鹅掌楸属(木兰科)次生木质部导管穿孔板的比较研究
2016-10-14李红芳刘江梅
李红芳,刘江梅
(1 渭南职业技术学院 农学院,陕西渭南 714021;2 陕西师范大学 生命科学学院,西安 710119; 3 榆林学院 生命科学学院,陕西榆林 719000)
鹅掌楸属(木兰科)次生木质部导管穿孔板的比较研究
李红芳1,2,刘江梅3
(1 渭南职业技术学院 农学院,陕西渭南 714021;2 陕西师范大学 生命科学学院,西安 710119; 3 榆林学院 生命科学学院,陕西榆林 719000)
利用扫描电子显微镜对鹅掌楸属仅存的2个自然种鹅掌楸和北美鹅掌楸的次生木质部导管穿孔板特征进行了详细的研究。结果显示,鹅掌楸属的2个种均以梯状穿孔板为主,同时存在网状-梯状混合穿孔板。鹅掌楸和北美鹅掌楸的导管穿孔板具有明显差异:(1)鹅掌楸具网状穿孔板,而北美鹅掌楸没有观察到;(2)北美鹅掌楸具单穿孔板,而鹅掌楸在该实验中未发现;(3)北美鹅掌楸具有横隔较粗的梯状穿孔板且横隔数目较多;(4)鹅掌楸的导管穿孔板多数横隔较少;(5)北美鹅掌楸的穿孔板倾斜角度较大;(6)北美鹅掌楸具麻黄式穿孔板的存在,且有纹孔膜残留存在。研究认为,北美鹅掌楸导管分子穿孔板分化较鹅掌楸更为剧烈。
鹅掌楸,北美鹅掌楸,木兰科,穿孔板,导管分子
植物的形态学研究一直是系统分类学研究的核心内容,因此,系统分类性状的发掘与分析、重新认识与评价就成了系统分类学家最重要的工作内容。作为被子植物创新性状之一的导管的形态研究,也是形态学研究的重要部分。木兰科属于基部被子植物木兰分支(Magnoliids)[1],其次生木质部导管分子的研究,对于基部被子植物的基础特征及木兰科的系统发育及演化分析都具有重要意义,并且作为有重要应用价值的树种,其研究对木材的鉴定也有重要作用。
鹅掌楸属(Liriodendron)是木兰科(Magnoliaceae)中比较特化的类群[2-6]。该属现仅存鹅掌楸(Liriodendronchinense(Hemsl.) Sarg.)和北美鹅掌楸(LiriodendrontulipiferaLinn.)2个种。鹅掌楸现主要分布于中国,延伸至越南边境,而北美鹅掌楸则主要分布于美国东部[7-8],它们被认为是分化相当好的2个种,在属内形成1对典型的东亚-北美间断分布的对应种[8-9]。由于该属树种是研究群体遗传结构、杂交和杂种优势、植物系统与进化、植物地理学等领域的理想材料,因此,国内外众多学者对其进行了大量的研究[10-16],其中也有一些木质部的研究[17-22],但这些研究中,木质部的研究都是利用光学显微镜进行观察的,缺乏扫描电子显微镜(SEM,以下简称扫描电镜)下对木质部尤其是对导管及其穿孔板进行的详细系统的研究。扫描电镜的应用对于导管的研究具有重要意义,如草珊瑚、水青树、昆栏树等无导管类群中导管分子的发现[23-27]、纹孔膜残留的发现[28]、蕨类植物导管的发现[29]等。因此,本文利用扫描电镜对鹅掌楸属仅存2个自然种的次生木质部导管穿孔板的特征进行详细观察,并对其导管穿孔板进行比较,以期为该属的研究提供更为详尽的系统学资料,也为木材的鉴定提供解剖学依据。
1 材料和方法
1.1 材 料
实验材料选取3年生以上枝。鹅掌楸取自陕西镇坪野生种(凭证标本:赵亮090504,SANU);北美鹅掌楸材料取自加拿大的Nova Scotia, Saint Mary’s University(凭证标本:任毅AUS001,SANU)。凭证标本存放于陕西师范大学生命科学学院植物标本室(SANU)。
1.2 方 法
1.2.1 离析材料处理 将材料以FAA固定,切成1cm长火柴棍粗细的细条,用Jeffrey 离析液(10%铬酸∶10%硝酸= 1∶1),在40 ℃下离析约48 h,离析后用清水冲洗备用。
1.2.2 观察方法 离析材料铺于载玻片,自然干燥,喷金镀膜后在Hitachi S-570扫描电子显微镜(SEM)下观察。
2 观察结果
2.1 鹅掌楸
鹅掌楸导管侧壁无螺纹加厚(图版Ⅰ,1~4),穿孔板多为典型的梯状,较短,倾斜角度比较小(图版Ⅰ,5~15);横隔数目较少且细,横隔间穿孔较大(图版Ⅰ,5~17)。横隔数目从2~6不等(图版Ⅰ,5~15),偶尔会出现1个横隔(图版Ⅰ,17)。穿孔板具短喙(图版Ⅰ,5、14)或不具喙(图版Ⅰ,6、9、11、12),少数情况会有很长的喙部(图版Ⅱ,1),其喙上可能具小孔(图版Ⅱ,1)。倾斜角度较小,有的穿孔板接近平截(图版Ⅰ,16)。
梯状穿孔板会出现横隔分叉的情况(图版Ⅰ,12、16;图版Ⅱ,1)。偶尔可以见到网状穿孔板(图版Ⅱ,6)。横隔较粗的梯状穿孔板(图版Ⅱ,7)较少出现,多位于较细的导管上。
网状-梯状混合的穿孔板(图版Ⅱ,2~5)存在,梯状部分横隔较细,网状部分可能位于梯状下部(图版Ⅰ,11)、上部(图版Ⅱ,5)或上下都存在(图版Ⅱ,2~4右)。
此外,还观察到一种奇怪的穿孔板,上、下各有一个较大穿孔,中间被一段与侧壁相似的超大横隔分隔,看起来似乎象在一个大的穿孔中央有一段与侧壁相同的壁覆盖其上(图版Ⅱ,8)。
侧壁上可以看到网状穿孔板的存在(图版 Ⅰ,3、4)。
2.2 北美鹅掌楸
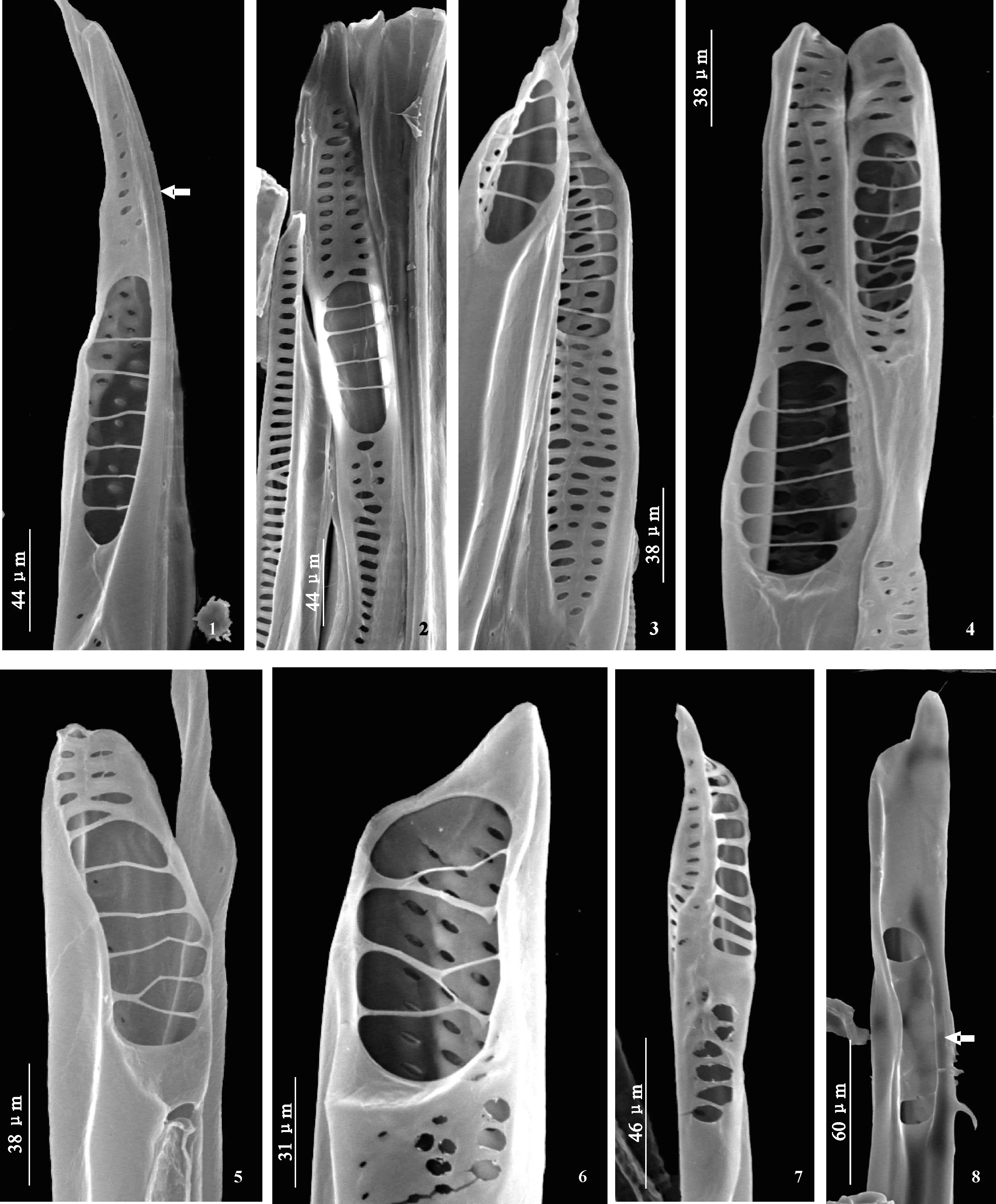
北美鹅掌楸导管侧壁同样无螺纹加厚(图版 Ⅲ,1~3),穿孔板也以典型的梯状为主,同时也观察到拟梯状、网状-梯状混合的穿孔板、单穿孔板等多种类型的穿孔板。
穿孔板多数为梯状穿孔板,横隔粗细、数目、穿孔的大小以及穿孔板倾斜的角度都有差异。有的梯状穿孔板倾斜角度比较大(图版Ⅲ,6、7、13)。梯状穿孔板的横隔数目从一到几十不等(图版Ⅲ,1~13)。
横隔粗细均匀的梯状穿孔板多数较短,横隔很细且数目较少(图版Ⅲ,1~5、7),横隔中有的细如丝(图版Ⅲ,1~4),有的稍粗(图版Ⅲ,5、7);横隔间穿孔较大(图版Ⅲ,1~5、8~12)。具1个横隔的穿孔板比较多(图版Ⅲ,1上、下,11,13左);穿孔板具短喙(图版Ⅲ,1上,9)或不具喙(图版Ⅲ,1下,4、5),也会有较长的喙部(图版Ⅲ,11、12、17)。横隔较粗的梯状穿孔板较长,横隔数量较多,横隔间穿孔较小(图版Ⅲ,6、7、13;图版 Ⅳ,3),同一穿孔板上穿孔大小也有差异。
此外,在梯状穿孔板类型中,有的在同一穿孔板上横隔粗细差异很大(图版Ⅲ,14),同时,有的穿孔板中2列梯状与横隔较细的梯状共同存在(图版 Ⅳ,2)。
网-梯混合的梯状穿孔板类型中,横隔较细的梯状穿孔板与网状同时出现在同一穿孔板上(图版Ⅲ,16;图版 Ⅳ,1)。网状横隔一般较粗,穿孔较小,与梯状部分差异明显。
梯状穿孔板中有些穿孔呈2列分布(图版Ⅳ,4),横隔的粗细各有不同。麻黄式穿孔板存在,其上穿孔只是形成小孔,甚至分成3列分布,且较长(图版Ⅲ,15;图版 Ⅳ,6)。
北美鹅掌楸中观察到了单穿孔板,具喙(图版 Ⅳ,9~10)或不具喙(图版Ⅳ,11)。在单穿孔带喙的穿孔板中,也观察到了喙上具很小的穿孔(图版Ⅳ,11)。
在北美鹅掌楸中,部分穿孔板中观察到纹孔膜的存在(图版Ⅳ,5、7、8)。这些穿孔板上的穿孔一般会小一些。偶尔也可以观察到侧壁穿孔板存在(图版Ⅲ,3)。
3 分析与讨论
3.1 鹅掌楸与北美鹅掌楸穿孔板的特征及比较
通过扫描电镜的详细观察,发现鹅掌楸属2个种都以梯状穿孔板为主,且都不具螺旋加厚,同时存在网-梯混合的穿孔板。网-梯混合的穿孔板与北美鹅掌楸中观察到的单穿孔板与麻黄式穿孔板都是首次报道。以前的报道一般认为鹅掌楸属是梯状穿孔板[30-31],仅Brown曾有过网状穿孔板的报道[17]。王丰等通过光镜对鹅掌楸属的解剖学特征进行了观察,认为在鹅掌楸中单穿孔板存在,而在北美鹅掌楸中未发现单穿孔板[22]。本研究则在北美鹅掌楸中发现了单穿孔板,麻黄式穿孔板也存在;同时,北美鹅掌楸中具有纹孔膜残留。另外在鹅掌楸中出现了穿孔板形态的个例(穿孔板中间出现的壁状物)。
同时,北美鹅掌楸与鹅掌楸的穿孔板也具有明显的差异(表1),北美鹅掌楸的穿孔板较鹅掌楸类型多,分化也更加明显。具体来说:
(1)在鹅掌楸中,观察到了网状穿孔板的存在(图版Ⅱ,6),北美鹅掌楸中,则并未见到;(2)北美鹅掌楸中观察到了比较原始的近似于麻黄式穿孔板的存在(图版Ⅳ,6),鹅掌楸中未发现;(3)北美鹅掌楸中发现了单穿孔板(图版Ⅳ,9~11),而在实验中鹅掌楸中并未发现单穿孔板的存在。虽然王丰等通过光学显微镜观察认为鹅掌楸中有单穿孔板的存在,但由于本实验中没有发现单穿孔板,故不适合在不一样的实验条件下进行比较;(4)在北美鹅掌楸中梯状穿孔板出现1个横隔的比例较鹅掌楸中大;在鹅掌楸导管分子上,横隔≤4的梯状穿孔板的导管分子占具梯状穿孔板导管分子的75.1%,而北美鹅掌楸仅占22.4%,即在鹅掌楸中,以具1~4横隔的梯状穿孔板为主,其他的也多数具5~6个横隔;(5)在北美鹅掌楸中观察到了明显的纹孔膜的存在(图版Ⅳ,5、7、8),而在鹅掌楸的穿孔板中并未见到;(6)北美鹅掌楸的穿孔板倾斜角度总体较大,没有观察到鹅掌楸中几乎平截的穿孔板(图版Ⅰ,16)。(6)在鹅掌楸穿孔板中,典型的梯状穿孔板,多横隔较细,穿孔较大,未观察到横隔较粗的梯状穿孔板;在北美鹅掌楸中则横隔较粗的梯状穿孔板较为常见(图版Ⅲ,6、7、13;图版Ⅳ,3)。

表1 鹅掌楸属穿孔板的比较
这些特征明确表明了2个种的穿孔板存在明显差异。在北美鹅掌楸中观察到了演化程度最高的单穿孔板的存在,又同时包含麻黄式穿孔板、横隔较粗的梯状穿孔板、不典型的梯状穿孔板以及残存的纹孔膜等较原始的特征;鹅掌楸的梯状穿孔板横隔少、特化程度高,然而并未观察到单穿孔板存在。由此可见,北美鹅掌楸的次生木质部的导管穿孔板分化更为剧烈,特征中同时包含了原始性状与演化性状,说明这些木质部的特征或为平行演化。
3.2 鹅掌楸与北美鹅掌楸的系统关系
韦仲新和吴征镒、刘玉壶等认为鹅掌楸原始[12-13],惠利省观察到北美鹅掌楸存在更为进化的花粉皱疵状纹饰,因此也推测北美鹅掌楸比鹅掌楸更为进化[15],这与韦仲新和吴征镒、徐进等在对鹅掌楸属花粉外壁的超微结构研究[12,32]中得到结论相一致。而方炎明认为他们是姊妹群[8]。罗群凤利用基于Chs、Adh序列的结果表明鹅掌楸进化速率快于北美鹅掌楸[17],此分子的研究结论与之前的形态研究观点不一致,然而这个结论得到本实验与王丰等对其解剖学结构观察结果[23]的支持。虽然王丰等基于鹅掌楸单穿孔的存在,北美鹅掌楸单穿孔板不存在,认为鹅掌楸较北美鹅掌楸进化,但根据横隔数目及射线也可支持此结论[22]。从以上可以看出,鹅掌楸属原始或进化的特征并没有集中在同一个类群中,而是互相嵌合。本实验结果中北美鹅掌楸的次生木质部的解剖学特征同时包含了原始性状与进化性状,分化更加剧烈些,此结果与罗光佐等利用RAPD标记技术所做的遗传多样性分析的结果相吻合[33]。由于他们的亲缘关系可追溯到上白垩纪,2个种的分化产生于中新世纪,2个种可以自由交配,自中新世纪晚期,分布区开始分裂,产生了地理隔离[7,10]。鹅掌楸的遗传变异主要来自于地理种源内,而北美鹅掌楸的遗传变异主要来自于地理种源间[33],结合他们穿孔板的特征,即鹅掌楸的穿孔板处于演化的较高阶段,但未形成单穿孔板,而北美鹅掌楸则形成了单穿孔板,却仍存在穿孔板演化的早期(麻黄式穿孔板)与中期(横隔较多的梯状穿孔板)阶段,可以推测木质部的特征或为平行进化,由于生活环境(北美鹅掌楸较鹅掌楸纬度上高8度)和进化速率不同,而产生了不同的导管分子与穿孔板特征,形成了2个种的穿孔板特征的差异,而这种差异也表明北美鹅掌楸与鹅掌楸的导管演化是各自独立地进行的,也可以推论,这种差异可能来自于地理隔离之后。当然,这个问题还需要更多角度的证据。
[1] The Angiosperm Phylogeny Group. An update of the Angiosperm Phylogeny Group classification for the orders and families of flowering plants: APGIII[J].Bot.J.Linn.Soc, 2009, 161(10): 105-121.
[2] METCALFE C R. Anatomy of the dicotyledons, 2ed edn, Volume III. Magnoliales, Illiciales, and Laurales[M]. Oxford: Clarendon Press, 1987.
[3] QIU Y L, CHASE M W, PARKS C R. A chlorplast DNA phylogenetic study of the easterm Asia-eastern North America Disjunct section Rytidospermun ofMagnolia(Magnoliaceae)[J].Amer.J.Bot., 1995, 82(12): 1 582-1 588.
[4] 李 捷. 木兰科植物的分支分析[J]. 云南植物研究, 1997, 19(4): 342-356.
LI J. A cladistic analysis of magnoliaceae[J].ActaBotanicaYunnanica, 1997, 19(4): 342-356.
[5] 张秀军,陈玉芹,柴凤瑞,等. 中国木兰科各属次生木质部的解剖特征及系统演化[J]. 木本植物研究, 2000, 20(3): 318-323.
ZHANG X J, CHEN Y Q, CHAI F R,etal. Anatomical characteristics and phylogenetic relationships of secondary xylem of Magnoliaceae in China[J].BulletinofBotanicalResearch, 2000, 20(3): 318-323.
[6] 徐凤霞,徐信兰,胡晓颖. 五种木兰属植物的花粉形态[J]. 云南植物研究, 2004, 26(1):83-88.
XU F X, XU X L, HU X Y. Pollen morphology of five species fromMagnolia[J].ActaBotanicaYunnanica, 2004, 26(1):83-88.
[7] 方炎明. 中国鹅掌楸的地理分布与空间格局[J]. 南京林业大学学报, 1994, 18(2): 13-18.
FANG Y M. Geographical distribution and spatial pattern ofLiriodendronchinense(Hemsl.) Sarg[J].JournalofNanjingForestryUniversity, 1994, 18(2): 13-18.
[8] 李 帅. 鹅掌楸属植物基因差异表达与杂种优势的关系[D]. 南京:南京林业大学, 2009.
[9] 朱晓琴,马建霞,姚青菊,等. 鹅掌揪 (Liriodendronchinense) 遗传多样性的等位酶论证[J]. 植物资源与环境, 1995, 4(3): 9-14.
ZHOU X Q, MA J X, YAO Q J,etal. Allozyme verification on the polulation differentiation ofLiriodendronchinense(Hemsl.) Sarg.[J].JournalofPlantResourcesandEnvironment, 1995, 4(3): 9-14.
[10] PARKS C R, WENDEL J F. Molecular divergence between Asian and North Amercian species ofLiriodendron(Magnoliaceae) with implications for interpretation of fossil floras[J].Amer.J.Bot., 1990, 77(10): 1 243-1 256.
[11] PARKS C R, WENDEL J F, SEWELL M M,etal. Genetic control of isozyme variation in the genusLiriodendronL. (Magnoliaceae)[J].J.Hered., 1990, 81(4): 317-323.
[12] 韦仲新,吴征镒. 鹅掌揪属花粉的超微结构研究及系统学意义[J]. 云南植物研究, 1993, 15(2): 163-166.
WEI Z X, WU Z Y. Pollen ultra structure ofLiriodendronand its systematic significance[J].ActaBotanicaYunnanica, 1993, 15(2): 163-166.
[13] 刘玉壶,夏念和,杨惠秋. 木兰科(Magnoliaceae)的起源、进化和地理分布[J]. 热带亚热带植物学报, 1995, 3(4): 1-12.
LIU Y H, XIA N H, YANG H Q. The origin, evolution and phytogeography of Magnoliaceae[J].JournalofTropicalandSubtropicalBotany, 1995, 3(4): 1-12.
[14] CAI Z Q, PENAFLOR C, KUEHL J V,etal. Complete plastid genome sequences ofDrimys,Liriodendron, andPiper: implications for the phylogenetic relationships of magnoliids[J].BMC.Evol.Biol., 2006,(6): 77.
[15] 惠利省. 马褂木遗传多样性及系统地理学研究[D]. 南京:南京林业大学,2010.
[16] 罗群凤. 鹅掌楸属基因进化差异研究——基于Chs、Adh序列证据[D]. 南京:南京林业大学, 2013.
[17] BROWN F B H. Scalariform pitting a primitive feature in angiospermous secondary wood[J].Science, 1918, 1 227(48): 16-18.
[18] 陈静波. 几种木兰科植物的比较解剖[J]. 玉溪师专学报(自然科学版), 1990,(5): 91-98.
CHEN J B. Comparative anatomical studies of some species in Magnoliaceae[J].JournalofYuxinTeachers’College, 1990,(5): 91-98.
[19] 吴树明,林金星,李正理. 中国木兰科9 属6 6种植物木射线的比较解剖学研究[J]. 植物学报, 1993, 35(4): 268-279.
WU S M, LIN J X, LI Z L. Comparative anatomical studies on the wood rays of sixty-six species in nine genera of Magnoliaceae in China[J].ActaBotanicaSinica, 1993, 35(4): 268-279.
[20] 韩丽娟,林月惠,吴树明. 不同纬度对鹅掌楸次生木质部结构的影响[J]. 植物学通报, 2001, 18(3):375-377.
HAN L J, LIN Y H, WU S M. The effect of latitudes on the structure of secondary xylem inLiriodendronchinenseSarg[J].ChineseBulletinofBotany, 2001, 18(3):375-377.
[21] 潘 彪,徐朝阳,王章荣. 杂种鹅掌楸木材解剖性质及其径向变异规律[J]. 南京林业大学学报(自然科学版), 2005, 29(1): 79-82.
PAN B, XU C Y, WANG Z R. Wood anatomical properties and its radial variation ofLiriodendronchinense×L.tulipifera[J].JournalofNanjingForestryUniversity(Nat. Sci. Edi.), 2005, 29(1): 79-82.
[22] 王 丰,潘 彪,唐 菁,等. 鹅掌楸属次生木质部解剖特征及自然种系统演化[J]. 安徽农业大学学报, 2014, 41(3): 451-455.
WANG F, PAN B, TANG J,etal. Comparative study on secondary xylem structure inLiriodendronand its implication on phylogeny[J].JournalofAnhuiAgriculturalUniversity, 2014, 41(3): 451-455.
[23] CARLQUIST S. Presence of vessels in wood ofSarcandra(Chloranthaceae); comments on vessel origins in angiosperms[J].Amer.J.Bot., 1987, 74(12): 1 765-1 771.
[24] TAKAHASHI A. Morphology and ontogeny of stem xylem elements inSarcandraglabra(Thunb.) Nakai (Chloranthaceae): Additional evidence for the occurrence of vessels[J].Bot.Mag., 1988, 101(4): 387-395.
[25] 张遂申,雷立功,刘宏颀,等. 中国特有植物海南草珊瑚管状分子的初步研究——草珊瑚属导管存在的例证[J]. 西北植物学报, 1990, 10(2): 95-98.
ZHANG S S, LEI L G, LIU H X,etal. A preliminary study on tracheary elements in the endemic speciesSarcandrahainanensisfrom China——evidence for the occurrence of vessels inSarcandra[J].ActaBotanicaBoreali-OccidentaliaSinica, 1990, 10(2): 95-98.
[26] REN Y, CHEN L, TIAN X H,etal. Discovery of vessels inTetracentron(Trochodendraceae) and its systematic significance[J].PlantSyst.vol., 2007, 267(1): 155-161.
[27] LI H F, CHAW S M, DU C M,etal. Vessel elements present in the secondary xylem ofTrochodendronandTetracentron(Trochodendraceae)[J].Flora, 2011, 206(6): 595-600.
[28] 李红芳,任 毅. 领春木茎次生木质部中导管穿孔板的变异[J]. 植物分类学报, 2005, 43(1): 1-11.
LI H F, REN Y. The variation of perforation plates of vessels in the secondary xylem ofEupteleapleiosperma(Eupteleaceae)[J].ActaPhytotaxonomicaSinica, 2005, 43(1): 1-11.
[29] CARLQUIST S, SCHNEIDER E L. SEM studies of vessels in ferns 14[J].AquatBot., 2000, 66(1): 1-8.
[30] CARLQUIST S. Comparative Wood Anatomy[M]. Berlin: Springer-Verlag, 1988.
[31] 吴树明,李正理. 八属木兰科植物木材导管分子的比较解剖[J]. 植物学报, 1988, 30(1):33-39.
WU S M, LI Z L. Comparative anatomical studies on the vessel elements of eight genera in Magnoliaceae[J].ActaBotanicaSinica, 1988, 30(1):33-39.
[32] 徐 进,王章荣. 鹅掌揪属种间杂种与亲本花粉壁的超微结构的比较观察[J]. 植物学通报, 2005, 22(1): 35-38.
XU J, WANG Z R. Comparison survey of the morphology and ultrastructure of pollens fromLiriodendronhybrids and their parents[J].ChineseBulletinofBotany, 2005, 22(1): 35-38.
[33] 罗光佐,施季森,尹佟明,等. 利用RAPD标记分析北美鹅掌楸与鹅掌楸种间遗传多样性[J]. 植物资源与环境学报, 2000, 9(2): 9-13.
LUO G Z, SHI J S, YIN T M,etal. Comparison of genetic diversity betweenLiriodendrontulipiferaLinn .andLiriodendronchinense(Hemsl.) Sarg. by means of RAPD markers[J].JournalofPlantResourcesandEnvironment, 2000, 9(2): 9-13.
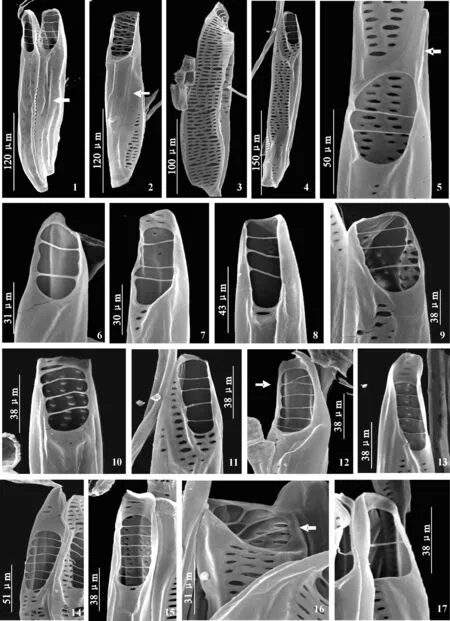
图版Ⅰ 鹅掌楸的导管及穿孔板形态1. 2个导管分子的整体观,梯状穿孔板,箭头示侧壁无螺旋加厚;2. 1个导管分子整体观,箭头示侧壁无螺旋加厚;3. 示网状侧壁;4. 侧壁穿孔;5. 梯状穿孔板,具较长的喙(箭头所示);6. 具2个横隔的梯状穿孔板;7. 具3个横隔的梯状穿孔板,具短喙;8、9. 横隔为3的梯状穿孔板,不具喙;10. 梯状穿孔板,横隔为4,上面1个横隔较粗;11. 横隔为4,其下具穿孔;12. 横隔为5的穿孔板,箭头示横隔分叉;13. 横隔为4的梯状穿孔板,具喙;14. 梯状穿孔板,具较长的喙;15. 梯状穿孔板,横隔为6;16. 箭头示倾斜角度小,接近平截的穿孔板,其上有横隔的联合;17. 仅具1个横隔的穿孔板PlateⅠ Vessel elements and the perforation plates in secondary xylem of L. chinense under SEM.Fig.1. Two whole vessel elements with scalariform perforation plates, and arrow shows no spirally thicken on the lateral wall. Fig.2. A whole vessel element, arrow shows no spirally thicken on the lateral wall. Fig.3. The perforation plate with reticulate on lateral wall. Fig.4. Perforations in the lateral wall. Fig.5. The scalariform perforation plate with long tip (arrow). Fig.6. The scalariform perforation plate with two bars. Fig.7. The scalariform perforation plate with three bars and short tip. Fig.8-9. The scalariform perforation plate, no tips. Fig.10. The scalariform perforation plate with four bars, the first bar is wider than others. Fig.11. The scalariform perforation plate with four bars, with perforation under it. Fig.12. The scalariform perforation plate with five bars, arrow shows the forked bar. Fig.13. The scalariform perforation plate with four bars and tip. Fig.14. The scalariform perforation plate with longer tip. Fig.15. The scalariform perforation plate with six bars. Fig.16. Less obliquity perforation plate with forked bars. Fig.17. The scalariform perforation plate with one bar
图版 Ⅱ 鹅掌楸的穿孔板形态1. 示梯状穿孔板具长喙 (箭头);2~5. 网梯混合的穿孔板;6. 网状穿孔板;7. 梯状穿孔板,横隔为8,横隔较粗,8. 在一个大的穿孔中央出现一长截与侧壁相同的壁(箭头所示)。Plate Ⅱ Perforation plates in secondary xylem of L. chinense under SEMFig.1. The scalariform perforation plate with long tip. Fig.2-5. Reticulate- scalariform perforation plates. Fig.6. Reticulate perforation plate. Fig.7. The scalariform perforation plate with 8 bars, bars are a little wide. Fig.8. A perforation with wall as well as lateral wall in the middle.
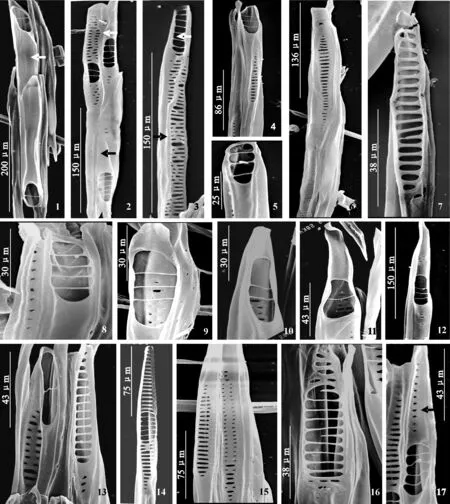
图版 Ⅲ 北美鹅掌楸导管及其穿孔板形态 1. 2个导管分子,梯状穿孔板(上、下),仅1个横隔,不具喙,侧壁无螺旋加厚(箭头);2. 导管的整体观,侧壁无螺旋加厚(黑色箭头所示),左上为混合穿孔板(白色箭头所示),右上为梯状穿孔板,具喙,横隔数目为4,右下示梯状穿孔板,具喙,横隔数目为5;3. 梯状穿孔板(白色箭头),横隔数目为6;黑色箭头示侧壁穿孔板;4. 梯状穿孔板,横隔数目为3,横隔很细;5. 梯状穿孔板,横隔数目为3,横隔稍粗;6. 梯状穿孔板较长,穿孔较小,横隔粗;7. 梯状穿孔板较长,穿孔较大,横隔较多且粗;8. 梯状穿孔板,横隔数目为4,不具喙;9. 梯状穿孔板,横隔数目为3,具短喙;10. 梯状穿孔板,横隔数目为2,具短喙;11. 梯状穿孔板,横隔数目为1,具长喙;12. 梯状穿孔板,横隔数目为3,具长喙;13. 梯状穿孔板,左示穿孔板仅一横隔(左),横隔细,不具喙;右示长梯状穿孔板,多横隔;14. 梯状穿孔板,横隔粗细不均匀,中间横隔较细,上部为梯状,下部为拟梯状;15. 梯状穿孔板(左),穿孔板较长,穿孔较小,横隔较多且较宽;右侧穿孔板示穿孔呈孔状,非常小,呈拟梯状排列;16. 混合穿孔板,下部为横隔较细的梯状,上部为近网状或拟梯状;17. 梯状穿孔板,横隔数目为4,具喙,其喙上具均匀穿孔(箭头所示)。Plate Ⅲ Vessel elements and the perforation plates in secondary xylem of L. tulipifera under SEMFig.1. Two whole vessel elements with scalariform perforation plates(upper, lower), one bar, no tip; no spirally thicken on the lateral wall (arrow). Fig.2. Whole vessels, no spirally thicken on the lateral wall (blank arrow), mix perforation plate in the upper left, the scalariform perforation plate with tip and four bars in the upper right, the scalariform perforation plate with tip and five bars in the lower right. Fig.3. The scalariform perforation plate with six bars (white arrow), the blank arrow shows the lateral wall. Fig.4. The scalariform perforation plate with three bars and the bars are narrow. Fig.5. The scalariform perforation plate with three bars and the bars are wide. Fig.6. The scalariform perforation plate is long, the perforations are small and the bars are wide. Fig.7. The scalariform perforation plate is long, the perforations are big and the bars are many and wide. Fig.8. The scalariform perforation plate with four bars, no tip. Fig.9. The scalariform perforation plate with three bars, short tip. Fig.10. The scalariform perforation plate with two bars, short tip. Fig.11. The scalariform perforation plate with one bar, long tip. Fig.12. The scalariform perforation plate with three bars, long tip. Fig.13. The scalariform perforation plate, one bar (left), the bar are narrow, no tip; the perforation plate is long and the bars are many (right). Fig.14. The scalariform perforation plate, the bars are different, the bars are narrow in the middle and the scalariform in the upper and the scalariform like in the lower. Fig.15. The scalariform perforation plate, the plate (left) is long, the scalariform-like perforation (right) is small, the bars are many and wide. Fig.16. The mix perforation plate, the scalariform with narrow bars in the lower and the reticulate in the upper. Fig.17. The scalariform perforation plate with four bars, evenly perforations on tip (arrow)

图版 Ⅳ 北美鹅掌楸穿孔板形态1. 梯状穿孔板,中间穿孔较大,横隔较细,上部和下部横隔较粗,拟梯状或网状;2. 梯状穿孔板,上部横隔较细,下部横隔较粗且分为两列;3. 梯状穿孔板,较长,横隔较粗;4. 拟梯状穿孔板;5. 梯状穿孔板,横隔粗细不均匀,具纹孔膜残留(箭头所示);6. 麻黄式穿孔板,较长;7. 梯状穿孔板,穿孔较窄,具纹孔膜(箭头所示);8. 示纹孔膜残留;9. 单穿孔板,具喙,喙上有穿孔;10. 单穿孔板,具短喙;11. 单穿孔板,无喙。Plate Ⅳ Perforation plates in secondary xylem of L. tulipifera under SEMFig.1. The scalariform perforation plate, the perforations in the middle are big and the bars are narrow, the wide bars in the upper and lower. Fig.2. The scalariform perforation plate, the bars in the upper are narrow and the bars in the lower are wide. Fig.3. The scalariform perforation plate,long and wide bars. Fig.4. The scalariform-like perforation plate. Fig.5. The scalariform perforation plate, the bars are different, arrow shows pit membrane remains. Fig.6. Epledroidforation. Fig.7. The scalariform perforation plate, the perforations are narrow, arrow shows pit membrane remains. Fig.8. Pit membrane remains. Fig.9. Simple perforation plate with tip, some perforations on tip. Fig.10. Simple perforation plate with short tip. Fig.11. Simple perforation plate, no tip
(编辑:潘新社)
Comparative Investigation of Vessel Elements in the Secondary Xylem ofLiriodendron
LI Hongfang1,2, LIU Jiangmei3
(1 College of Agriculture, Weinan Vocational and Technical College, Weinan, Shaanxi 714021, China; 2 College of Life Sciences, Shaanxi Normal University, Xi’an 710119, China; 3 College of Life Sciences, Yulin College, Yulin, Shaanxi 719000, China)
The perforation plates of vessel elements in secondary xylem ofLiriodendronwhich include 2 species were observed in detail by scanning electron microscope (SEM), and many details were observed that were not reported by light microscope. Reticulate-scalariform perforation plates exist inLiriodendron, and the simple perforation plate, epledroidforation and pit membrane remnants were observed inL.tulipifera. The results showed: (1) forLiriodendronchinense(Hemsl.) Sarg., the perforation plates are typically scalariform with a few bars and reticulate-scalariform perforation plates, but occasionally are reticulate perforation plate; (2) forLiriodendrontulipifera, the perforation plates are scalariform perforation plates and reticulate-scalariform perforation plates, but occasionally are simple perforation plates; (3) part of scalariform perforation plates inL.tulipiferawith wider and more bars; (4) perforation plates with a few bars are more inL.chinensethan inL.tulipifera; (5) the perforated plates inL.tulipiferaare more inclined than inL.chinense; (6) epledroidforation and pit membrane remnants were observed inL.tulipifera. The conclusion is that the perforation differentiation ofL.tulipiferais more violent.
Liriodendron;LiriodendrontulipiferaLinn.; Magnoliaceae; perforation plate; vessel elements
1000-4025(2016)08-1585-09
10.7606/j.issn.1000-4025.2016.08.1585
2016-05-26;修改稿收到日期:2016-06-28
陕西省教育厅科学研究计划专项(16JK2052)
李红芳(1975-),女,博士,副教授,从事植物系统发育研究。E-mail: hflee@126.com
Q944.66
A
