钒对雏鸡大脑皮层γ-氨基丁酸表达的影响
2016-11-01辛丹宁田春洋崔恒敏
唐 丽,辛丹宁,田春洋,崔恒敏
(四川农业大学动物医学院,成都 611130)
钒对雏鸡大脑皮层γ-氨基丁酸表达的影响
唐丽,辛丹宁,田春洋,崔恒敏*
(四川农业大学动物医学院,成都 611130)
为了研究钒对雏鸡大脑皮层γ-氨基丁酸(GABA)表达的影响,选用280只1日龄艾维因肉鸡,随机分为4组,每组7个处理,每个处理10只,分别饲喂对照日粮和钒添加日粮(低剂量组:15 mg·kg-1;中剂量组:30 mg·kg-1;高剂量组:45 mg·kg-1)6周,采用免疫组织化法检测雏鸡大脑皮层中GABA的表达变化。结果表明,42 d雏鸡,15 mg·kg-1钒添加组的大脑皮层GABA表达量最高(P<0.01或P<0.05),其次为30 mg·kg-1钒添加组的(P<0.01或P<0.05),45 mg·kg-1钒添加组的高于对照组的(P>0.05);45 mg·kg-1钒添加组,28 d时大脑皮层GABA表达量均极显著(P<0.01)高于14和42 d的;14 d时大脑皮层水平细胞和星形胶质细胞GABA表达量均高于42 d的(P>0.05)。综上表明,日粮钒与雏鸡大脑皮层GABA的表达存在量效与时效关系,提示其调控作用可能与GABA的表达上调相关。
钒;大脑皮层;γ-氨基丁酸;免疫组化法;雏鸡
钒是动物的必需微量元素之一,对维持机体生长发育、增强机体免疫力、消化能力、抗氧化能力及维持机体内环境恒定等起着重要的作用[1-9]。钒缺乏时,动物机体的生理功能会受到严重的影响;钒过量可引起动物机体的钒中毒[10-12],其中家禽对钒毒性最敏感。
γ-氨基丁酸(γ-aminobutyric acid,GABA)是中枢神经系统的一种重要的抑制性神经递质,对动物机体正常的生理功能起着重要的作用,具有调节机体消化摄食、增加免疫性能及抗氧化能力、调节内分泌等多种生理活性[13]。GABA在脑内的含量很高,脑内有超过30%的突触是以GABA 作为神经递质传递信息的[14]。GABA存在于大脑的多种中间神经元和投射神经元,参与调节突触可塑性,对兴奋扩散及传导过程中起负调节作用[15-17]。研究表明[18],大脑皮层星形胶质细胞能与从丘脑来的特异传入纤维(传入的上行纤维)形成突触;大脑皮层水平细胞的突起与锥体细胞顶树突联系,是大脑皮质区的局部(中间)神经元;大脑皮层锥体细胞的长轴突进入白质,组成投射纤维(下行至脑干或脊髓)或联合纤维(到同侧或对侧的另一皮质区),是大脑皮质的主要投射(传出)神经元。以上3种传递方式由此构成了皮质信息传递的复杂微循环[19]。
文献表明,氟和砷可引起大鼠大脑皮质GABA含量的变化,导致中枢神经功能紊乱[20]。R.Poggioli等[21]报道了钒染毒组大鼠出现明显的神经症状。目前,关于钒对雏鸡的研究多集中于消化、免疫和生殖系统等方面[1-9]。作者通过研究钒对雏鸡大脑皮层GABA表达的影响,初探其可能的调节机制,为钒在家禽饲养及提高抗应激能力中提供理论参考依据。
1 材料与方法
1.1动物
1日龄健康艾维茵肉鸡280只,购自四川省成都市温江正大畜禽有限公司。
1.2主要药物
偏钒酸铵、普通饲料(钒含量73 μg·kg-1)、4%多聚甲醛、PBS缓冲液、柠檬酸盐缓冲液、兔抗鸡多克隆抗体GABA(北京博奥森生物技术有限公司,抗体货号:bs-2252R,稀释倍数:1∶100)、二抗、SABC(兔IgG)、DAB显色剂、多聚赖氨酸(武汉博士德生物工程有限公司)等。
1.3主要仪器
101—2型电热鼓风干燥箱(上海仪表厂)、微波炉(格兰仕)、电子天平(沈阳龙腾电子有限公司)、石蜡切片机(Leica RM2135)、数显电热恒温水浴锅(上海医疗器械七厂WSZ-261-7) 、光学显微镜(Olympus)、图像分析系统(江苏省捷达科技发展有限公司)。
1.4方法
1.4.1实验动物模型的建立及分组将健雏280只,随机分为4组,每组70只,分别饲喂对照日粮和钒添加日粮(低剂量组:15 mg·kg-1;中剂量组:30 mg·kg-1;高剂量组:45 mg·kg-1),钒源为偏钒酸铵。饲养环境:温度(20±4) ℃,湿度(60±20)%室内整洁安静,通风良好,每笼饲养10只,每周更换垫料2次。常规管理,自由饮水和采食(标准基础饲料),试验期为6周。
1.4.2样本采集及处理雏鸡分别于14、28和42 d颈动脉放血致死,取脑置于4%多聚甲醛-0.1%磷酸缓冲液中固定24 h,制作光镜样本。
1.4.3样本制作取大脑(矢状切)石蜡包埋,连续切片(厚度为4~6 μm),SABC 法染色,OLYMPUS 显微镜观察并显微摄影。
1.4.4免疫组化染色程序按SABC试剂盒说明书操作。
1.4.5结果判定对照组切片染色为阴性(-),细胞内无阳性细胞产物。阳性细胞胞核出现棕黄色,根据显色程度分3级,并结合Image Pro Plus图像分析系统中的积分光度对结果进行判定:(+++)深棕色,强阳性;(++)中等着色,棕色,阳性;(+)着色浅,淡黄色,弱阳性。
1.4.6统计分析在10×40镜下,利用美国Image Pro Plus图像分析系统,每张切片按统一标准随机截取10个视野,测定阳性产物的平均光密度(OD)和积分光密度(IOD),利用SPSS13.0软件进行数据统计分析,以平均值±标准差表示,用多因素方差分析(ANOVA)差异显著性。P<0.05为差异显著,P<0.01为差异极显著。
2 结 果
GABA阳性产物为棕黄色或褐色,雏鸡大脑皮层神经元胞质和神经胶质细胞内均有阳性产物的分布(图1)。
2.1不同剂量钒对42日龄雏鸡大脑皮层GABA表达的影响
与对照组相比,试验组中雏鸡大脑皮层GABA表达量增多,15或30 mg·kg-1钒添加组GABA阳性信号高于或同于对照组;15或30 mg·kg-1钒添加组的三种细胞(大脑皮层水平细胞、星形胶质细胞和中型锥体细胞)的GABA表达量均显著(P<0.05)或极显著(P<0.01)高于对照组;45 mg·kg-1钒添加组的三种细胞的GABA表达量均与对照组差异不显著(P>0.05)。15 mg·kg-1钒添加组的三种细胞GABA表达量显著(P<0.05)或极显著(P<0.01)高于其他组(图1~2,表1)。
大脑皮层水平细胞:与其它组相比,15 mg·kg-1钒添加组GABA平均光密度与平均积分光密度最高(P<0.01);30 mg·kg-1钒添加组GABA的平均积分光密度极显著(P<0.01)低于15 mg·kg-1钒添加组的,显著高于(P<0.05)高于45 mg·kg-1钒添加组和
对照组的;45 mg·kg-1钒添加组和对照组GABA的平均积分光密度差异不显著(P>0.05)。
星形胶质细胞:15 mg·kg-1钒添加组GABA的平均积分光密度最高,极显著高于(P<0.01)高于45 mg·kg-1钒添加组和对照组的,与30 mg·kg-1钒添加组的差异不显著(P>0.05);30 mg·kg-1钒添加组GABA的平均积分光密度极显著高于(P<0.01)高于45 mg·kg-1钒添加组和对照组的,其GABA平均光密度高于其他试验组的;45 mg·kg-1钒添加组和对照组GABA的平均积分光密度差异不显著(P>0.05)。
锥体细胞:15 mg·kg-1钒添加组GABA的平均积分光密度最高,极显著(P<0.01)或显著高于(P<0.05)其他组的;30 mg·kg-1钒添加组GABA的平均积分光密度显著高于(P<0.05)高于45 mg·kg-1钒添加组和对照组的,其GABA平均光密度高于其他试验组的。
表1不同剂量钒对42日龄雏鸡大脑皮层GABA表达的影响
Table 1Effect of vanadium on the expression of GABA in the cerebral cortex of 42-day-old chickens
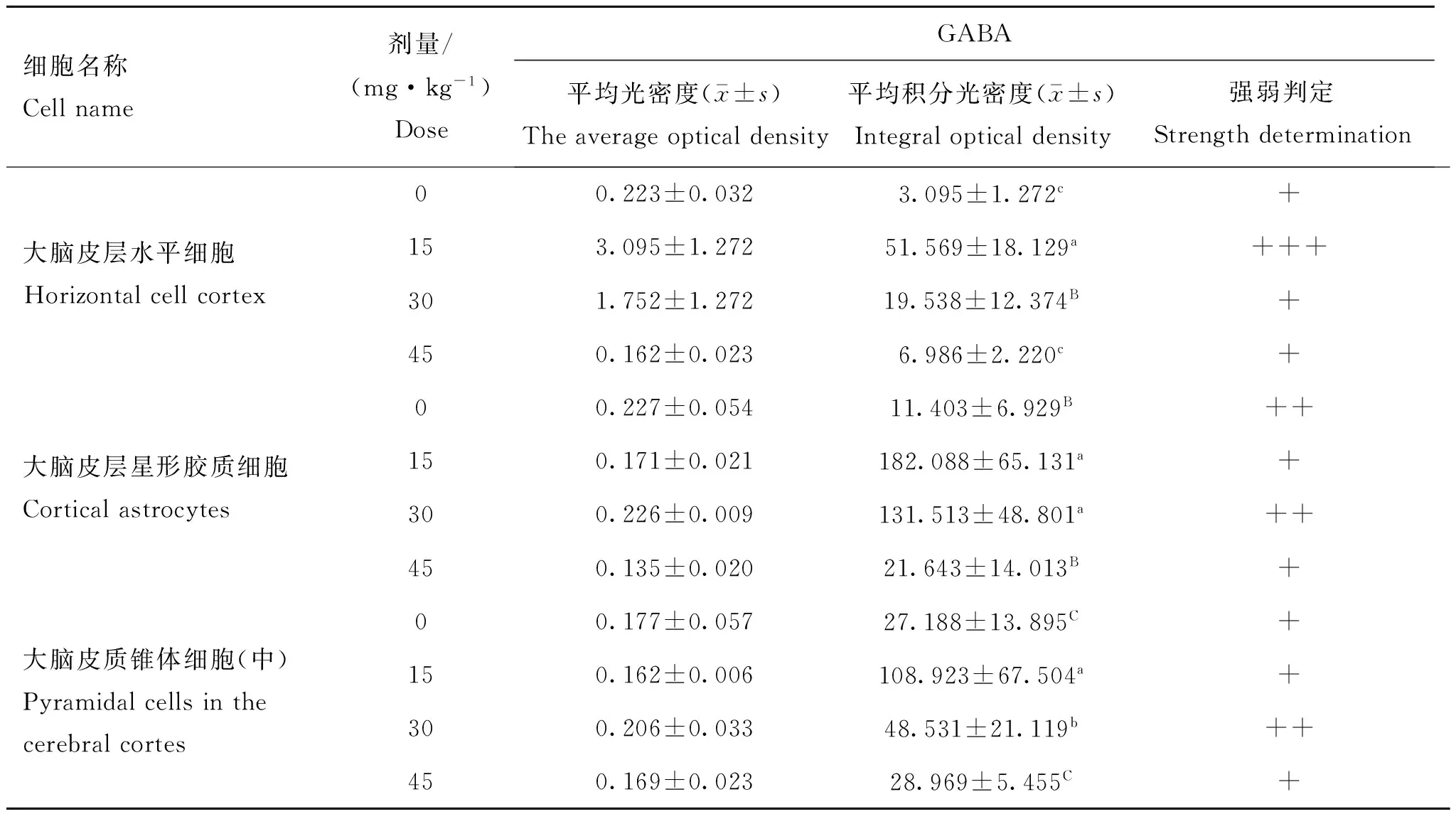
细胞名称Cellname剂量/(mg·kg-1)DoseGABA平均光密度(x-±s)Theaverageopticaldensity平均积分光密度(x-±s)Integralopticaldensity强弱判定Strengthdetermination大脑皮层水平细胞Horizontalcellcortex00.223±0.0323.095±1.272c+153.095±1.27251.569±18.129a+++301.752±1.27219.538±12.374B+450.162±0.0236.986±2.220c+大脑皮层星形胶质细胞Corticalastrocytes00.227±0.05411.403±6.929B++150.171±0.021182.088±65.131a+300.226±0.009131.513±48.801a++450.135±0.02021.643±14.013B+大脑皮质锥体细胞(中)Pyramidalcellsinthecerebralcortes00.177±0.05727.188±13.895C+150.162±0.006108.923±67.504a+300.206±0.03348.531±21.119b++450.169±0.02328.969±5.455C+
同列标注不同小写字母表示差异显著(P<0.05),不同大写字母表示差异极显著(P<0.01)
A column,treatment means without a common miniscule letter differs significantly(P<0.05),within a coulumn,treatment means without a common capital letter differs too much significantly (P<0.01) and the same
2.245 mg·kg-1钒添加量对不同日龄雏鸡大脑皮层GABA表达的影响
钒添加量为45 mg·kg-1时,28 d时三种细胞(大脑皮层水平细胞、星形胶质细胞和中型锥体细胞)GABA表达量均极显著(P<0.01)高于14和42 d的;14 d的三种细胞GABA表达量高于42 d的,但差异不显著(P>0.05)。28 d时大脑皮层水平细胞GABA平均光密度均高于14 d和42 d的(图2,表2)。
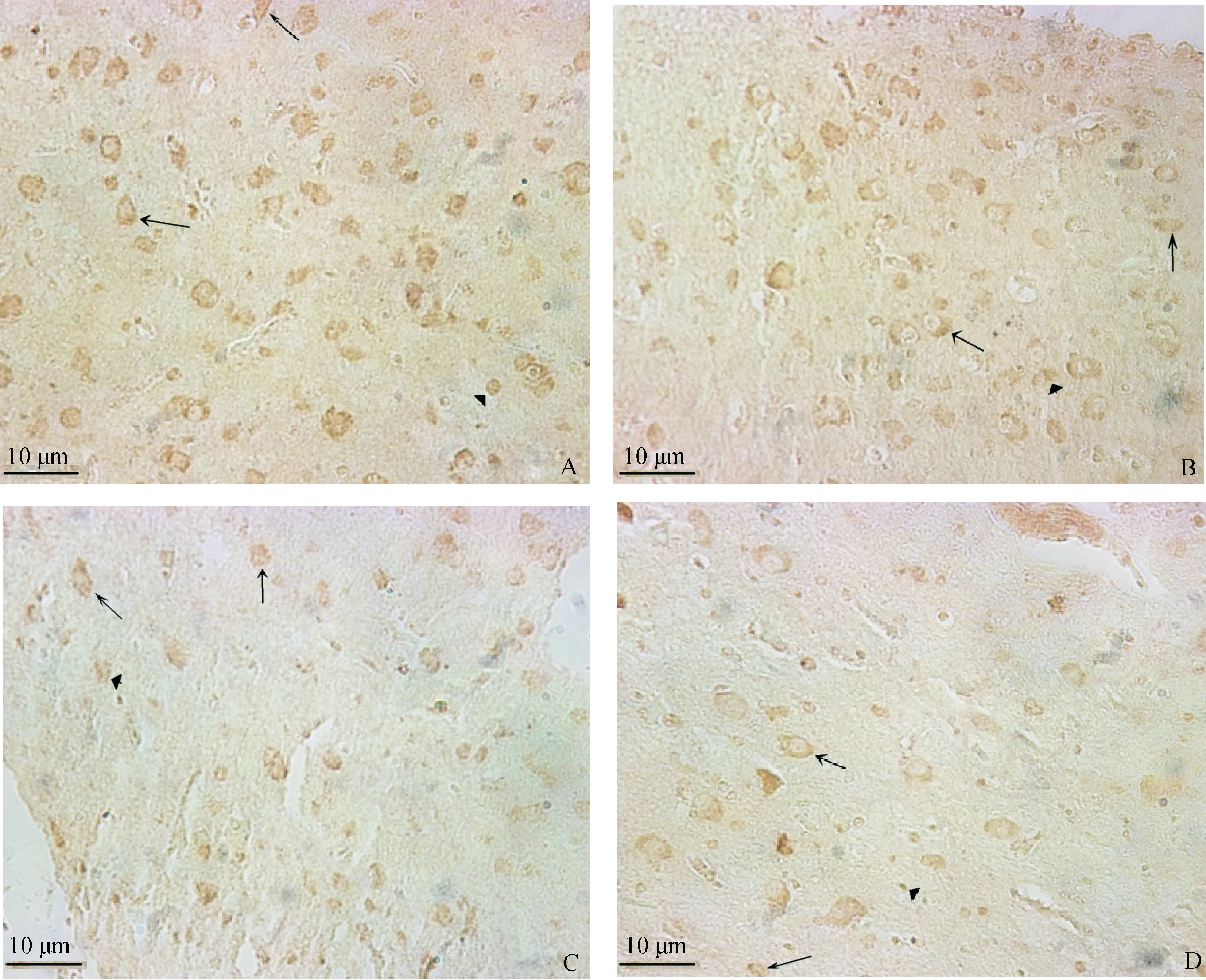
棕黄色或褐色为阳性信号。“→”为水平细胞;“”为锥体细胞;“▲”为星形胶质细胞。A.42 d对照组大脑皮层GABA;B.42 d 15 mg·kg-1试验组大脑皮层GABA;C.42 d 30 mg·kg-1试验组大脑皮层GABA;D.42 d 45 mg·kg-1试验组大脑皮层GABAThe postive signals of immunohistochemistry were tan and brown.“→”show the horizontal cells;“”show the cones;“▲”show the astrocytes.A.Control;B.15 mg·kg-1 experiment group;C.30 mg·kg-1 experiment group;D.45 mg·kg-1 experiment group图1 不同剂量钒对42日龄雏鸡大脑皮层GABA表达的影响Fig.1 Effect of vanadium on the expression of GABA in the cerebral cortex of 42-day-old chickens
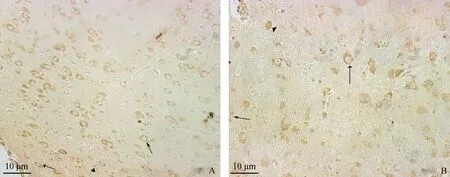
棕黄色或褐色为阳性信号。“→”为水平细胞;“”为锥体细胞;“▲”为星形胶质细胞A.14 d大脑皮层GABA;B.28 d大脑皮层GABA;42 d大脑皮层GABA的表达见图1DThe positive signals of immunohistochemistry were tan and brown “→”show the horizontal cells;“”show the cones;“▲”show the astrocytes.A.14 d;B.28 d;42 d′s figure is showed in the Fig.1 D图2 45 mg·kg-1钒添加量对不同日龄雏鸡大脑皮层GABA表达的影响Fig.2 Effects of GABA expression in the cerebral cortex of different day-old chickens with 45 mg·kg-1 vanadium
表245 mg·kg-1钒添加量对不同日龄雏鸡大脑皮层GABA表达的影响
Table 2Effects of GABA expression in the cerebral cortex of different day-old chickens with 45 mg·kg-1vanadium
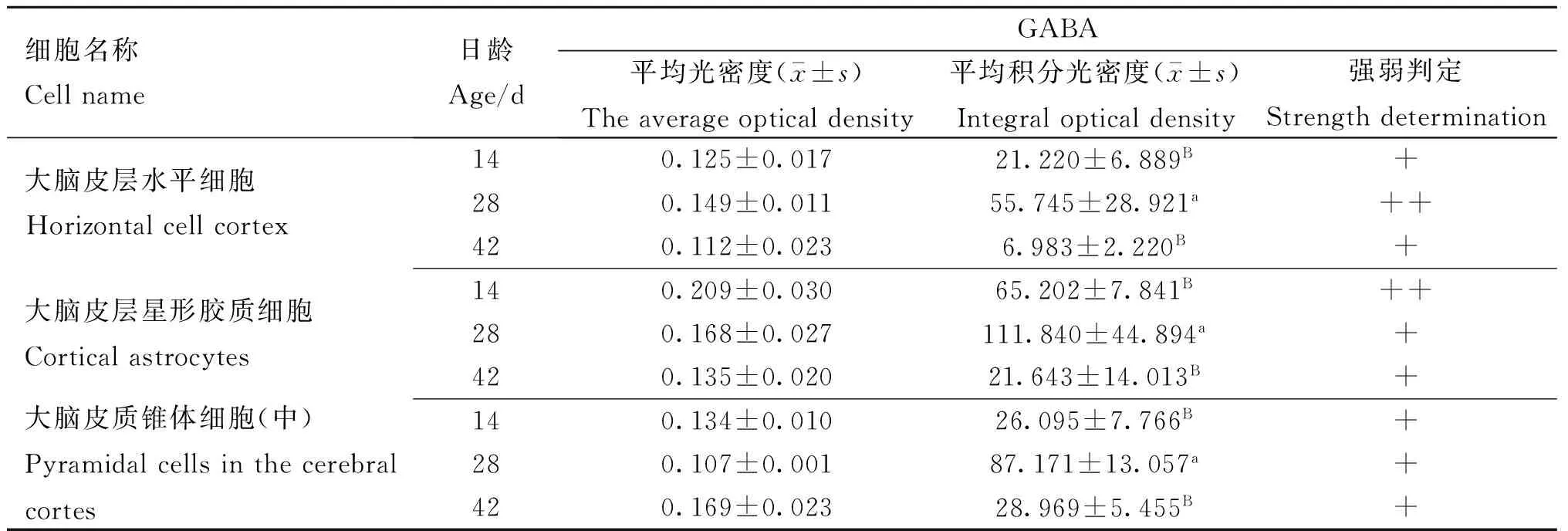
细胞名称Cellname日龄Age/dGABA平均光密度(x-±s)Theaverageopticaldensity平均积分光密度(x-±s)Integralopticaldensity强弱判定Strengthdetermination大脑皮层水平细胞Horizontalcellcortex140.125±0.01721.220±6.889B+280.149±0.01155.745±28.921a++420.112±0.0236.983±2.220B+大脑皮层星形胶质细胞Corticalastrocytes140.209±0.03065.202±7.841B++280.168±0.027111.840±44.894a+420.135±0.02021.643±14.013B+大脑皮质锥体细胞(中)Pyramidalcellsinthecerebralcortes140.134±0.01026.095±7.766B+280.107±0.00187.171±13.057a+420.169±0.02328.969±5.455B+
3 讨 论
3.1不同剂量钒对42日龄雏鸡大脑皮层GABA表达的影响
黄选洋等[10]指出,添加 15 mg·kg-1的钒可使雏鸡巨噬细胞数目增多,活性增强,淋巴细胞增殖,有助免疫器官的发育;30 mg·kg-1的钒可抑制雏鸡免疫功能,但也有研究指出其促进了免疫器官组织结构的发育,故30 mg·kg-1钒添加量对鸡免疫器官的影响尚有争议。
本课题组研究表明[22],15 mg·kg-1钒添加组雏鸡的消化、免疫等器官指数、组织学指标、抗氧化能力等均有正向升高;30 mg·kg-1钒添加量对大部分器官有正向影响,但个别器官表现组织学结构受损。
GABA是三羧酸循环中“GABA 环路”的产物,在体内也作为供能物质参与能量代谢[23]。资料表明[24],GABA具有增强黏膜免疫、促进消化吸收、调节神经—内分泌—免疫网络、减轻应激等作用。GABA可促进下丘脑分泌生长激素释放激素,作用垂体使生长激素分泌增加,影响胃肠道激素的分泌;可通过抑制交感-肾上腺系统,抑制去甲肾上腺素的分泌,促使自由基的产生量减少;抑制神经元的缺氧损伤,在神经元的发生、发育、可塑性等方面发挥着积极作用,使神经元处于“稳态”[14];此外,GABA对自身免疫性炎症有良好的抑制作用,原理是通过GABA干预CD4+(辅助性T细胞)和CD8+(细胞毒性T细胞)的数量及其比值发挥作用,通过抑制免疫反应,维持机体内环境的稳定[15-17]。
本试验结果显示,42 d雏鸡,15 mg·kg-1钒添加组的大脑皮层GABA表达量最高,其次为30 mg·kg-1钒添加组的,45 mg·kg-1钒添加组的高于对照组的。提示15 mg·kg-1钒添加组雏鸡大脑皮层高水平的GABA表达,可能与机体的供能、消化、免疫、抗氧化等的增强相关。30和45 mg·kg-1钒添加组GABA表达的上调,可能与机体抑制神经元的缺氧损伤,维持机体内稳态有关,其机制尚需进一步的研究。
3.245 mg·kg-1钒添加量对不同日龄雏鸡大脑皮层GABA表达的影响
研究表明[10],鸡日粮中添加高剂量钒时,添加初期其抗氧化酶的活性代偿性升高,但长期摄入高剂量的钒,则对机体造成氧化损伤。资料指出[24],小鼠在应激初期,GABA含量未升高,甚至下降,机体处于应激适应期;在应激后期,GABA积聚,以避免应激对机体造成过度伤害。阮怀珍等[25]指出,艾维因肉鸡胃蛋白酶随日龄的增长而逐渐增加,于28 d达到峰值,随后降低。
课题组研究表明[22],45 mg·kg-1钒添加量,在添加初期,部分器官表现较好的耐受性。本试验结果显示,45 mg·kg-1钒添加组,28 d时大脑皮层GABA表达量极显著高于14 d和42d的;14 d的三种细胞GABA表达量高于42 d。提示45 mg·kg-1钒添加量在添加时间为14 d时,大脑皮层GABA的较高表达可能与机体部分器官有较好的耐受性有关;而添加时间28 d时,可能因钒剂量的累积,以及28 d可能是雏鸡细胞分化的重要阶段,为维持机体内稳态的稳定,大脑皮层GABA的表达量达到最高值;钒添加时间为42 d时,大脑皮层GABA的表达略高于对照组,可能是为避免钒毒性累积对机体造成过度伤害。其具体机制仍需进一步的探讨。
4 结 论
15 mg·kg-1钒添加量与大脑皮层GABA表达呈正相关;45 mg·kg-1钒添加量,添加时间28日龄时,大脑皮层GABA表达呈拐点。提示日粮钒与雏鸡大脑皮层GABA的表达存在量效与时效关系,其调控作用可能与GABA的表达上调相关。
[1]CUI W,GUO H,CUI H.Vanadium toxicity in the thymic development[J].Oncotarget,2015,6(30):28661-28677.
[2]DENG Y,CUI H,PENG X,et al.Effects of high dietary fluoride on serum biochemistry and oxidative stress parameters in broiler chickens[J].Health,2014,06(14):1840-1848.
[3]LIU J,CUI H,LIU X,et al.Dietary high vanadium causes oxidative damage-induced renal and hepatic toxicity in broilers[J].BiolTraceElemRes,2012,145(2):189-200.
[4]CUI W,CUI H,PENG X,et al.Dietary vanadium induces lymphocyte apoptosis in the bursa of Fabricius of broilers[J].BiolTraceElemRes,2011,146(1):59-67.
[5]WANG K,CUI H,DENG Y,et al.Effect of dietary vanadium on the ileac T cells and contents of cytokines in broilers[J].BiolTraceElemRes,2011,147(1-3):113-119.
[6]DENG Y,CUI H,XI P,et al.Changes of IgA+cells and cytokines in the cecal tonsil of broilers fed on diets supplemented with vanadium[J].BiolTraceElemRes,2012,147(1-3):149-155.
[7]CUI W,CUI H,PENG X,et al.Dietary vanadium induces decrease in antioxidant enzyme activities and oxidative stress in the spleens of broilers[J].MedChem,2012,02(2):103-106.
[8]DENG Y,CUI H,PENG X.The relevance of cytokines for mucosal immunity induced by dietary vanadium in the intestine of broilers[J].MedChem,2012,02(2):051-056.
[9]CUI H.Effect of vanadium on TLR4 and TLR7 mRNA expression in the lymphoid organs of broilers[J/OL].MedChem,2012.[2016-06-23].http://www.omicsonline.org/2161-0444/2161-0444-2-103.pdf.
[10]黄选洋,王建萍,丁雪梅,等.钒对鸡健康的影响及其作用机理[J].动物营养报,2015,27(8):2335-2341.
HUANG X Y,WANG J P,DING X M,et al.Effects of vanadium on chicken health and its mechanism[J].JournalofAnimalNutrition,2015,27(8):2335-2341.(in Chinese)
[11]DENG Y,CUI H,PENG X,et al.Dietary vanadium induces oxidative stress in the intestine of broilers[J].BiolTraceElemRes,2012,145(1):52-58.
[12]WANG K,CUI H,DENG Y,et al.Effect of dietary vanadium on intestinal microbiota in broiler[J].BiolTraceElemRes,2012,149(2):212-218.
[13]戴四发.谷氨酰胺和γ-氨基丁酸对肉鸡抗热应激和肉品质的影响及机理探讨[D].南京:南京农业大学,2012.
DAI S F.Effects and mechanism of glutamine and gamma-aminobutyric acid supplementation on anti-stress and meat quality of broilers under heat stress[D].Nanjing:Nanjing Agricultural University,2012.(in Chinese)
[14]徐同锏.包被γ-氨基丁酸对奶牛热应激的防治效果及机理的研究[D].安徽:安徽农业大学,2013.
XU T J.Effects coated gamma-aminobutyric acid on alleviating heat stress of cows and investigation on the mechanism[D].Anhui:Anhui Agricultural University 2013.(in Chinese)
[15]张志荣.Gabarap在炎症反应过程中的功能研究[D].厦门:厦门大学,2014.
ZHANG Z R.The study of roles of Gabarap in inflammatory responses[D].Xiamen:Xiamen University,2014.(in Chinese)
[16]BHAT R,AXTELL R,MITRA A,et al.Inhibitory role for GABA in autoimmune inflammation[J].ProcNat1AcadSciUSA,2010,107(6):2580-2585.
[17]马玉华,王斌,孙进,等.γ-氨基丁酸对高脂膳食小鼠免疫功能的影响[J].免疫学杂志,2014,(7):599-603.
MA Y H,WANG B,SUN J,et al.The effect of GABA on immune function of mice fed with high fat diet[J].JournalofImmunology,2014,(7):599-603.(in Chinese)
[18]程肖蕊,周文霞,张永祥.中枢神经系统谷氨酸转运体的研究进展[J].生命科学,2009,(2):246-252.
CHENG X R,ZHOU W X,ZHANG Y X.Glutamate transporters in central nervous system[J].LifeScience,2009,(2):246-252.(in Chinese)
[19]杨银凤.家畜解剖学及组织胚胎学[M].北京:中国农业出版社,2011.
YANG Y F.Animal anatomy and embryology[M].Beijing:China agriculture press,2011.(in Chinese)
[20]曾洋,郑国颖,李明艳,等.氟和砷染毒对子代大鼠海马和大脑皮质组织谷氨酸与γ-氨基丁酸含量的影响[J].环境与健康杂志,2014,31(5):402-405.
ZENG Y,ZHENG G Y,LI M Y,et al.Effects of fluoride and arsenic exposure on contents of glutamate and gamma-aminobutyric acid in hippocampus and cortex in offspring rats[J].Journalofenvironmentandhealth,2014,31(5):402-405.(in Chinese)
[21]POGGIOLI R,ARLETTI R,BERTOLINI A,et al.Behavioral and developmental outcomes of prenatal and postnatal vanadium exposure in the rat[J].PharmacolRes,2001,43(4):341-347.
[22]邓苑馨.日粮钒添加水平对雏鸡肠道免疫功能和抗氧化功能影响的研究[D].雅安:四川农业大学,2012.
DENG Y X.The effects of dietary vanadium on intestinal immune function and antioxidant function in broilers[D].Yaan:Sichuan Agricultural University,2012.(in Chinese)
[23]史婧.γ-氨基丁酸、γ-氨基丁酸受体、γ-氨基丁酸受体基因在脑梗死中的作用[J].包头医学,2011,35(1):3-6.
SHI J.Role GABA,GABA receptors,GABA receptor gene in cerebral infarction[J].JournalofBaotouMedicine,2011,35(1):3-6.(in Chinese)
[24]李慧.γ-氨基丁酸对高温条件下蛋鸡产蛋性能及抗热应激的影响[D].杭州:浙江大学,2010.
LI H.Effects of gamma-aminobutyric acid on laying performance and heat stress resistance capability in hens under high temperature in summer[D].Hangzhou:Zhejiang University,2010.(in Chinese)
[25]阮怀珍,杨萍,张吉强,等.医学神经生物学课程体系的建设与实践[J].基础医学教育,2008,10(4):404-406.
RUAN H Z,YANG Z,HU Z A,et al.Construction and practice in the curriculum system of medical nerve biology[J].BasicMedicalEducation,2008,10(4):404-406.(in Chinese)
(编辑白永平)
Effect of Vanadiumon on the Expression of GABA in the Cerebral Cortex in Broilers
TANG Li,XIN Dan-ning,TIAN Chun-yang,CUI Heng-min
(CollegeofVeterinaryMedicine,SichuanAgriculturalUniversity,Chengdu611130,China)
To study the effect of vanadium on the chick cerebral cortex γ-aminobutyric acid (GABA) expression,280 1-day-old Avian broilers were chosen and randomly divided into four groups,each group divided into seven sample groups,each sample group have 10 broilers,and fed control diet and diet added vanadium (low dose group:15 mg·kg-1;dose group:30 mg·kg-1;high dose group:45 mg·kg-1) for 6 weeks using the immunohistochemical method to detect the expression of chick cerebral cortex GABA Variety.The results showed that the expression level of the cerebral cortex GABA 42 d chicks,15 mg·kg-1of vanadium-added group was the highest (P<0.01 orP<0.05),followed by 30 mg·kg-1of vanadium added group (P<0.01 orP<0.05),45 mg·kg-1of vanadium-added group than the control group (P>0.05);45 mg·kg-1of vanadium-added group,when at 28 d the cerebral cortex expression levels were significantly (P<0.01) higher than the 14 and 42 d;when at the 14 cortex horizontal cells and astrocytes of GABA expression were higher than 42 d (P>0.05).In summary,it showed that the dietary vanadium and the expression of chick’s cortex GABA exist in a dose and time-effect relationship,suggesting that the regulation of expression may be related to up-regulation of GABA.
vanadium;cerebral cortex;GABA;immunohistochemical method;broilers
10.11843/j.issn.0366-6964.2016.09.024
2016-04-12
教育部博士后基金项目(04310611)
唐丽(1979-),女,陕西洛南人,副教授,博士,主要从事禽类神经内分泌系统的研究,E-mail: tangyimingtt@163.com;Tel:028-86291348。注:唐丽和辛丹宁贡献相同,为共同第一作者。
崔恒敏,教授,E-mail:cuihengmin2008@sina.com
S852.3
A
0366-6964(2016)09-1940-07
