肥大细胞对重组口蹄疫病毒VP1-VP4应答的蛋白质表达谱检测
2016-11-01王燕张莎莎王蕾刘雪芳张恩李娜高云欢张义明李丽敏王家鑫
王燕,张莎莎,王蕾,刘雪芳,张恩,李娜,高云欢,张义明,李丽敏,王家鑫
肥大细胞对重组口蹄疫病毒VP1-VP4应答的蛋白质表达谱检测
王燕,张莎莎,王蕾,刘雪芳,张恩,李娜,高云欢,张义明,李丽敏,王家鑫
河北农业大学动物医学院免疫学实验室,河北保定 071000
为揭示肥大细胞抗口蹄疫病毒VP1-VP4蛋白的天然免疫作用,以重组口蹄疫病毒VP1-VP4蛋白刺激小鼠腹腔肥大细胞 (Peritoneal mast cells, PMCs),用高通量ELISA芯片检测PMCs的蛋白质表达谱。结果显示,VP1-VP4蛋白刺激的PMCs (VP1-VP4组) 表达CCL19、L-selectin、CCL17和TNF-α的水平极显著低于对照组 (PMCs) (0.001),而VP1-VP4蛋白刺激经甘露糖受体(Mannose receptor, MR) 抑制剂预处理的PMCs (MR组) 表达CCL19、IL-15、IL-9、G-CSF和Galectin-1的水平则极显著高于对照组 (0.01),IL-10表达水平也有显著升高 (0.05)。MR组与VP1-VP4组相比,PMCs表达IL-10、IL-17、CCL20、IL-15、IL-9、L-selectin、CCL17、TNF-α和CCL19的水平极显著升高 (0.01),CCL21和G-CSF的表达也显著高于VP1-VP4组 (0.05)。生物信息学差异表达分析结果显示,与对照组相比,VP1-VP4组PMCs表达的L-selectin和CCL17为下调性差异表达蛋白(Log(ratio)≤–1)。MR组与VP1-VP4相比,PMCs表达的CCL20、CCL19、L-selectin和IL-15为上调性差异表达蛋白 (Log(ratio)≥1)。这表明,PMCs可自发分泌CCL19、L-selectin、CCL17和TNF-α,而VP1-VP4则对PMCs的天然免疫功能具有抑制作用。由于阻断MR后PMCs的蛋白质表达水平显著升高,所以VP1-VP4对小鼠PMCs的免疫抑制作用可能是由MR介导的。
肥大细胞,甘露糖受体,口蹄疫病毒,天然免疫,差异表达
近年来,关于肥大细胞在抗细菌和抗寄生虫感染中的作用报道很多,但对其抗病毒作用的研究相对较少[1-3]。肥大细胞吞噬能力很弱,主要通过细胞膜表面的模式识别受体与Fc受体识别病原体,从而启动天然免疫应答[4]。研究发现,肥大细胞可通过TLR3识别新城疫病毒,并释放MIP-1β、KC和RANTES,促进CD8+T细胞向感染部位迁移[5]。Al-Afif等发现,用少量的呼吸道合胞体病毒感染肥大细胞即可诱导其产生Ⅰ型干扰素、CXCL10、CCL4和CCL5[6]。Fukuda等发现,肥大细胞以RIG-I样受体 (RLR) 和MDA5受体识别水疱性口炎病毒,诱导其产生IFN-α和IFN-β,从而发挥抗病毒作用,但是TLR3与此作用无关,而且RLR和MDA5受体识别水疱性口炎病毒并不引起肥大细胞脱颗 粒[7]。在登革病毒感染早期,肥大细胞迅速脱颗粒,随后病毒通过RLR和MDA5受体激活肥大细胞,产生TNF-α、IFN-α、CCL5、CXCL12和CX3CL1,从而发挥其免疫监视作用[8]。有报道将肥大细胞脱颗粒成分加入流感病毒疫苗后免疫小鼠,可使小鼠抵抗致死量的流感病毒攻 击[9]。因此,肥大细胞成为抗感染免疫研究的新热点。口蹄疫病毒 (Foot-and-mouth disease virus,FMDV) 有7种血清型,主要通过呼吸道和消化道感染动物,但目前对肥大细胞在抗FMDV感染免疫中的作用尚不清楚。
口蹄疫是严重危害动物健康的急性传染病,目前仍无有效的防治措施[10-11]。灭活口蹄疫疫苗起效慢,免疫保护期短,不能防止感染的发生,甚至还促使部分被感染牛形成持续感染[12-13],且不同血清型口蹄疫疫苗彼此不能产生交叉免疫保护。口蹄疫基因工程疫苗不仅在临床上未能产生优于灭活疫苗的效果,而且还不时引起动物过敏或死亡[14-16]。因此,研制安全有效的口蹄疫疫苗仍是一项迫在眉睫的任务[17-18]。FMDV衣壳由VP1–VP4四种结构蛋白组成,研究发现,VP1既可刺激机体产生中和抗体,又可刺激机体产生对7种血清型FMDV具有交叉保护作用的CD8+T细胞应答[19];VP4则可被多种单体型MHC分子识别[20],故VP1和VP4被视为研制通用型口蹄疫疫苗的理想后备抗原。分布于皮肤和粘膜中的肥大细胞是机体抗感染免疫的第一线免疫细胞[1]。因此,我们通过基因重组制备VP1-VP4融合蛋白,以此加载肥大细胞,检测其所产生的细胞因子、趋化因子和生长因子等蛋白质功能分子,以揭示肥大细胞抗FMDV的天然免疫作用。
1 材料与方法
1.1 材料
10−12周龄的BALB/c小鼠购自北京维通利华实验动物技术有限公司 (合格证编号:11400700089374),饲养于独立通气笼 (IVC) 中。大肠杆菌DH5α感受态细胞、BL21 (DE3) 感受态细胞和离心柱型质粒提取试剂盒购自天根生化科技公司。限制性内切酶HⅠ、Ⅰ和Ⅰ购自TaKaRa公司,小鼠抗6×His单克隆抗体和AP标记的羊抗鼠IgG购自Abcam公司,Percoll分层液购自Pharmacia公司,DMEM培养液和opti-MEMⅠ低血清培养液购自Gibco公司,胎牛血清购自HyClone公司,PMCs培养液为含5%胎牛血清的DMEM完全培养液。甘露糖受体 (MR) 抑制剂购自Sigma公司,小鼠高通量ELISA芯片购自RayBiotech公司。含VP1基因全长的重组质粒pET32a-VP1和含VP4基因全长的pBluescript II SK(+)-VP4由本实验室构建[21]。
1.2 重组质粒pET32a--的构建与鉴定
对重组质粒pET32a-(核酸序列中含Ⅰ和HⅠ酶切位点) 和pBluescriptⅡ SK (+)-(核酸序列中含HⅠ和Ⅰ酶切位点) 进行HⅠ和Ⅰ双酶 切[21],回收目的片段,在16 ℃下连接过夜,用连接产物转化DH5α感受态细胞,挑取单克隆菌落接种于Amp-LB培养液中,37 ℃振荡培养12−16 h,抽提重组质粒并进行Ⅰ和Ⅰ双酶切鉴定,将鉴定为阳性克隆的重组菌送生工生物工程 (上海) 股份有限公司测序,并利用Blast软件进行同源性比较。
1.3 重组VP1-VP4蛋白的原核表达、纯化与鉴定
将重组质粒pET32a--转化BL21 (DE3) 感受态细胞,挑取单克隆接种于Amp-LB培养液中,按照质粒提取试剂盒说明书的方法抽提质粒,将双酶切鉴定正确的重组菌按1∶100的比例接种于Amp-LB液体培养液,37 ℃振荡培养至600=0.8−1.0。加入IPTG,30 ℃振摇,诱导培养5 h。离心菌液,弃上清,用PBS重 悬,加入5×SDS上样缓冲液,充分混匀后 12 000 r/min离心2 min,取上清液进行12% SDS-PAGE凝胶电泳。用蒸馏水洗涤3次凝胶,切除浓缩胶部分,放入4 ℃的含1 mmol/L DTT的0.25 mol/L KCl溶液中,4 ℃显色10 min至电泳条带清晰,切下目的条带,装入盛有PB缓冲液透析袋中洗脱,然后倒转电极电泳5 min,收集透析袋中的目的蛋白,进行SDS-PAGE,考马斯亮染色,或以小鼠抗His单克隆抗体为一抗,碱性磷酸酶标记的羊抗鼠IgG为二抗做Western blotting鉴定。若鉴定结果和预期相符,则以此方法为工作模式制备重组VP1-VP4蛋白,用微量核酸蛋白检测仪测定其浓度,计算出VP1-VP4蛋白的含量,置–20 ℃冰箱中保存备用。
1.4 小鼠腹腔肥大细胞的收集
以颈椎脱臼法处死小鼠,腹腔注射3 mL PMCs培养液,轻按腹部数次,收集腹腔液,室温下1 700 r/min离心5 min,弃上清。用70% Percoll分层液重悬细胞沉淀,然后滴加PMCs培养液,3 000 r/min离心15 min,弃上清,用PMCs培养液重悬细胞沉淀,1 700 r/min离心 5 min,弃上清[22],用PMCs培养液重悬沉淀细胞后进行计数和活力检测。将分离收集的PMCs制成涂片,按常规进行甲苯胺蓝染色,快速水洗,分色,在普通光学显微镜下进行纯度鉴定。
1.5 不同处理PMCs的蛋白质表达谱检测
将收集的PMCs按每孔2×105个/mL铺于12孔细胞板中,放于5% CO2培养箱培养3 h,然后改用opti-MEM低血清培养液培养4 h。将细胞板分为3区,每区3孔,第1区以15 μg/mL剂量VP1-VP4蛋白加载 (VP1-VP4组);第2区先用MR抑制剂甘露聚糖处理1 h,再以 15 μg/mL剂量加载VP1-VP4蛋白 (MR组);第3区不做任何处理,为空白对照组。于加载VP1-VP4蛋白24 h后收集上清液,用高通量ELISA芯片检测各组PMCs的蛋白质表达谱,然后用化学发光检测仪扫描芯片,将获取的图像信息用软件AlphaView SA3.4.0转换为灰度值。
1.6 统计学处理
2 结果与分析
2.1 重组质粒pET32a--的构建与鉴定
已知基因全长为639 bp,基因全长为207 bp,但由于的50–56 bp为核酸内切酶Ⅰ的作用靶点,所以琼脂糖电泳后可现3条带,依次为pET32a载体片段、基因和部分、180 bp的剩余片段 (图1A),这表明重组已成功插入载体中。对重组质粒的序列进行比对发现,插入序列与GenBank中登录号为AY333431的FMDV O isolate O/NY00的和序列完全一致。
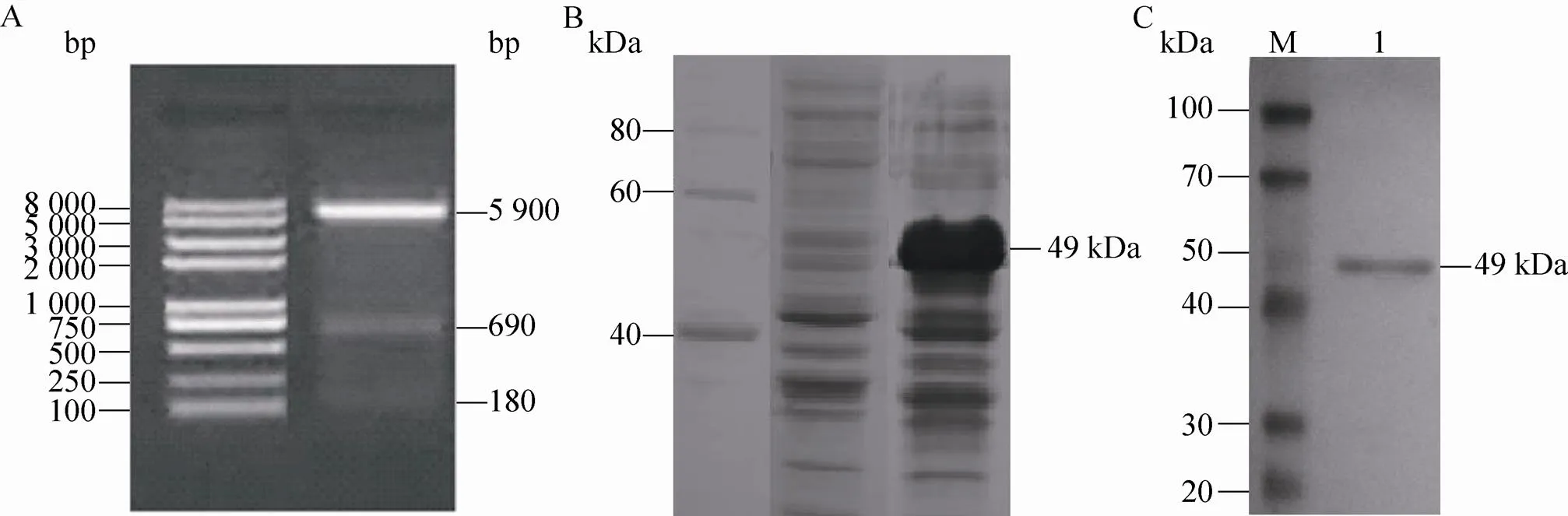
图1 重组VP1-VP4蛋白的鉴定
2.2 重组VP1-VP4蛋白的原核表达、纯化和鉴定
将IPTG加入重组菌液中,振摇诱导5 h,然后进行12% SDS-PAGE电泳,考马斯亮蓝染色,与未经诱导的重组菌 (图1B) 相比,在 49 kDa处可见与预期目的蛋白大小一致的条带 (图1B)。对电洗脱纯化后的VP1-VP4蛋白进行Western blotting,可见在49 kDa处出现一特异性条带 (图1C)。这表明重组VP1-VP4蛋白被成功制备。
2.3 小鼠腹腔肥大细胞的分离与鉴定
对用Percoll分层液分离得到的PMCs进行甲苯胺蓝染色,可见胞质呈紫红色,细胞膜轮廓清晰完整,无脱颗粒现象 (图2A),基本处于未活化状态,纯度为97%。全自动细胞活力检测显示细胞活力为98%。用VP1-VP4蛋白刺激PMCs可引起脱颗粒,此时细胞质着色变浅 (图2B),表明VP1-VP4蛋白可被PMCs识别,并引起活化。

图2 小鼠腹腔肥大细胞的鉴定(400×)
2.4 不同处理PMCs的蛋白质表达谱检测
与对照组相比,VP1-VP4组PMCs的CCL19、L-selectin、CCL17和TNF-α表达水平下降极为显著 (**0.01)。出乎意料的是,用MR抑制剂处理的PMCs不仅蛋白质表达水平没有下降,其所表达的CCL19、IL-15、IL-9、G-CSF和Galectin-1反而还极显著升高 (**0.01),IL-10也升高明显 (*0.05)。MR组与VP1-VP4组相比,PMCs表达IL-10、IL-17、CCL20、IL-15、IL-9、L-selectin、CCL17、TNF-α和CCL19的水平升高极为显著 (**0.01),CCL21、G-CSF也升高显著 (*0.05) (图3)。

图3 不同处理组PMCs分泌的主要蛋白质表达水平
2.5 不同处理PMCs的蛋白质表达
根据生物信息学差异表达分析原理,表达水平增高一倍,即Log(ratio)≥1为差异表达上调;表达水平下降一半,即Log(ratio)≤–1为差异表达下调[23]。与对照组相比,VP1-VP4组PMCs表达的L-selectin和CCL17为下调的差异表达。MR组与VP1-VP4相比,PMCs表达的CCL20、L-selectin、IL-15和CCL19为上调的差异表达。对不同处理组PMCs蛋白质表达量Log() 值进行聚类分析,将结果以热图 (Heat map) 表示。从该图可以看出,VP1-VP4组与对照组相比,PMCs表达的蛋白质水平呈下调 (绿色) 变化,而抑制MR的PMCs与对照组相比,PMCs分泌的蛋白质有1/3呈上调表达 (红色),与VP1-VP4组相比,抑制MR的PMCs分泌的蛋白质有2/3呈上调表达 (红色) (图4)。
图4 不同处理组PMCs蛋白质表达谱
3 讨论
尽管近年来对肥大细胞在抗病毒感染中的作用研究取得了一定进展[6-7],但对肥大细胞在抗FMDV感染中的作用仍不清楚。由于灭活FMDV疫苗的免疫诱导期长,7种血清型的灭活疫苗之间不能交叉保护,并且基于VP1蛋白的各种基因工程疫苗也只发挥短期免疫保护作用,所以深入研究机体抗FMDV的免疫应答机制,以筛选出最佳疫苗抗原就成为抗FMDV感染免疫研究的重要课题。大量研究证明,疫苗的有效免疫保护作用依赖中和抗体、活化的T细胞和免疫记忆[24]。FMDV的VP1可诱导动物产生中和抗体,并且O型FMDV的VP1可诱导具有交叉免疫保护作用的CD8+T细胞应答,故被视为研制通用型口蹄疫疫苗的理想后备抗原[19]。Van Lierop等研究发现,FMDV的VP4具有可被至少4种单体型MHC识别的T细胞抗原表位[20],故也被认为是研制通用型疫苗的理想后备抗原。因此,我们设想,将整个VP1蛋白与整个VP4蛋白连接在一起,有可能刺激动物产生高水平的中和抗体,有效激活T细胞,从而产生广谱免疫保护作用。为防止可能的移码发生,VP1与VP4之间未插入连接物。分布于皮肤和粘膜中的肥大细胞是机体抗感染免疫的第一线细胞[1]。因此,我们用重组VP1-VP4刺激PMCs,以揭示其在抗FMDV感染中的潜在作用。结果显示,与不加任何刺激的PMCs相比,负载VP1-VP4蛋白的PMCs表达CCL19、CCL17、L-selectin和TNF-α的水平显著下降,这表明PMCs可自发分泌趋化因子和炎症因子,而VP1-VP4蛋白则对PMCs具有免疫抑制作用。由于肥大细胞是发挥抗感染作用最迅速的免疫细胞,并可决定树突状细胞的抗原提呈方向[25],所以重组VP1-VP4对PMCs的抑制可能是FMDV形成急性感染的一种机制。
肥大细胞表达TLR、NLR、RLR、MR和清道夫受体 (Scavenger receptor,SR),藉此识别各种病原体相关分子模式,从而启动炎症反应和抗病毒免疫应答[26-27]。我们最近发现,大鼠肥大细胞系P815通过MR识别VP1-VP4可启动TNF-α分泌,而通过SR识别VP1-VP4则引起肥大细胞脱颗粒[28]。为进一步研究肥大细胞抗FMDV的天然免疫作用机制,我们用VP1-VP4分别加载经MR抑制剂预处理的PMCs和未作处理的PMCs,比较二者的细胞因子、趋化因子和生长因子等蛋白质功能分子的表达谱,结果发现,抑制PMCs的MR不仅没有降低CCL20、L-selectin、CCL17、TNF-α、IL-15、CCL19和CCL21等功能分子的表达,反而还显著升高。差异表达分析显示,CCL20、L-selectin、IL-15和CCL19为上调性差异表达。这表明,FMDV对机体的免疫抑制作用可能是通过MR介导的,该发现对于设计口蹄疫疫苗佐剂具有一定的指导意义。由于这些免疫分子在营造局部免疫应答环境中发挥重要的决定作用,故可作为评价疫苗的技术指标[29]。
上述结果表明,肥大细胞是对FMDV发挥免疫监视作用的天然免疫细胞,在抗感染早期发挥主要作用。同时本文也揭示了FMDV通过肥大细胞MR抑制机体免疫功能的现象,因此,将后备抗原VP1-VP4研制成通用型口蹄疫疫苗尚需进一步研究模式识别所启动的天然免疫应答。
REFERENCES
[1] Johnzon CF, Rönnberg E, Pejler G. The role of mast cells in bacterial infection. Am J Pathol, 2016, 186(1): 4–14.
[2] Hughes MR, McNagny KM. Mast Cells: Methods and Protocols. 2nd ed. New York: Humana Press, 2015: 93–119.
[3] Cao ZR, Yan WJ, Wang B, et al. Pattern recognition and activation effect of mast cellsinfected byS2. Chin J Cell Mol Immunol, 2013, 29(11): 1137–1140 (in Chinese). 曹志然, 闫伟娇, 王蓓, 等. 肥大细胞对布鲁菌S2株的模式识别及活化效应的体外研究. 细胞与分子免疫学杂志, 2013, 29(11): 1137–1140.
[4] Pinke KH, de Lima HG, Cunha FQ, et al. Mast cells phagocyteand produce nitric oxide by mechanisms involving TLR2 and Dectin-1. Immunobiology, 2016, 221(2): 220–227.
[5] Orinska Z, Bulanova E, Budagian V, et al. TLR3-induced activation of mast cells modulates CD8+T-cell recruitment. Blood, 2005, 106(3): 978–987.
[6] Al-Afif A, Alyazidi R, Oldford SA, et al. Respiratory syncytial virus infection of primary human mast cells induces the selective production of type I interferons, CXCL10, and CCL4. J Allergy Clin Immunol, 2015, 136(5): 1346– 1354.e1.
[7] Fukuda M, Ushio H, Kawasaki J, et al. Expression and functional characterization of retinoic acid-inducible gene-I-like receptors of mast cells in response to viral infection. J Innate Immun, 2013, 5(2): 163–173.
[8] St John AL, Rathore AP, Yap H, et al. Immune surveillance by mast cells during dengue infection promotes natural killer (NK) and NKT-cell recruitment and viral clearance. Proc Natl Acad Sci USA, 2011, 108(22): 9190–9195.
[9] St John AL, Chan CY, Staats HF, et al. Synthetic mast-cell granules as adjuvants to promote and polarize immunity in lymph nodes. Nat Mater, 2012, 11(3): 250–257.
[10] Saeed A, Kanwal S, Arshad M, et al. Foot-and-mouth disease: overview of motives of disease spread and efficacy of available vaccines. J Anim Sci Technol, 2015, 57: 10.
[11] Habiela M, Seago J, Perez-Martin E, et al. Laboratory animal models to study foot-and- mouth disease: a review with emphasis on natural and vaccine-induced immunity. J Gen Virol, 2014, 95(Pt 11): 2329–2345.
[12] Stenfeldt C, Eschbaumer M, Pacheco JM, et al. Pathogenesis of primary foot-and-mouth disease virus infection in the nasopharynx of vaccinated and non-vaccinated cattle. PLoS ONE, 2015, 10(11): e0143666.
[13] Elnekave E, Zamir L, Hamd F, et al. Risk factors for foot and mouth disease outbreaks in grazing beef cattle herds. Prev Vet Med, 2015, 120(2): 236–240.
[14] Li GZ, Wu LK, Mo RGGW, et al. Safety trial of foot-and-mouth disease vaccines of types O, A, and Asia I administration in sheep. J Anim Sci Vet Med, 2015, 32(4): 45–46 (in Chinese). 李国中, 乌兰扣, 莫日根高娃, 等. 羊两种三型口蹄疫疫苗免疫注射的安全性试验. 畜牧兽医杂志, 2015, 34(2): 45–46.
[15] Li YF, Miao SH, Zhu LX. Porcine immune response to foot-and-mouth disease vaccine and prevention measures. Chin Anim Hus Vet Med Abstr, 2013, 29(10): 108 (in Chinese).李艳粉, 苗绍华, 朱丽仙. 猪口蹄疫疫苗的免疫反应及防治措施. 中国畜牧兽医文摘, 2013, 29(10): 108.
[16] Rodriguez LL, Gay CG. Development of vaccines toward the global control and eradication of foot-and-mouth disease. Expert Rev Vaccines, 2011, 10(3): 377–387.
[17] Ruiz V, Mozgovoj MV, Dus Santos MJ, et al. Plant-produced viral bovine vaccines: what happened during the last 10 years? Plant Biotechnol J, 2015, 13(8): 1071–1077.
[18] Ludi A, Rodriguez L. Novel approaches to foot-and-mouth disease vaccine development. Dev Biol (Basel), 2013, 135: 107–116.
[19] Guzman E, Taylor G, Charleston B, et al. Induction of a cross-reactive CD8+T cell response following foot-and-mouth disease virus vaccination. J Virol, 2010, 84(23): 12375–12384.
[20] Van Lierop MJ, Nilsson PR, Wagenaar JP, et al. The influence of MHC polymorphism on the selection of T-cell determinants of FMDV in cattle. Immunology, 1995, 84(1): 79–85.
[21] Li N, Li LM, An PL, et al. T lymphocyte activation by dendritic cells pulsed with VP1-VP4 fusion protein of foot-and-mouth disease virus. Chin J Vet Sci, 2012, 32(6): 818–822 (in Chinese). 李娜, 李丽敏, 安鹏丽, 等. 负载口蹄疫病毒VP1-VP4融合蛋白质的树突状细胞对T细胞的活化效应. 中国兽医学报, 2012, 32(6): 818–822.
[22] Kovarova M. Isolation and characterization of mast cells in mouse models of allergic diseases//Allen IC, ED. Mouse Models of Allergic Disease. New York: Humana Press, 2013, 1032: 109–119.
[23] Sun ZQ, Xu YY. Medical Statistics. 4th ed. Beijing: People's Medical Publishing House, 2014: 472 (in Chinese). 孙振球, 徐勇勇. 医学统计学. 4版. 北京: 人民卫生出版社, 2014: 472.
[24] Pennock ND, Kedl JD, Kedl RM. T cell vaccinology: beyond the reflection of infectious responses. Trends Immunol, 2016, 37(3): 170–180.
[25] Dawicki W, Jawdat DW, Xu N, et al. Mast cells, histamine, and IL-6 regulate the selective influx of dendritic cell subsets into an inflamed lymph node. J Immunol, 2010, 184(4): 2116–2123.
[26] St John AL, Abraham SN. Innate immunity and its regulation by mast cells. J Immunol, 2013, 190(9): 4458–4463.
[27] Vukman KV, Ravidà A, Aldridge AM, et al. Mannose receptor and macrophage galactose-type lectin are involved inmast cell interaction. J Leukoc Biol, 2013, 94(3): 439–448.
[28] Li LM, Wang Y, Cui BB, et al. The effectiveness of mast cell pattern recognition of recombinant VP1-VP4 of foot-and-mouth disease virus. Chin J Anim Vet Sci, 2015, 46(9): 1644–1649 (in Chinese).李丽敏, 王燕, 崔贝贝, 等. 肥大细胞对重组口蹄疫病毒VP1-VP4蛋白的模式识别效应. 畜牧兽医学报, 2015, 46(9): 1644–1649.
[29] Moon TC, Befus AD, Kulka M. Mast cell mediators: their differential release and the secretory pathways involved. Front Immunol, 2014, 5: 569.
(本文责编 陈宏宇)
Protein expression profile of mast cells in response to recombinant VP1-VP4 of foot-and-mouth disease virus
Yan Wang, Shasha Zhang, Lei Wang, Xuefang Liu, En Zhang, Na Li, Yunhuan Gao, Yiming Zhang, Limin Li, and Jiaxin Wang
,,,071000,,
To reveal the innate immunity of mast cells against recombinant VP1-VP4 protein of foot-and-mouth disease virus (FMDV), mouse peritoneal mast cells (PMCs) were pulsed with recombinant VP1-VP4 protein. The supernatants harvested from PMCs cultures were applied to the high throughput ELISA array. Our results show that the expression levels of CCL19, L-selectin, CCL17, and TNF alpha released from PMCs pulsed with recombinant VP1-VP4 were significantly down-regulated compared with PMCs alone (0.001). Surprisingly, in comparison with PMCs alone, the expression levels of CCL19, IL-15, IL-9, G-CSF, and Galectin-1 in PMCs with the mannose receptor (MR) inhibitor were significantly up-regulated (0.01), and the expression level of IL-10 was also remarkably up-regulated (0.05). Importantly, the protein expression levels in PMCs treated with MR inhibitor were higher than PMCs pulsed with VP1-VP4, including IL-10, IL-17, CCL20, IL-15, IL-9, L-selectin, CCL17, TNF alpha, and CCL19 (0.01) as well as CCL21, and G-CSF (0.05). Differential expression analysis in bioinformatics shows that both L-selectin and CCL17 were recognized as differentially expressed protein molecules (Log(ratio)≤–1) when compared with PMCs alone. Furthermore, the up-regulation of the expression levels of CCL20, CCL19, L-selectin, and IL-15 in PMCs treated with MR inhibitor was defined as differential expression (Log(ratio)≥1). These data indicate that PMCs are capable of secreting CCL19, L-selectin, CCL17, and TNF alpha spontaneously and the recombinant VP1-VP4 has an inhibitive potential to PMCs during their performance of innate immune response. Given the protein expression levels from PMCs pre-treated with MR inhibitor were significantly increased, it can be deduced that immunosuppression of FMDV is presumably initiated by the VP1 recognition of MR on mast cells.
mast cells, mannose receptor, foot-and-mouth disease virus, innate immunity, differential expression
December 15, 2015; Accepted: April 15, 2016
Supported by:National Natural Science Foundation of China (No. 31402174), Comprehensive Reform Project of Veterinary Medicine in Universities, Hebei Province, Key Project of Hebei Education Department (No. ZD2015040).
Limin Li. Tel: +86-312-7528372; Fax: 86-312-7520275; E-mail: lilimin03@163.com Jiaxin Wang. Tel: +86-312-7528372; Fax: 86-312-7520275; E-mail: mastwang@163.com
国家自然科学基金(No. 31402174),河北省高等学校专业综合改革项目,河北省教育厅重点项目 (No. ZD2015040) 资助。
网络出版时间:2016-05-06 网络出版地址:http://www.cnki.net/kcms/detail/11.1998.Q.20160506.1637.002.html
王燕, 张莎莎, 王蕾, 等. 肥大细胞对重组口蹄疫病毒VP1-VP4应答的蛋白质表达谱检测. 生物工程学报, 2016, 32(9): 1194–1203.
Wang Y, Zhang SS, Wang L, et al. Protein expression profile of mast cells in response to recombinant VP1-VP4 of foot-and-mouth disease virus. Chin J Biotech, 2016, 32(9): 1194–1203.
