灯盏花发根培养条件筛选研究
2016-10-26陈瑞兵
黄 鑫 季 倩 陈瑞兵 何 德 张 磊
(1. 西南林业大学生命科学学院,云南 昆明 650224;2. 第二军医大学药学院,上海 200433)
灯盏花发根培养条件筛选研究
黄鑫1,2季倩2陈瑞兵2何德1张磊1,2
(1. 西南林业大学生命科学学院,云南 昆明 650224;2. 第二军医大学药学院,上海 200433)
为建立灯盏花的发根培养体系,用C58C1发根农杆菌空菌株和携带目的基因的菌株转化灯盏花叶片获得发根,测定发根的生长曲线,比较不同因素对发根生长量的影响。结果表明:利用发根农杆菌C58C1菌株成功从灯盏花中诱导出了发根,并建立了灯盏花发根最优的培养体系,其诱导的最佳农杆菌菌液浓度为OD600=0.6,最佳浸染时间为10 min,最佳共培养时间为2 d,在此条件下诱导率高达60%。经PCR鉴定证明Ri质粒的T-DNA已成功转化灯盏花的发根。
发根农杆菌;诱导率;遗传转化体系;灯盏花;培养基
灯盏花 (Erigeronbreviscapus) 又名灯盏细辛、短葶飞蓬,为菊科飞蓬属多年生草本药用植物,全草入药。其主要有效成分为灯盏乙素,又名野黄芩苷[1]。灯盏乙素具有舒张血管、改善微循环、抑制血小板及红细胞聚集,治疗冠心病、心绞痛、肾衰和高血脂症等疾病,具多种作用[2-4]。但是作为云南最有发展前景的药物之一,却面临着因滥采滥挖及种质退化导致资源紧缺的严峻挑战[5-7]。因此,从处于代谢节点处的关键基因着手,改善灯盏花的种质,提高灯盏花素的含量成为亟待解决的问题。
目前,灯盏花种质资源的改善主要依靠人工栽培驯化,驯化改善的灯盏花品种中有效成分灯盏乙素虽然可达1.053%~2.628%[8],但仍有很大提升空间,而且还面临着种质很容易退化的严重问题。植物细胞的遗传转化是植物基因工程的一个关键环节。以发根农杆菌 (Agrobacteriumrhizogenes) Ri质粒为载体可将植物次生代谢物合成关键酶基因整合到植物基因组中并表达,从而达到稳定改良植物性状、提高次生代谢物含量的目的[9]。发根体系具有操作过程简便,遗传稳定性强,生长快、易于培养获得转基因材料等优势,发展潜力巨大。
植物次生代谢工程技术的应用已在多种植物中取得成果,如将矮牵牛 (Petuniahybrida) 中黄酮代谢途径中的查尔酮异构酶基因CHI导入西红柿中并过量表达,使转基因西红柿中果皮黄酮醇含量提高了78倍[10]。过量表达莨菪类生物碱合成的关键酶基因PMT和H6H,成功将莨菪转基因发根中东莨菪碱的含量提高了9倍[11]。过表达丹参JAs合成途径中的关键酶基因AOC、AOS和JMT,显著提高了丹参发根中丹参酮ⅡA的含量[12]。过量表达关键酶基因PLR成功提高了菘蓝发根中落叶松脂素的含量[13]等。而对于灯盏花,目前尚无建立遗传转化体系的报道。
本研究利用发根农杆菌Ri质粒成功地从灯盏花叶片诱导出发根,初步建立了最适灯盏花发根培养的遗传转化培养体系,为进一步利用遗传代谢工程手段调节灯盏花次生代谢物含量提供了可行方法,以期用此方法部分替代野生资源,减轻滥采滥挖现状及解决种质退化问题。
1 材料与方法
1.1材料与试剂
1.1.1材料来源
供试灯盏花种子由西南林业大学刘江华教授提供,农杆菌C58C1由第二军医大学药学院保存。
1.1.2试剂
试验用到的HgCl2,MS、B5、YEB等培养基,蔗糖,琼脂,琼脂糖凝胶,头孢、潮霉素、利福平等抗生素,各试剂均为国产分析纯试剂。rolB、rolC基因引物由苏州金唯智生物公司合成。PCR反应的rTaqDNA 聚合酶Mix购自Takara生物公司。
1.2试验方法
1.2.1植物材料处理
灯盏花种子用0.1%升汞浸泡10 min消毒,无菌水冲洗3~5次,灭菌吸水纸吸除残留水分。将灭菌种子播种到MS固体空白培养基上,置于4 000 lx、16 h日光照,25 ℃条件下培养。种子3~5 d后开始发芽,大约15 d后长出无菌实生苗,45~60 d长成可供剪取叶盘的灯盏花无菌苗。
1.2.2发根农杆菌菌株活化
取-80 ℃冻存的C58C1菌液,于含40 mg/L利福平 (Rif) 的YEB平板上划线,28 ℃恒温暗培养2 d。挑取单菌落于1 mL YEB (含40 mg/L Rif) 液体培养基中活化培养12~16 h。取活化的C58C1菌液200 μL加入到30 mL YEB (40 mg/L Rif) 液体培养基中28 ℃,200 r/min振荡培养至菌液OD600分别为0.4、0.6、0.8。常温下,5 000 r/min离心10 min,去上清,分别加30 mL B5、1/2B5、1/2MS、MS (均含100 μmol/L乙酰丁香酮) 液体培养基重悬菌体,28 ℃,100 r/min振荡孵育30 min,用以浸染叶盘。最后统计分析不同浓度的菌液对转化效率的影响。
1.2.3叶盘法转化
将灯盏花无菌苗叶片剪成约0.5 cm2大小的叶盘,分别置于B5、1/2B5、1/2MS、MS固体空白培养基上25 ℃恒温暗培养2 d (预培养)。将部分叶盘不作农杆菌浸染处理,以作对照。其余预培养后的叶盘分别放入制备好的OD600为0.4、0.6、0.8的3种浓度的农杆菌菌液中,分别浸染5、10、15 min,捞出后吸干残留菌液。再分别放回预培养时的B5、1/2B5、1/2MS、MS固体培养基,25 ℃,分别无光恒温共培养0、1、2、3 d。之后将共培养的叶盘对应转到含500 mg/L头孢噻污钠 (Cef) 的B5、1/2B5、1/2MS、MS固体培养基,25 ℃无光恒温除菌培养10~15 d后,一般可看到叶盘边缘伤口处有发根长出,待发根长至2~3 cm后将其剪下,分别对应在新的含500 mg/L Cef及10 mg/L潮霉素B (Hyb) 的B5、1/2B5、1/2MS、MS固体培养基上进行除菌筛选培养,每15~20 d更换1次培养基。每更换1次培养基,培养基其他成分不变,抗生素Cef浓度按500 mg/L → 300 mg/L → 100 mg/L → 0 mg/L阶梯性降低,直至除菌完全。
最后统计比较不同菌液浓度 (处理A—OD600=0.4;B—处理OD600=0.6;C—处理OD600=0.8)、不同浸染时间 (处理a—5 min;处理b—10 min;处理c—15 min) 和不同共培养时间 (处理1—0 d;处理2—1 d;处理3—2 d;处理4—3 d) 对叶盘发根诱导率、叶盘死亡率及染菌率等的影响,从而得出灯盏花发根最佳诱导培养条件。
1.2.4发根鉴定
取在除菌筛选培养基上仍生长良好的发根约100 mg,采用CTAB法提取总DNA,以总DNA为样品模板,同时以C58C1菌为阳性对照,以非转化根DNA为阴性对照,PCR鉴定毛状根总DNA中是否含发根农杆菌C58C1 Ri质粒的rolB和rolC基因:1) primers forrolB:rolB-F: 5′-CGA GGG GAT CCG ATT TGC TT-3′;rolB-R: 5′-GAC GCC CTC CTC GCC TTC CT-3′。2) primers forrolC:rolC-F: 5′-TCG CCA TGC CTC ACC AAC TCA C-3′;rolC-R: 5′-CCT TGA TCG AGC CGG GTG AGA A-3′。3) 反应体系 (20 μL):ddH2O 7 μL, Template DNA 1 μL, Forward Primer (10 μmol/L) 1 μL,Reverse Primer (10 μmol/L) 1 μL, 2 × rTaq PCR SuperMix 10 μL。4) PCR反应程序:94 ℃ 5 min;94 ℃ 30 s,55 ℃ 30 s,72 ℃ 40 s,35个循环;72 ℃ 5 min。凝胶成像系统下观察DNA条带。
1.2.5液体振荡扩大培养
选取除菌完全且鉴定为阳性的发根,剪取约300 mg,分别放入装有200 mL B5、1/2B5、1/2MS、MS液体培养基的三角瓶,置于摇床上,25 ℃避光,100 r/min振荡培养,每周更换1次培养基并称质量,比较不同培养基对发根生长速率的影响。
2 结果与分析
结果显示,对照组未经农杆菌浸染诱导处理的叶盘也有极少部分长出了不定根。灯盏花发根诱导流程见图1。经农杆菌浸染处理的试验组叶盘,因培养条件不同,表现出了不同的诱导率。具体诱导率见表1。
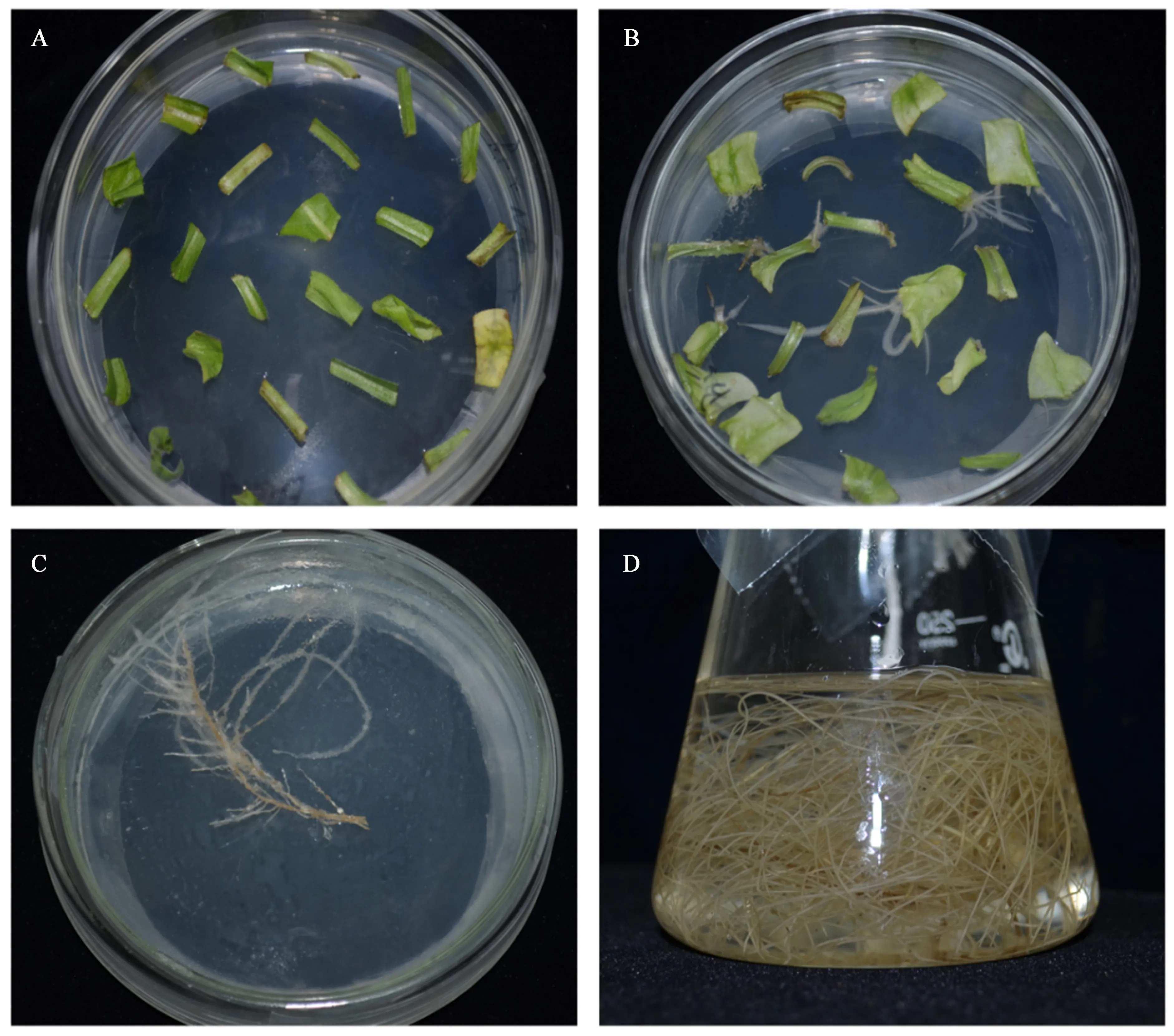
A. 叶盘预培养; B. 诱导; C. 除菌筛选培养; D. 液培
图1灯盏花发根诱导流程

Fig.1 The flow chart of hairy root induction of Erigeron breviscapus
注:为方便统计,表中所述长出发根的叶盘数是指凡是叶盘边缘长出了根的叶盘的数量,这些根不一定是发根,因此,诱导率也并不一定是阳性诱导率;死亡和染菌严重的叶盘数指培养过程中黄化死亡未能长出发根的,及虽长出了发根但染菌严重,无法脱菌完全的叶盘总数。
2.1发根的鉴定
按上述发根鉴定方法对脱菌完全的发根提取DNA进行PCR鉴定,其中对照组绝大部分在除菌筛选培养时已黄化死亡,少数存活的不定根PCR鉴定也均不含rolB、rolC基因,因此不是发根。试验组则大部分是有rolB、rolC基因的阳性发根。PCR鉴定结果如图2。阳性发根则于液体培养基中扩大培养。

M: T-2000 DNA Maker; N: 阴性对照; P: 阳性对照; 1~10: 样品
图2发根中rolB、rolC基因PCR鉴定结果
Fig.2The PCR detection ofrolBandrolCin transformed hairy root
2.2菌液浓度对发根诱导率的影响
在农杆菌介导的遗传转化中,菌液浓度是否适当是转化成功与否的关键问题之一。若菌液浓度过低,可能会使农杆菌无法有效侵染叶盘而导致转化效率过低;若菌液浓度过高,则可能造成侵染过度或菌体产生代谢物对植物细胞造成伤害而导致叶盘死亡。此外,如果浓度过高,可能使农杆菌在共培养培养基上也会生长过旺,导致后续的脱菌培养无法进行。
试验中发现当菌液浓度的OD600值为0.4时,诱导率普遍较低,甚至未能成功诱导出阳性发根;菌液浓度在OD600值为0.8时虽然转化率较高,但外植体死亡率或染菌率也高,发根后续脱菌培养过程难以抑制农杆菌生长,造成无法液培。而OD600为0.6时,整体上诱导率相对较高,外植体死亡率较低,且基本不影响后续的脱菌培养及液培。因此综合取OD600=0.6为最佳菌液浓度。
2.3浸染时间对发根诱导率的影响
农杆菌菌液浸染叶盘的时间对转化率有很大影响。若浸染时间太短,可能农杆菌未能成功侵入叶盘细胞而导致转化失败;若浸染时间太长,则农杆菌容易对叶盘造成过度伤害而使叶盘黄化死亡。
试验中发现当菌液浓度较低时,浸染时间越长,则整体上诱导率越高,但叶盘死亡率或染菌率也随之增高;菌液浓度较高时,浸染时间在10 min以内,诱导率随浸染时间的增加而整体呈增加的趋势,但浸染时间达15 min时,则因叶盘大量黄化死亡而导致诱导率直线下降。因此,综合取浸染时间为10 min最佳。
2.4共培养时间对发根诱导率的影响
浸染后共培养的目的,是使叶盘表面的农杆菌充分侵入细胞。因此,共培养时间也是影响诱导率的重要因素。试验表明,菌液浓度较低,浸染时间较短时,0~3 d内,共培养时间越长,诱导率基本就越高,但同样存在随共培养时间增加,叶盘死亡率和染菌率也增高的问题;而菌液浓度较高,浸染时间较长时,共培养时间在0~2 d时,诱导率整体呈增长趋势,但超过2 d,则转化率不会再显著升高,甚至反而会因为过度生长的农杆菌抑制植物细胞的正常生长,造成叶盘死亡率急剧升高,或者因脱菌不完全而导致无法液培。因此,共培养时间以2 d为最好。
2.5不同液体培养基对发根生长速率的影响
发根脱菌完全后,转移到液体培养基中进行扩大培养。在上述4种不同的液体培养基中,发根生长速率不同,总体上是B5>1/2MS≈1/2B5>MS。在含200 mL B5液体培养基的250 mL锥形瓶中,发根整个生长周期为60~70 d,其中前3周,发根生长速率极慢,基本看不到明显的生长现象,第4周开始表现出较快的生长速率,在第5~7周生长速率达到最快,在第8~9周以后生长基本进入停滞期,10周之后发根开始变硬,部分根尖开始褐化。不同培养基生长速率比较如图3。

图3发根在不同培养基中生长速率
Fig.3The growth rate of hairy roots in different liquid mediums
3 结论与讨论
本研究利用发根农杆菌C58C1菌株侵染灯盏花叶片,成功诱导出毛状根。同时,通过对菌液浓度、浸染时间和共培养时间3个影响因素的试验,优化了灯盏花发根的诱导条件。
结果显示农杆菌菌液浓度直接影响发根诱导效率,灯盏花发根诱导的最佳菌液浓度为OD600=0.6,李景滨等[14]的研究发现,金铁锁 (Psammosilenetunicoides) 毛状根诱导的最佳菌液浓度为OD600=0.8;农杆菌对叶盘的浸染时间是遗传转化中的重要因素,灯盏花最佳浸染时间为10 min,黄伟剑等[15]的研究表明,广藿香 (Pogostemoncablin) 毛状根诱导的最佳浸染时间为25 min;共培养时间的长短也与转化效率密切相关,适于灯盏花发根诱导的共培养时间为2 d,刘连旺等[16]的研究显示,适于地黄 (Rehmanniaglutinosa) 毛状根诱导的最佳共培养时间为4 d。由此可见不同植物对农杆菌敏感性不同,因此选择合适的菌液浓度、浸染时间和共培养时间有助于提高诱导率。另外,灯盏花发根扩大培养的最佳培养基为B5液体培养基。
本研究初步建立了灯盏花发根遗传转化体系。随着对植物中次生代谢途径了解的深入,基因克隆和高效遗传转化系统的发展,利用遗传代谢工程手段在植物细胞工厂大量生产有药理活性的次生代谢化合物已成为可能。如果将灯盏花中灯盏乙素代谢途径中催化速率限制步骤的关键酶基因采用代谢工程的方法在灯盏花植物体或发根中过量表达,则理论上也可以提高灯盏乙素的含量。
[1]国家药典委员会. 中华人民共和国药典: 一部[M]. 2010年版. 北京: 医药科技出版社, 2010.
[2]Feng Y, Zhang S, Tu J, et al. Novel function of scutellarin in inhibiting cell proliferation and inducing cell apoptosis of human Burkitt lymphoma Namalwa cells[J]. Leuk Lymphoma, 2012, 53(12): 2456-2464.
[3]Chan J Y, Tan B K, Lee S C. Scutellarin sensitizes drug-
evoked colon cancer cell apoptosis through enhanced caspase-6 activation[J]. Anticancer Res, 2009, 29(8): 3043-3047.
[4]Wang S, Wang H, Guo H, et al. Neuroprotection of scutellarin is mediated by inhibition of microglial inflammatory activation[J]. Neuroscience, 2011, 185: 150-160.
[5]王平理, 杨生超, 杨建文, 等. 云南灯盏花种质资源的考察与采集[J]. 现代中药研究与实践, 2007, 22(2): 25-26.
[6]刘涛, 牟兰, 梁艳丽, 等. 灯盏花查尔酮合成酶基因表达与灯盏乙素含量关系的研究[J]. 中国中药杂志, 2013, 38(14): 2241-2243.
[7]Yang S, Wang P, Yang J. Effects of genotypic and environmental on yield and scutellarin content ofErigeronbreviscapus[J]. Chinese Agricultural Science Bulletin, 2011, 27(8): 140-143.
[8]史云东, 贾琳, 张霁, 等. 不同地区灯盏花总黄酮与野黄芩苷含量比较[J]. 安徽农业科学, 2011, 39(4): 2102, 2112.
[9]陆倍倍, 张磊, 陈万生, 等. 莨菪发根培养体系的建立[J]. 中草药, 2005, 36(12): 1864-1868.
[10]Muir S, Collins G, Robinson S, et al. Overexpression of petunia chalcone isomerase in tomato results in fruit containing increased levels of flavonols[J]. Nat Biotechnol, 2001, 19: 470-474.
[11]Zhang L, Ding R, Chai Y, et al. Engineering tropane biosynthetic pathway inHyoscyamusnigerhairy root cultures[J]. PNAS, 2004, 101: 6786-6791.
[12]Gu X C, Chen J F, Xiao Y, et al. Overexpression of allene oxide cyclase promoted tanshinone/phenolic acid production inSalviamiltiorrhiza[J]. Plant Cell Rep, 2012, 31(12): 2247-2259.
[13]Xiao Y, Ji Q, Gao S, et al. Combined transcriptome and metabolite profiling reveals that Ii PLR1 plays an important role in lariciresinol accumulation inIsatisindigotica[J]. Exbotj, 2015, 66(20): 6259-6271.
[14]李景滨, 刘同祥, 王培忠, 等. 金铁锁毛状根诱导及培养体系的建立[J]. 中国中药杂志, 2011, 36(5): 547-550.
[15]黄伟剑, 何梦玲, 张宏毅, 等. 发根农杆菌诱导广藿香毛状根的研究[J]. 广东药学院学报, 2015, 31(2): 156-160.
[16]刘连旺, 张永清, 李先恩, 等. 地黄毛状根的诱导及优化[J]. 山东农业科学, 2015, 47(1): 47-50.
(责任编辑张坤)
Establishment of Hairy Root Culture ofErigeronbreviscapus
Huang Xin1,2, Ji Qian2, Chen Ruibing2, He De1, Zhang Lei1,2
(1. College of Life Sciences, Southwest Forestry University, Kunming Yunnan 650224, China;2. Department of Pharmacy, Second Military Medical University, Shanghai 200433, China)
To establish the cultural system for hairy root ofErigeronbreviscapusroot ofE.breviscapuswas obtained from leaf explants after infected withAgrobacteriumrhizogenesC58C1 strains, determine the growth curve of hairy roots, and compared the effect of hairy roots increment by different factors. The results showed that the hairy root ofE.breviscapuswas successfully induced by usingA.rhizogenesstrain C58C1, and the best system of hairy root culture was established. The best concentration ofA.rhizogenesliquid is OD600=0.6, the best infection time is 10 min, and the best co-culture time is 2 d. These conditions allow the inducement rate was 60%. The transformation of T-DNA from Ri plasmid to the hairy root was confirmed by PCR analysis.
Agrobacteriumrhizogenes, induction rate, genetic transformation system,Erigeronbreviscapus, culture medium

10. 11929/j. issn. 2095-1914. 2016. 05. 004
2016-02-02
张磊 (1977—),男,博士,副教授,硕士生导师。研究方向:中药药效物质基础及品质评价研究。Email: leizhang100@163.com。
S723.1
A
2095-1914(2016)05-0021-06
第1作者:黄鑫 (1990—),男,硕士生。研究方向:药用植物次生代谢工程。Email: huangxin6635@qq.com。
