT)[3-4](图1)。STAT3的作用较为广泛,与调节细胞凋亡、血管发生、炎症反应以及急性期反应都有一定联系。但是由于IL-6Ra分布的局限性,此种信号传导路径主要存在于肝细胞及造血细胞表面,而缺乏IL-6膜特异性结合受体的细胞主要通过跨信号转导途径来进行信号的传导。IL-6经典信号转导途径除了在炎症过程中发挥作用,它还具有协调内环境稳定的性质,它在中性粒细胞增减、胆固醇变化和增加体重方面都有重要角色[5]。
2 跨信号转导途径及其受体
2.1 sIL-6R可溶性IL-6受体(Soluble IL-6R,sIL-6R)分子量为50 kDa,其产生机制主要有两种:IL-6Ra蛋白水解和mRNA选择性剪接,前者是产生的主要途径。在IL-6Ra蛋白水解过程中,解整联蛋白和金属蛋白酶(ADAM)17起到了重要作用,它使IL-6Ra从细胞膜上脱落并对其进行水解形成sIL-6R,所以提高这种蛋白酶的含量会使机体sIL-6R含量上升[6]。sIL-6R在机体中以游离的形式存在,分布范围较广,而且它与IL-6结合的亲和力和IL-6Ra相当,所以当细胞膜缺乏IL-6Ra时IL-6主要与sIL-6R结合形成复合物进行下一步反应。
2.2 sgp130 sgp130是gp130的一种可溶性形式,最初的发现是在循环系统中。后来在风湿性关节炎患者的尿液、血浆及滑液中发现了一种结构形式更为简单的sgp130,这种形式结构的sgp130被称为gp130-RAPS,分子量约为50 000。sgp130的形成过程与sIL-6的形成过程并不相似,它仅通过选择性剪接的mRNA翻译获得而无蛋白酶水解途径。有人设计并合成了一种sgp130的Fc段融合蛋白(sgp130Fc),通过实验证明,sgp130和sgp130Fc均可阻滞IL-6跨信号转导途径,但是对IL-6经典信号转导途径没有明显抑制作用。更有趣的是,虽然gp130也是IL-27、CNTF、LIF、OSM受体所共用的亚单位,但至今并没有发现sgp130Fc对这些受体发挥相似抑制作用[7]。
2.3 信号转导过程细胞膜缺乏膜特异性结合受体(IL-6Ra)的细胞可以通过sIL-6R与IL-6的结合做出应答反应。IL-6/sIL-6R复合物通过激活靶细胞来表达gp130从而引起细胞内信号路径的传导(图1)。除了对STAT3的磷酸化外,跨信号转导作用与经典信号转导作用的路径与形式不尽相同[8]。由于sIL-6R在机体内以游离形式广泛存在,故机体内IL-6信号的转导大部分为跨信号转导。sgp130和sgp130Fc对这种跨信号转导路径产生抑制作用,使这种转导途径的各受体在正常生理条件下保持动态平衡。
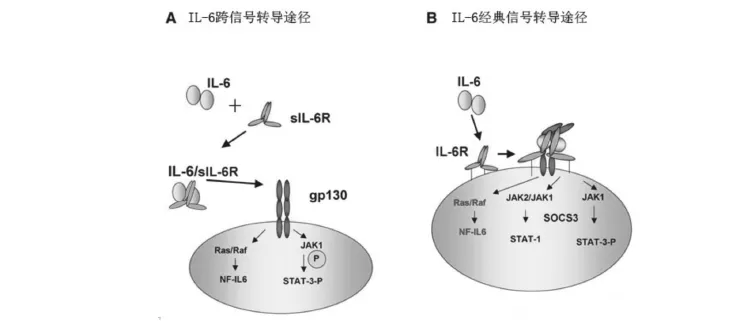
图1 IL-6跨信号转导途径和经典信号转导途径示意图
3 跨信号转导作用与疾病的关系
IL-6跨信号转导作用与多种疾病的发生机制有关,早前即有报道称它与风湿性关节炎、哮喘、炎性肠病等疾病相关,并且发现天然抑制因子sgp130可对这些疾病有明显的改善作用[9-11]。
3.1 中枢神经系统在中枢神经系统中,IL-6跨信号转导途径在多个方面发挥作用。由于sgp130可阻滞IL-6的跨信号转导作用而对经典信号转导途径无效,有人利用IL-6和IL-6/sgp130转基因小鼠模型(前者在中枢神经系统分泌较多IL-6,后者除IL-6外还分泌较多sgp130)对中枢神经系统中IL-6跨信号转导途径的作用进行研究。尽管这两种小鼠对IL-6的分泌水平相似,但是通过比较发现不仅某些已确定的通过IL-6跨信号转导途径进行应答的细胞和基因在两种模型鼠中表现不同,而且很多其他不确定通过IL-6哪种途径转导的各方面如星形胶质细胞增生、小胶质细胞增生、血脑屏障障碍、血管增生、神经退行性病变等也出现了差别:星形胶质细胞增生的出现主要与IL-6信号转导过程STAT3激活有关,其他一些gp130分子家族成员如LIF和CNTF也可以通过活化STAT3使星形胶质细胞应答。在实验中证实sgp130可阻滞IL-6这一过程,因此可以推测星形胶质细胞在IL-6信号转导过程中主要通过跨信号转导途径进行细胞应答。对于小神经胶质细胞来说,早期的研究证明其主要通过IL-6经典信号转导作用进行应答[12],但是后期发现在某些情形下会有sIL-6R的出现并对其进行调节,所以这种细胞可能有两种信号转导方式并存的可能性。对中枢神经系统中的某些基因来说,也存在着单一跨信号转导途径或与其他途径并存的现象。如Serpina3n在IL-6模型鼠中大量存在,但在IL-6/sgp130模型鼠中却寥寥无几,说明这种基因主要依靠跨信号途径进行转导,而神经中枢中存在的另一种基因Socs3却可能因多种转导途径的存在而并无类似表现。在血管方面虽然早期研究中有证实脑血管内皮细胞主要对IL-6经典信号转导途径进行应答,但在sgp130参与后,血管内皮细胞的增生发生了很大改变。在实验中IL-6对神经元的变性作用也因为sgp130的加入而有所缓解,说明神经退行性变与IL-6跨信号转导途径也有密切关系[13]。
3.2 循环系统IL-6在循环系统中占据重要作用,而且目前已有相关药物使用。托珠单抗是IL-6受体的人源化单克隆抗体,规范使用托珠单抗可有益于类风湿性关节炎患者的血管内皮功能并减轻大动脉僵硬现象[14]。由于血管壁的内皮细胞和平滑肌细胞膜缺乏IL-6R,因此IL-6跨信号转导作用在此发挥了主要作用。动脉粥样硬化也是循环系统中常见的疾病,国内外研究发现动脉粥样硬化斑块中含大量IL-6。IL-6通过跨信号转导作用进行自分泌来加速血管平滑肌的炎症反应,包括急性期蛋白表达和早期动脉粥样硬化形成过程中内皮细胞的增生和迁移。另外它还通过诱导表达趋化蛋白因子和粘附分子来募集内膜下免疫细胞促进内皮细胞活化,激活巨噬细胞完成其分化和迁移,这种分化迁移对促进动脉粥样硬化具有潜在作用[15]。刘艳红[16]研究认为,脂多糖在动脉粥样硬化进程中影响着IL-6的含量。另研究证明患有冠脉疾病的患者血浆中sgp130的水平含量较低[17],虽然sgp130低水平改变的具体病理机制还未知,但可猜想IL-6跨信号转导途径在冠脉疾病中占据重要位置。
3.3 呼吸系统在肺组织中,早前就有人证实IL-6在哮喘的呼吸道重建过程中有一定影响,它参与了诱导肺纤维母细胞向肌纤维母细胞转化的过程。它对肺纤维化的确定性作用首次由O'Donoghue等[18]提出,后经进一步研究发现,IL-6含量升高对肺纤维化疾病的发生和发展起到促进作用,它的跨信号转导作用在疾病发展过程中可引起细胞过度增殖及细胞外基质蛋白在肺中的产生和沉积。在研究过程中通过对sIL-6R的观察发现,sIL-6R的含量水平与疾病发展呈正相关,甚至可以将其看作为是肺纤维化疾病进展的一个标志物来衡量疾病发展程度。它在肺纤维化中呈区域分布,只发生在纤维化的肺组织当中。此外,由于机体主要依靠ADAM 17裂解IL-6Ra来产生sIL-6R,在实验中ADAM 17也表现出和sIL-6R类似的与疾病间的关系,因此有人提出通过针对以sIL-6 和ADAM 17为靶点进行研究可能为肺纤维化的治疗开辟新的途径[19]。
3.4 消化系统早前有报道称IL-6在炎性肠病中发挥重要作用[11],且针对这一途径已有相关药物应用于临床。最近研究发现在重症急性胰腺炎发展过程IL-6不仅参与到疾病病理发展中而且其含量可作为评价疾病进展程度的重要参考。IL-6与sIL-6R形成的复合物在胰腺中磷酸化STAT3,释放促炎因子增强炎症反应。STAT3持续不断地活化可引起CXCL1水平上升,CXCL1是一种中性粒细胞诱导剂,它的上升又可介导粒细胞渗入到肺内,继而引起致命性急性肺损伤[20]。另外有人对消化系统另一个重要脏器肝脏也进行了相关研究。早期他们通过实验发现IL6/ sIL-6R复合物可能对促进肝脏再生产生潜在作用,后续又发现肝损伤后sIL-6R水平立即升高。利用sgp130Fc融合蛋白阻滞IL-6跨信号转导作用时发现肝损伤严重程度明显上升,而且与肝损伤相关的氧化应激因子也随之升高。同期有人利用肝细胞STAT3基因敲除小鼠进行肝脏损伤实验发现与对照组相比,基因敲除小鼠的损伤程度要明显加重,而且肝细胞的再生能力也显著下降[21]。说明在肝脏损伤与再生的过程中IL-6跨信号转导途径起到了重要作用(虽然IL-6R主要存在于肝细胞表面),这一信号转导所引起的STAT3激活可能是其关键因素。另外本实验也说明了IL-6跨信号转导途径对机体抵抗氧化应激也可能起到重要作用[22]。
3.5 其他在骨质吸收过程中IL-6可通过活化STAT3来提高成纤维细胞中NF-κB受体活化蛋白配体(RANKL)的表达[23],RANKL又可增强骨组织中破骨细胞的分化并提高其活性。破骨细胞分化和某些感染性疾病引起的骨质破坏过程中会产生一种辅助T淋巴细胞因子Th17。在牙周疾病的发展过程中,IL-6通过跨信号转导途径促进破骨细胞激活从而引起牙周组织宿主细胞Th17积累和RANKL的表达,使牙周损害呈进行性发展[24]。IL-6与肿瘤的关系也见于大量报道。它的信号表达可促进某些肿瘤尤其是与慢性炎症相关的肿瘤的发生,如炎症相关性结肠癌、胰腺癌、肝细胞癌等[25-27]。IL-6不仅可直接刺激细胞本身引起增殖,它还可抑制肿瘤浆细胞的凋亡。在多发性骨髓瘤中,血清IL-6和sIL-6R的水平与肿瘤严重程度密切相关[28],sIL-6R的含量更被认为是预测多发性骨髓瘤的一个重要因子[29]。同时,IL-6还是髓样抑制细胞扩增和募集的重要介导因子[30],在乳腺癌细胞转移过程中,通过增加IL-6和sIL-6R的分泌可加剧细胞的侵袭和远处转移[31]。另外IL-6跨信号转导作用还可刺激女性排卵,卵巢血管内皮细胞中高浓度的IL-6经跨信号转导途径可直接增加血管的通透性,同时也能刺激血管内皮生长因子的产生[32]。
4 IL-6跨信号转导作用展望
IL-6跨信号转导途径在疾病的发生、发展和预后等方面都有重要作用。由于其表达调节是在受体水平上进行,所以针对这一途径相关疾病的治疗可通过以受体作为靶点进行研究。目前已见关于利用阻滞IL-6跨信号转导途径来治疗相关疾病的报道,如利用sgp130Fc治疗大肠炎、风湿性关节炎等自身免疫性疾病[9,11]。但由于对IL-6信号转导途径的阻滞可能会损害其他器官,故针对这一途径的治疗方法需进一步研究。
[1]ChalarisA,GarbersC,Rabe B,etal.The soluble Interleukin 6 receptor∶generation and role in inflammation and cancer[J].Eur JCell Biol,2011,90(6-7)∶484-494.
[2]Scheller J,Chalaris A,Schmidt-Arras D,et al.The proand anti-inflammatory propertiesof the cytokine interleukin-6[J].Biochim BiophysActa,2011,1813(5)∶878-888.
[3]Reipschlager S.Toxin-induced RhoA activity mediates CCL1-triggered signal transducersand activatorsof transcription protein signaling[J].JBiolChem,2012,287(14)∶11183-11194.
[4]Birukova AA,Tian Y,Meliton A,et al.Stimulation of Rho signaling by pathologic mechanical stretch is a“second hit”to Rho-independent lung injury induced by IL-6[J].Am JPhysiolLung Cell Mol Physiol,2012,302(9)∶965-975.
[5]Jones SA,Scheller J,Rose-John S.Therapeutic strategies for the clinical blockade of IL-6/gp130 signaling[J].JClin Invest,2011,121 (9)∶3375-3383.
[6]Cesaro A,Abakar-Mahamat A,Brest P,et al.Differential expression and regulation of ADAM 17 and TIMP3 in acute inflamed intestinal epithelia[J].Am JPhysiol Gastrointest Liver Physiol,2009,296(6)∶281-294.
[7]Stefan Rose-John.IL-6 Trans-Signaling via the Soluble IL-6 Receptor∶Importance for the Pro-Inflammatory Activities of IL-6[J].International Journalof BiologicalSciences,2012,8(9)∶1237-1247.
[8]Scheller J,Chalaris A,Schm idt-Arras D,et al.The pro-and anti-inflammatory propertiesof the cytokine interleukin-6[J].Biochim BiophysActa,2011,1813(5)∶878-888.
[9]Cronstein BN.Interleukin-6∶a key mediator of system ic and local symptoms in rheumatoid arthritis[J].Bull NYU Hosp Jt Dis,2007, 65(Suppl1)∶11-15.
[10]Finotto S,Eigenbrod T,Karwot R,etal.Localblockade of IL-6R signaling induces lung CD4+T cellapoptosis in amurinemodel of asthma via regulatory T cells[J].Int Immunol,2007,19(6)∶685-693.
[11]Matsumoto S,Hara T,M itsuyama K,et al.Essential roles of IL-6 trans-signaling in colonic epithelial cells,induced by the IL-6/soluble-IL-6 receptor derived from lam ina propria macrophages,on the development of colitisassociated premalignant cancer in a murine model[J].JImmunol,2010,184(3)∶1543-1551.
[12]Lin HW,Levison SW.Context-dependent IL-6 potentiation of interferon-gamma-induced IL-12 secretion and CD40 expression in murineM icroglia[J].JNeurochem,2009,111(3)∶808-818.
[13]Iain L,Campbell ME,Sue LL,et al.Trans-Signaling Is a Dom inant Mechanism for the Pathogenic Actions of Interleukin-6 in the Brain [J].The Journalof Neuroscience,2014,34(7)∶2503-2513.
[14]Protogerou AD,ZampeliE,Fragiadaki K,etal.A pilotstudy of endothelial dysfunction and aortic stiffness after interleukin-6 receptor inhibition in rheumatoid arthritis[J].Atherosclerosis,2011,219(2)∶734-736.
[15]Dan Q,Jian L,ChiWL,et al.IL-6 in diabetes andcardiovascular complications[J].British Journal of Pharmacology,2014,171(5)∶3595-3603,3595.
[16]刘艳红.动脉粥样硬化进程中单核/巨噬细胞IL-1、IL-6的表达[J].海南医学,2015,26(19)∶2818-2820.
[17]Schuett H,Oestreich R,Waetzig GH,etal.Transsignaling of interleukin-6 crucially contributes to atherosclerosis inm ice[J].Arterioscler Thromb Vasc Biol,2012,32(2)∶281-290.
[18]O'Donoghue RJ,KnightDA,RichardsCD,etal.Genetic partitioning of interleukin-6 signalling inmice dissociates Stat3 from Smad3-mediated lung fibrosis[J].EMBOMolMed,2012,4(9)∶939-951.
[19]Thanh-Thuy TL,Harry KQ,Ernestina M,et al.Blockade of IL-6 trans signaling attenuates pulmonary fibrosis[J].J Immunol,2014, 193(7)∶3755-3768.
[20]Zhang H,Neuhöfer P,Song L,et al.IL-6 trans-signaling promotes pancreatitis-associated lung injury and lethality[J].J Clin Invest, 2013,123(3)∶1019-1031.
[21]Horiguchi N,Lafdil F,M iller AM,et al.Dissociation between liver inflammation and hepatocellular damage induced by carbon tetrachloride in myeloid cell-specific signal transducer and activator of transcription 3 gene knockout mice[J].Hepatology,2010,51(5)∶1724-1734.
[22]Gew iese-Rabsch J,Drucker C,Malchow S,et al.Role of IL-6 trans-signaling in CCl4 induced liver damage[J].Biochim ica et BiophysicaActa,2010,1802(11)∶1054-1061.
[23]Choe JY,Park KY,Park SH,etal.Regulatory effectof calcineurin inhibitor,tacrolimus,on IL-6/sIL-6R-mediated RANKL expression through JAK2-STAT3-SOCS3 signaling pathway in fibroblast-like synoviocytes[J].ArthritisResearch and Therapy,2013,15(1)∶R26.
[24]Hosokawa Y,Shindo S,Hosokawa I,et al.IL-6 trans-signaling enhances CCL20 production from IL-1β-stimulated human periodontal ligamentcells[J].Inflammation,2014,37(2)∶381-386.
[25]Grivennikov S,Karin E,Terzic J,et al.IL-6 and Stat3 are required for survivalof intestinalepithelialcellsand developmentof colitis-associated cancer[J].Cancer Cell,2009,15(2)∶103-113.
[26]Lang SA,Moser C,Gaumann A,et al.Targeting heat shock protein 90 in pancreatic cancer impairs insulin-like grow th factor-I receptor signaling,disrupts an interleukin-6/signal-transducer and activator of transcription 3/hypoxia-inducible factor-1alpha autocrine loop,and reduces orthotopic tumor grow th[J].Clin Cancer Res,2007,13(2)∶6459-6468.
[27]Park EJ,Lee JH,Yu GY,et al.Dietary and genetic obesity promote liver inflammation and tumorigenesis by enhancing IL-6 and TNF expression[J].Cell,2010,140(2)∶197-208.
[28]Jurczyszyn A,Czepiel J,Biesiada G,etal.HGF,sIL-6R and TGF-β1 play a significant role in the progression of multiple myeloma[J]. Journalof Cancer,2014,5(7)∶518-524.
[29]Tsirakis G,Pappa CA,Kaparou M,et al.The relationship between soluble receptor of interleukin-6 w ith angiogenic cytokines and proliferation markers in multiple myeloma[J].Tumour Biol,2013,34 (2)∶859-864.
[30]Gabrilovich DI,Ostrand-Rosenberg S,Bronte V.Coordinated regulation of myeloid cells by tumours[J].Nat Rev Immunol,2011,12 (12)∶253-268.
[31]Oh K,Lee OY,Shon SY,et al.A mutual activation loop between breast cancer cells and myeloid-derived suppressor cells facilitates spontaneousmetastasis through IL-6 trans-signaling in amurinemodel[J].BreastCancer Research,2013,15(5)∶1-16.
[32]Cope C.The role of IL-6 trans-Signaling in vascular leakage∶implications for ovarian hyperstimulation syndrome in amurinemodel[J]. JClin EndocrinolMetab,2013,98(3)∶539-545.
Research progress of IL-6 trans-signaling and its related diseases.
HOU Xiao-fei1,GAO Fang-you2.1.Guizhou Medical University,Guiyang 550004,Guizhou,CHINA;2.People'sHospitalofGuizhou Province,Guiyang 550002,Guizhou, CHINA
Interleukin-6(IL-6)plays an important role in the body,and its signal transduction ismainly displayed through two ways∶IL-6 classical signaling and IL-6 trans-signaling.In the classical signaling,IL-6mainly binds w ith IL-6 specificmembrane receptor to activate the downstream signaling.In the trans-signaling,IL-6 mainly binds w ith soluble IL-6R(sIL-6R)to activate the downstream signaling.IL-6 classical signaling is notw idely applied due to its limitation of the IL-6 receptor.Herewe review the rolesof IL-6 trans-signaling in diseases in thispaper.
IL-6;Trans-signaling;Classicalsignaling;Related diseases;Progress
R331.1+42
A
1003—6350(2016)16—2667—04
2015-12-10)
doi∶10.3969/j.issn.1003-6350.2016.16.030
国家自然科学基金(编号:81260200)
高方友。E-mail:946672705@qq.com
