欧美杨与藏川杨杂交子代苗期性状QTLs定位分析
2016-10-26安新民
李 娟 郭 斌,2 安新民
(1. 北京林业大学林木育种国家工程实验室,北京 100083;2. 山西省林业科学研究院,山西 太原 030012)
欧美杨与藏川杨杂交子代苗期性状QTLs定位分析
李娟1郭斌1,2安新民1
(1. 北京林业大学林木育种国家工程实验室,北京 100083;2. 山西省林业科学研究院,山西 太原 030012)
以欧美杨I-108和藏川杨为亲本杂交得到427个子代为试材,对其进行苗期性状遗传图谱分析,并构建了1张总长度969.1 cM,标记间平均距离25.14 cM的杂交杨树遗传图谱。结果表明:该图谱包含6个连锁群,连锁群上标记数量37个,用Genemap软件绘制标记连锁图谱;检测到株高 (hs)、单叶光合速率 (Pn)、胞间CO2浓度 (Ci)、蒸腾速率 (Tr) 4个表型指标符合正态分布。利用MAPMAKER软件的区间作图法进行QTL定位和效应估计, 共检测到8个与苗期生长和光合性状相关的QTL,包括3个与hs相关的QTL,贡献率分别为16.4%、20.1%和17.8%;2个与Pn相关的QTL,贡献率分别为10.2%和13.7%;2个与Ci相关的QTL,贡献率分别为6.7%和8.3%;1个与Tr相关的QTL,贡献率9.7%。
藏川杨;欧美杨;杂交;遗传图谱;QTL定位;苗期性状
林木遗传改良的长期实践证明,基因资源及能否科学有效地选择与利用这些资源对于育种至关重要[1]。在我国分布有丰富的乡土杨树资源,发掘、保护、利用这些极其重要的乡土杨树资源,是我国杨树遗传育种研究值得高度重视的方向。作为第1个被测序的木本植物,杨树 (Populusspp.) 是研究木本植物生物学和遗传学的模式树种[2],杨树的遗传图谱构建工作进展迅速,已发表的遗传图谱超过13项,涉及白杨、青杨、黑杨三大派别,共14个种或杂种[3]。早在1993年,Liu等以美洲黑杨 (P.deltoides) 5个全同胞家系为作图群体,采用54个RELP和3个等位酶标记构建了杨树第1张遗传连锁图谱[4]。在我国,苏晓华等以美洲黑杨×青杨 (P.cathayana) F2代为作图群体,采用RAPD技术构建了我国杨树第1张遗传连锁图谱,为开展杨树数量性状QTL研究奠定了基础[5]。之后张德强采用AFLP技术构建了乡土杨树毛白杨 (P.tomentosa) 的遗传连锁图谱,为毛白杨遗传改良奠定了理论基础[6]。尽管杨树遗传图谱构建工作已取得显著进展,但在我国尚有不少乡土杨树资源未被开发。
藏川杨 (P.szechuanicavar.tibetica) 是一种位于青藏高原的青杨组乡土树种,也是我国特有的杨树,是西南地区分布海拔高的杨树树种之一,其分布地区环境多变,能够很好地适应高原地区环境,具有抗低温、低氧等抗性[7]。然而迄今为止,利用藏川杨作为亲本进行杂交育种鲜有报道。本研究以藏川杨为亲本与欧美杨 (P.euramericana) 杂交获得生长量大、生长快速、具有较强抗逆性的杂交子代508株,并以保留的427株为作图群体,采用SSR标记构建了遗传连锁图谱,并对生长、光合等性状进行了QTL分析。研究结果为进一步开发利用藏川杨基因资源奠定了工作基础。
1 材料与方法
1.1试验材料
欧美杨I-108是欧洲黑杨 (P.nigra) 和美洲黑杨的杂交品种,林木良种编号为国R-SC-PE-029-2002[8],具有显著的速生特性,本试验中作为母本。藏川杨西南地区在低纬度高海拔地区广泛分布的乡土树种,具有抗寒的特性,本试验中作为父本。通过人工杂交,得到508个杂交子代单株。
1.2研究方法
1.2.1杂交群体构建
在平均温度为20~25 ℃的温室下,对采集的藏
川杨雄株花枝进行水培,收集花粉并干燥于4 ℃储藏。随后对雌株花枝进行水培,待雌花序柱头发亮时进行人工授粉3次。共得到杂交子代508株,由于后期管理导致部分杂交成苗数损失,最终选用427个子代为测定群体。
1.2.2DNA提取与浓度检测
分别摘取亲本和杂交子代植株顶端幼嫩叶片1~2片,用改良的CTAB提取DNA[9]。用紫外分光光度计 (Pharmcia, MLTROSPEC Ⅲ) 测定DNA浓度。采用1.0%琼脂糖凝胶电泳确定DNA的质量,用凝胶成像系统观测电泳结果。
1.2.3SSR分子标记分析
使用毛细管电泳技术代替传统的聚丙烯酰胺凝胶电泳技术,毛细管电泳技术采用不同颜色的荧光染料标记SSR引物,随后进行毛细管电泳,通过GeneMarker软件读取、分析数据。采用70对4种不同颜色的带TP-M13 (tailed primer-M13) 对SSR引物进行标记。试验中的杨树引物来自北京林业大学计算生物学中心,选取4个子代,筛选了300对SSR引物。
1.2.4苗期生长性状与光合特性的测定
杂交试验于北京林业大学温室进行,日平均温度为20~25 ℃,平均湿度为50%~70%。对选取的427个子代进行株高测定;采用美国Lico-6400光合作用分析系统测定叶片净光合速率 (Pn)、胞间CO2浓度 (Ci)、蒸腾速率 (Tr) 等指标。设定光照强度为1 400 μmol/(m2·s),每天重复3次,连续测定3 d。对选取的无性系子代进行同种无性系不同叶片Pn的测定,确定测定光合作用指标时的叶片最佳位置。采用SPSS 16.0软件对杂交子代株高及光合生理指标进行差异显著性分析。
1.2.5构建遗传图谱及定位QTL
遗传距离用Mapmaker 3.0软件计算以确定染色体上的标记顺序。以QTL存在和不存在该位置的2种可能性之比,将比值取以10为底的对数为LOD值,当LOD ≥ 3.0,重组率 < 0.5时,根据SSR标记构建每条染色体的框架,用Kosambi函数将标记之间的重组交换率换算为遗传图距单位 (cM)。依据标记顺序、标记间的距离和连锁群,用Genemap软件绘制标记连锁图谱。采用MAPMAKER/QTL 1.1软件进行QTL定位分析,QTL的基因作用方式按照Stuber等来执行[10-11]。
2 结果与分析
2.1杨树株高表型性状及差异分析
作为重要的数量性状,株高在玉米 (Zeamays)、大麦 (Hordeumvulgare)、大豆 (Glycinemax) 等农作物中已进行了QTL定位分析,并且其也是影响木材材积的重要因素[12-14]。本试验采用SPSS 16.0
软件对杂交子代株高及光合生理指标进行差异显
著性分析。子代株高 (hs) 与单叶光合速率 (Pn) 相关系数为0.368,与蒸腾速率 (Tr) 相关系数为0.359,有极显著相关性,株高与胞间CO2浓度 (Ci) 相关系数为0.172,无显著相关性。将子代的hs、Pn、Ci、Tr等特征指标进行频率分布分析,均呈偏度小于2的单峰分布 (图1所示),其基本符合正态分布,可用于QTL定位分析。

图1杂交子代表型性状频率分布分析
Fig.1Frequency distribution analysis of phenotypes variation in hybrid progeny
2.2SSR引物多态性分析及遗传图谱构建
根据已有的SSR反应体系,从300对引物通过4个子代筛选出多态性好的70对引物,然后用这些引物进行遗传图谱的构建,共产生了80个SSR标记。用MAPMAKER 3.0软件对产生的标记进行两点连锁分析,得到连锁群的总数量以及每个连锁群的最佳顺序。共得到6个标记数目大于4的大连锁群,三联体8个和连锁对5个。构建的遗传图谱框架图的总图距为751.6 cM,遗传图谱的总长度为969.1 cM,平均标记数为6.2个,平均间距为25.14 cM,每个连锁群上的标记数为4~37个 (表1)。
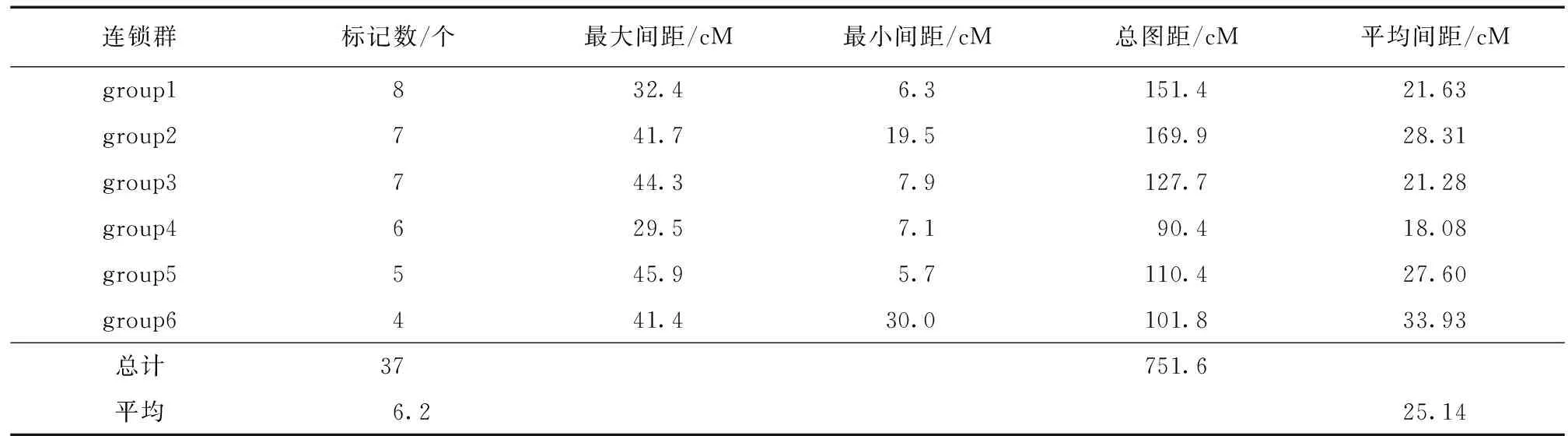
表1 欧美杨 × 藏川杨遗传图谱标记

图2欧美杨 × 藏川杨遗传连锁图谱
Fig.2The genetic linkage map ofP.euramericana×P.szechuanicavar.tibetica
2.3株高、光合速率、胞间CO2浓度及蒸腾速率的QTL分析
根据所作图谱和表型数据,利用MAPMAKER/
QTL 1.1软件对杂交子代苗期性状进行QTL分析,检测到与株高、Pn、Ci、Tr有关的QTL共8个 (表2)。共检测到3个与株高有关的QTLs,前两者分别位于group1连锁群上6.3 cM和32.4 cM处,总贡献率为36.5%。第3个与株高有关的QTL位于group4连锁群上20.3 cM处,贡献率为17.8%。检测到2个与Pn有关的QTLs,分别位于group2连锁群24.3 cM处、group3连锁群16.7处,贡献率分别为10.2%、13.7%。检测到2个与Ci有关的QTLs,分别位于group1连锁群12.7 cM处、group5连锁群5.7 cM处,贡献率分别为6.7%、8.3%。检测到一个与Tr有关的QTL,位于group1连锁群29.8 cM处,贡献率为9.7%。
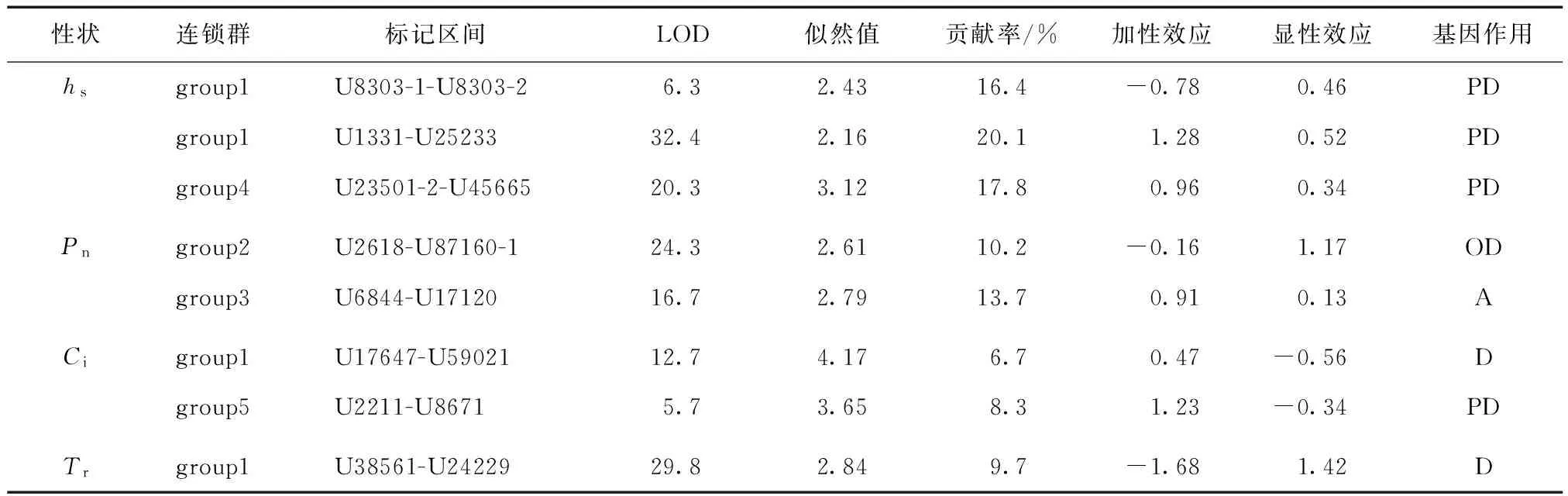
表2 欧美杨 × 藏川杨杂交子代的QTLs分析结果
注: A为当显性势d/a=0~0.20时为加性方式;PD为当显性势d/a=0.21~0.80时为部分显性方式;D为当显性势d/a=0.81~1.2时为显性方式;OD为当显性势d/a>1.20时为超显性方式。
3 结论与讨论
我国杨树资源十分丰富,乡土杨树资源的遗传图谱构建工作对于开发利用我国杨树资源十分重要。张德强等利用AFLP标记技术构建了我国乡土树种毛白杨的遗传连锁图谱,包括218个标记,全场2 683 cM,覆盖毛白杨全基因组87%[6]。李博等利用毛白杨与毛新杨 (P.tomentosa×P.bolleana) 的F1代回交群体,构建了根萌苗和木质部的2张转录组图谱,并对生长、生理和木材品质的28个数量性状进行了QTL分析[3]。选择遗传背景差别大的杨树亲本进行杂交,构建株系间遗传差异明显的作图群体是决定遗传连锁图谱中标记选择和标记密度的重要基础[15]。当遗传图谱分子标记平均间距在10~20 cM 就可以进行QTL初级定位[16]。藏川杨是我国西南地区低纬度高海拔地区广泛分布的乡土树种,具有抗寒、干性通直的优良特性。欧美杨I-108具有速生、抗旱等优良特性。本试验以欧美杨 × 藏川杨杂交子代427个单株为作图群体,采用SSR标记,构建的杨树遗传图谱的长度为969.1 cM,包含了80个SSR标记的6个大连锁群。根据Hulbert等提出的公式进行估算,基因组长度约为1 155 cM。本研究中构建的图谱总长度覆盖基因组约为83.9%,框架图覆盖基因组约为65.1%。导致本试验覆盖基因组范围较小的主要原因可能是采用的SSR标记数量较少,以及标记类型单一,这也是造成未连锁标记比例较大 (11.3%) 的主要原因。下一步工作可通过增加SSR标记,结合AFLP标记乃至组学数据增加遗传图谱的饱和度。
国内外专家对许多农作物的株高进行了QTL定位分析,如大豆 (Glycinemax)、玉米 (Glycinemax)[17-18]等,表明株高是影响植物光合等生理过程的重要因素。株高代表了树木的材积及生长过程,因此找出与株高相关的QTL至关重要,可为提高育种效率奠定基础[7]。光合作用是绿色植物能量和有机物质的来源,是植物成长的决定性因素之一。为了更好地对杨树遗传的改良,需对其光合指标进行QTL定位。本研究对株高、单叶光合速率、蒸腾速率和胞间CO2浓度等性状进行QTL定位。杂交子代的平均株高为644.9 mm,极值为22~1 490 mm,变异系数为0.43;平均单叶光合速率为19.37 μmol/(m2·s),极值为9.54~28.92 μmol/(m2·s),变异系数为0.16;胞间CO2浓度平均值为294.04 μmol/mol,极值为155.87~428.62 μmol/mol,变异系数为0.15;平均蒸腾速率为3.8 mol/(m2·s),极值为0.68~6.96 mol/(m2·s),变异系数为0.28。数据表明株高的变异系数最大,胞间CO2浓度的最小。对4个指标进行频率分布分析,均符合正态分布,可用于QTL定位。本试验采用区间作图法,用MAPMAKER软件,其依赖于完整的遗传图谱LOD值。LOD值与作图群体的种类和大小密切相关,LOD值通常与作图群体大小成正比[17]。但由于所用的标记数少,且集中在几条染色体,导致构建的连锁群远少于杨树的染色体条数,综合考虑,LOD值取为2[18]。试验群体为427,所定位的8个QTL中只有1个是超显性方式,其余分别为加性方式、显性方式和部分显性方式。研究表明,超显性方式的比重与群体的大小成反比,群体的大小决定了QTL定位的可靠性和准确性。因此,在育种中要重视QTL的超显性效应。图谱的标记密度与QTL定位也关系密切,标记密度要适中。除此之外,影响QTL的因素还包括控制该性状的QTL环境影响、遗传特性和实验误差等[19-20]。
本研究利用我国特有的乡土杨树藏川杨为父本,以欧美杨I-108为母本构建了作图群体,获得一批新的种质资源,利用该群体初步构建了遗传图谱,并对苗期生长、光合等相关性状进行了QTL分析,为进一步加密该遗传图谱和利用乡土藏川杨基因资源开展分子辅助育种工作奠定了基础。
[1]王明庥, 黄敏仁, 邬荣领, 等. 美洲黑杨 × X小叶杨杂交育种研究[M]. 北京: 科学技术文献出版社, 1991.
[2]Tuskan G A, Difazio S, Jansson S, et al. The genome of black cottonwood,Populustrichocarpa(Torr. & Gray) [J]. Science, 2006, 313(5793): 1596-1604.
[3]李博. 毛白杨与毛新杨转录组图谱构建及若干性状的遗传学联合分析[D]. 北京: 北京林业大学, 2009.
[4]Liu Z, Furnier G R. Comparison of allozyme, RFLP, and RAPD markers for revealing genetic variation within and between trembling aspen and bigtooth aspen[J]. Theoretical & Applied Genetics, 1993, 87(1/2): 97-
105.
[5]苏晓华, 张绮纹, 郑先武, 等. 美洲黑杨 (PopulusdeltoidesMarsh)×青杨 (P.cathayanaRehd.) 分子连锁图谱的构建[J]. 林业科学, 1998, 34(6): 29-37.
[6]Zhang D Q, Zhang Z Y, Yang K, et al. Genetic mapping in (Populustomentosa×Populusbolleana) andP.tomentosaCarr. using AFLP markers[J]. Tag. theoretical & Applied Genetics. theoretische Und Angewandte Genetik, 2004, 108(4): 657-662.
[7]薄文浩. 藏川杨遗传多样性及杂交子代遗传变异研究[D]. 北京: 北京林业大学, 2012.
[8]王维正, 刘红. 林木良种指南[M]. 北京: 中国林业出版社, 2003.
[9]宋国立, 崔荣霞, 王坤波, 等. 改良CTAB法快速提取棉花DNA[J]. 棉花学报, 1998(5): 273-275.
[10]Lander E S, Green P, Abrahamson J, et al. MAPMAKER: an interactive computer package for constructing primary genetic linkage map and natural populations[J]. Genomics, 1987, 1(2): 174-181.
[11]Stuber C W, Edwards M D, Wendel J E. Molecular marker-facilitated investigations of quantitative traits loci in maize. Factors influencing yield and its component traits[J]. Crop Science, 1987, 27: 639-648.
[12]万昆, 邱丽娟. 大豆遗传图谱的加密及重要农艺性状的QTL定位[C]// 2008中国作物学会学术年会论文摘要集, 2008.
[13]Thomas W T B, Powell W, Waugh R, et al. Detection of quantita-tive traits loci for agronomic, yield, grain and disease characters in spring barley (HordeumvulgareL) [J]. Theoretical and Applied Genetics, 1995, 91: 1037-1047.
[14]杨晓军, 路明, 张世煌, 等. 玉米株高和穗位高的QTL定位[J]. 遗传, 2008, 30(11): 1477-1486.
[15]Röder M S, Korzun V, Wendehake K, et al. A microsatellite map of wheat[J]. Genetics, 1998, 149: 2007-2023.
[16]方宣钧, 吴为人, 唐纪良. 作物DNA标记辅助育种[M]. 北京: 科学出版社, 2002.
[17]张君. 玉米株型及产量相关性状QTL定位与分析[D]. 郑州: 河南农业大学, 2010.
[18]王毅, 姚骥, 张征锋, 等. 基于玉米综合QTL图谱的比较分析及株高QTL的统合分析[J]. 科学通报, 2006, 51(15): 1776-1786.
[19]张福敏. 响叶杨 × 银白杨遗传图谱加密与相关QTL定位[D]. 南京: 南京林业大学, 2011.
[20]伊海法. 棉花产量和纤维品质性状QTL定位[D]. 泰安: 山东农业大学, 2014.
(责任编辑张坤)
Mapping of QTLs in Hybrid Progeny ofPopuluseuramericanaandPopulusszechuanicavar.tibetica
Li Juan1, Guo Bin1,2, An Xinmin1
(1. National Engineering Laboratory for Tree Breeding, Beijing Forestry University, Beijing 100083, China;2. Shanxi Academy of Forestry Sciences, Taiyuan Shanxi 030012, China)
A population including 427 germplasm resources, crossed by female parentPopuluseuramericanaI-108 and male parentPopulusszechuanicavar.tibeticawere used to construct the genetic linkage map in seedling stage. The map contains 37 SSR makers and 6 linkage groups spanning 969.1 cM with an average of 25.14 cM between markers. Genemap software was used to draw the marker linkage map. The four phenotype indicators of height (hs), single leaf photosynthetic rate (Pn), intercellular CO2concentration (Ci), transpiration rate (Tr) are in line with normal distribution. The interval mapping method of software package MAPMAKER were used to map andanalyze QTLs. 8 QTLs were detected for seedling growth and photosynthetic trait. 3 QTLs were detected for seedling height which explained 16.4%, 20.1% and 17.8% of the phenotypic variation. 2 QTLs were detected for photosynthetic rate which explained 10.2% and 13.7% of the phenotypic variation. 2 QTLs were detected for Intercellular CO2concentration which explained 6.7% and 8.3% of the phenotypic variation. 1 QTL was detected for transpiration rate which explained 9.7% of the phenotypic variation.
Populusszechuanicavar.tibetica,Populuseuramericana, artificial hybridization, genetic map, QTL locationing, seedling trait

10. 11929/j. issn. 2095-1914. 2016. 05. 002
2016-01-27
国家 “十二五” 科技支撑项目 (2012BAD01B0302) 资助。
安新民 (1968—),男,博士,教授,博士生导师。研究方向:林木功能基因组学与分子育种。Email: anxinmin@bjfu.edu.cn。
S722.3
A
2095-1914(2016)05-0010-06
第1作者:李娟 (1986—),女,硕士。研究方向:林木功能基因组学与分子育种。Email: lijuan3315@126.com。
共同第1作者:郭斌 (1986—),男,硕士。研究方向:杨树杂交育种。Email: guobin531188058@163.com。
