茶树咖啡碱合成代谢中N-甲基转移酶的研究进展
2016-10-13王荣秀江丽娜邓威威张正竹
王荣秀,江丽娜,邓威威*,张正竹
茶树咖啡碱合成代谢中N-甲基转移酶的研究进展
王荣秀,江丽娜,邓威威*,张正竹
(安徽农业大学茶树生物学与资源利用国家重点实验室,安徽合肥 230036)
咖啡碱是茶叶中的主要品质成分之一,是一种黄嘌呤生物碱化合物,约占茶叶干重的2%-5%。在茶树中咖啡碱的核心合成途径为:黄嘌呤核苷甲基化反应生成7-甲基黄嘌呤核苷,再通过脱核糖反应生成7-甲基黄嘌呤,然后经过两次甲基化依次生成可可碱和咖啡碱。咖啡碱合成途径中转甲基化反应是通过N-甲基转移酶催化进行的,该酶是咖啡碱合成的关键酶。本文综述了茶树中N-甲基转移酶的基因克隆、结构、功能分析及表达调控等方面的研究进展。
茶树;咖啡碱;N-甲基转移酶
咖啡碱是茶、咖啡和可可世界三大软饮料植物中共有的重要功能性成分之一,同时也是茶树中主要的生化活性物质,约占茶叶干重的2%-5%。19世纪20年代在咖啡和茶中发现并分离得到咖啡碱[1],20世纪60年代以来对咖啡碱在茶树中的代谢途径研究有了很大的进展,其生物合成途径也逐步的明晰起来。Ashihara等[2]用同位素示踪法证明了茶树中咖啡碱生物合成与茶树的生长发育有着密切的关系。咖啡碱在生物体内的核心合成途径为[3-4]:黄嘌呤核苷(Xanthosine,XR)甲基化反应生成7-甲基黄嘌呤核苷(7-methylxanthosine,7-mXR),再通过脱核糖反应生成7-甲基黄嘌呤(7-methylxanthine,7-MX),然后经过两次甲基化作用依次生成可可碱(Theobromine,Tb)和咖啡碱(Caffeine,Cf)。途径中转甲基化反应是通过N-甲基转移酶(N-methyltransferases,NMTs)催化进行的,NMTs是茶树咖啡碱合成的关键酶。2014年晏嫦妤[5]等综述了植物中NMTs的酶学特性、克隆、基因结构及表达等,本文通过近年来学者的研究对茶树NMTs基因克隆、结构、功能分析及表达调控等方面的深入研究,了解其分子机制,全面解析NMTs对茶树咖啡碱生物合成的影响。
1茶树中咖啡碱的概况
咖啡碱(1,3,7-三甲基黄嘌呤)是一种含氮化合物,茶树中的生物碱以嘌呤碱为主体,而咖啡碱又是构成嘌呤碱的主体,因此咖啡碱在茶树次生代谢中起到了举足轻重的作用。自然界中富含咖啡碱的植物为数不多,茶树就是其中的一个高等植物。茶树中的咖啡碱分布与其他植物也略有不同,在其他植物中咖啡碱主要分布在果实和种子中,叶片中的含量极少。而茶树中的咖啡碱除了种子外,在其他部位均有分布,其中以新梢嫩芽部位的含量最高,老叶和茎梗中次之,花和果实中最少,因此咖啡碱通常作为茶叶老嫩程度和判断茶叶真伪的重要成分和指标之一[6]。此外,茶树的品种,生长环境,栽培条件也是影响茶树中咖啡碱含量的因素。20世纪80年代初发现了一种天然无咖啡碱茶叶——南昆山毛叶茶,其体内生物碱的组成以可可碱为主,而不是咖啡碱[7]。
咖啡碱不仅在植物体内能够保护植物免受自然环境的伤害,促进植物的生长,同时也是一种普遍、有一定药用价值的食物。适量的摄入咖啡碱可以健胃消食、利尿[8、9]、降低高血压[10]、提高记忆力、抗癌[11]、降低II型糖尿病风险[12、13]等;但过量的摄入会引起失眠[14]、焦虑[15]、骨质疏松[16]和胎儿畸形[17、18]等。因此,如何调控茶树中咖啡碱的合成对于茶树资源的高效利用有着重大意义。
2茶树中咖啡碱的合成途径
植物中咖啡碱合成途径大多是针对咖啡树和茶树的研究,但是在其他含有咖啡碱的植物中其合成的主要途径是一致的[6]。咖啡碱的嘌呤环骨架来源于嘌呤核苷酸,在嘌呤甲基化过程中其甲基供体主要来自于S-腺苷甲硫氨酸,它是由甲硫氨酸与ATP作用转化而来的。
`在茶树中咖啡碱的合成途径已经基本研究清晰:主要分为核心途径和供体途径。核心途径为腺嘌呤核苷酸途径,其包括四个主要步骤(如图中方框部分),(图1):经过N-7、N-3、N-1位的三次甲基化反应和一步脱核苷反应[19-20]。在为核心途径提供甲基的供体途径中,合成黄嘌呤核苷有多种来源,已发现的有四条[20]:腺嘌呤核苷酸(AMP) 途径,鸟嘌呤核苷酸(GMP) 途径,嘌呤环从头合成途径,S-腺苷甲硫氨酸循环(SAM)途径。其中,AMP途径中,由于AMP 大量存在于植物细胞中,AMP成为咖啡碱合成最有效的前体[1,19-25]。此外,在茶树植物体内还有其他合成咖啡碱的途径,如茶叶碱通过补救途径由3-甲基黄嘌呤、可可碱合成咖啡碱[26]。
3茶树中NMTs基因的克隆
由于植物中嘌呤生物碱拥有多样的代谢途径,在一定的条件下它们也会相互转化,因此,咖啡碱作为一种黄嘌呤生物碱化合物,在茶树合成咖啡碱的途径中,可能存在很多咖啡碱合成相关的NMTs基因。2000年,Kato[27]等从茶树嫩叶中得到咖啡碱合成酶,并利用RACE技术从茶叶中克隆得到了TCS1(AB031280),其是催化咖啡碱生物合成后两步甲基化反应的N(3、1)-甲基转移酶基因,并通过体外酶活实验证实了TCS1基因功能。根据TCS1基因序列,近年来从山茶属中克隆出多个NMTs基因,许煜华[28]等和金基强[29]等研究表明茶树中存在着与嘌呤碱合成相关的NMT基因家族,克隆出多个NMTS的gDNA序列。在NCBI中已登录的茶树咖啡碱合成相关的N-甲基转移酶序列有大概20多个,此外还有部分序列尚未登陆NCBI。

图1 茶树中咖啡碱的合成途径
随着近年来对茶树咖啡碱合成相关的N-甲基转移酶研究逐步深入,在茶树中克隆得到具有双重催化活性的NMTs,可以催化N-3和N-1的甲基化反应[27,30],同时在可可茶、滇缅茶等山茶属植物中克隆得到具有催化N-3甲基化反应的NMTs。然而,在茶树中催化咖啡碱合成的第一步甲基化的7-NMT未有报道,许多疑似具有N-7催化作用的NMTs序列,都证明没有功能。此外,茶树体内咖啡碱合成的主要途径已经研究透彻,但其次要代谢途径研究并不完整,其相关酶的研究也较少。
4茶树中NMTs的表达
咖啡碱一般在植物子叶、种子、幼叶中分布,在茶树中咖啡碱不仅分布在这些部位,其生物活性也已在山茶属植物的雄蕊、花瓣中发现,但是可可碱只发现在茶树的雄蕊中[31]。众多研究者通过实验证明,茶树中NMTs的表达量与咖啡碱的含量呈正相关,这一表达规律与咖啡相似,且茶树中参与咖啡碱生物合成的主要三个NMTs在嫩叶、成熟叶片和老叶中的表达量与咖啡大体是一致的[32]。幼嫩组织中的表达量高于成熟组织,但是在不含嘌呤生物碱的山茶属植物中NMTs在成熟叶片中的表达高于嫩叶[32,33]。通过研究者的实验,生长周期和季节的变化对茶树不同部位的NMTs的表达有一定的影响。Mohanpuria等[34]的研究发现茶树中咖啡碱的合成和降解是发展变化的和受季节性调节,与休眠生长阶段相比,咖啡碱合成酶在非休眠状态下在顶端芽,第一叶,第二叶,幼茎的表达量比老叶低。陈丽萍[35]等采用Northern blot技术证明NMTs在不同品种茶树中春、夏、秋3个季节都有表达,且同一季节不同品种之间NMTs的表达量无明显差异。李金[36]等研究表明在茶树秋季嫩叶中,TCS1表达量与咖啡碱含量显著正相关,即TCS1表达量显著影响咖啡碱的合成。Fujimori[37]等通过实验表明四月份到六月份可可碱和咖啡碱在茶树嫩芽中被检测,但是没有检测到咖啡碱合成酶的活性。
此外,研究者通过改变外界条件和不同的胁迫处理方式来研究茶树中是否受到其他因素的影响。Kato[38]等通过对离体茶树枝条(20-30 cm)进行低温处理、机械损伤和盐胁迫,发现对TCS基因表达没有影响;对整株茶树和离体的叶片都进行MeJA处理,TCS表达量都没有变化;用重金属处理茶树叶片24小时后抑制了TCS的表达,但这种抑制对TCS基因表达不是特异性的,因为它同时抑制了茶树咖啡碱合成其他相关基因的表达;茶树叶片脱水处理后24小时TCS表达消失。此外,在对茶树叶片分别进行蛋氨酸、谷氨酸和硝酸钠处理2小时后TCS表达量高于对照组(水处理后的叶片)两倍;在氯化铵和尿素处理后叶片中TCS表达水平略高于对照组(水处理后的叶片)。这极有可能是咖啡碱生物合成受下游氮代谢变化的调控,这就需要进一步明确咖啡碱代谢和氮代谢的关系。Shinozaki[39]等提出ABA缺少影响TCS的基因表达,表明ABA拥有独立的通路参与TCS基因表达。Koshiishi[40]等研究表明,在光处理和暗处理下的茶树叶片中都发生了咖啡碱生物合成且光照条件的改变对TCS基因的表达并没有影响。后者可能是由于咖啡碱合成酶的活性与叶绿体有关[41],因此光照条件的改变对TCS基因的表达并没有影响。综上所述,研究者对茶树中影响NMTs表达因素做了大量的研究,如不同季节、不同组织部位、不同外界条件和胁迫作用,使得研究者了解NMTs的表达规律,从而进一步促进NMTs的表达调控研究,对调节茶树中咖啡碱合成具有积极意义。
5茶树中NMTs的活性和结构
茶树咖啡碱合成酶的氨基酸序列及相关酶基因表现出高度的相似性且呈现出其底物特异性(见表1),将17条基因用MEGA软件做系统发育树分析(见图2)后发现它们的相似度都在65%以上。茶树中NMTs共有的高保守序列有:motif A(AA/VDLGCAAGP), motif B’(VYLNDLF/PGNDFN),motif C(PGSFHGRLFP)和YFFF(AYLSQFHEDFTMFL)。甲基供体结合位点(SAM)有五个关键氨基酸残基,这五个都有相同的motif A,B’和C保守区。实验证明TCS1-5的gDNA序列均由4个外显子和3个内含子组成[29]。三个保守区(motif A,B’和C)的编码区域位于TCS1序列的第二段外显子区。相对应的甲基黄嘌呤结合的氨基酸编码区(在TCS1氨基酸序列中225,269,和318的位置)位于第三和第四段外显子[42]。孔祥瑞[43]等采用生物信息学分析方法对植物咖啡碱合成酶的等电点、亚细胞定位、信号肽、跨膜螺旋、保守性功能结构域及基序、二级结构与三级结构等重要参数进行预测与分析。结果表明,植物咖啡碱合成酶主要定位于胞质和胞核中,含有磷酸化、酰基化和糖基化修饰位点。

图2 山茶属植物NMT蛋白序列的进化树

表1 茶树中NMTs的克隆及功能[5,43]
植物中参与咖啡碱生物合成相关的NMTs拥有不同的表达方式和转录水平,这些不同可能是由于多个原因导致的。2007年,Mccarthy[44]等实验表明黄嘌呤核苷甲基转移酶(XMT)中316位丝氨酸可能是底物黄嘌呤核苷的识别位点,XMT中161位谷氨酰胺到3,7-二甲黄嘌呤甲基转移酶(DXMT)中160位组氨酸可能是催化的结果,266位苯丙氨酸到异亮氨酸的变化则对其辨别单甲基和二甲基转移酶至关重要。
TCS1-6的核苷酸序列拥有很高的同源性(TCS6为假基因[29]),但是这些NMTs的启动子核苷酸序列的相似度非常低[45]。Jin[40,42]等对不同茶树品种5’-UTR区(ATG前252bp)碱基序列和其自然变异的等位基因TCS1a(TCS1,AB031280)(为对照组),TCS1b(ICS1,AB056108),TCS1c(PCS1, AB207817),TCS1d(KT215399),TCS1e(KT215397)和TCS1f(KT215398)进行研究。实验结果表明TCS1d,e,f(包含关键氨基酸第225位和第269位)三个基因重组蛋白的咖啡碱合成酶活性都高于可可碱合成酶,TCS1bc的重组蛋白只有可可碱合成酶(TS)的活性,说明TCS1的自然变异改变了其活性和转录水平。同时,通过对定点突变实验证明了TCS1 氨基酸残基269位对TCS的活性和底物识别中起着重要的作用,但不能检测底物的特异性[45]。此外,在低咖啡碱的茶种质资源中,TCS1等位基因转录水平低或其编码的蛋白质只有TS活性这两个分子机制控制着咖啡碱的生物合成,导致了咖啡碱的积累减少[45]。在不同的茶树品种TCS1是多样性的,这是个体样本不同的遗传背景的影响导致的。Jin[46]等对中国14个省选择的44个拥有丰富遗传背景的茶树品种中,在其TCS1的外显子区发现了31个SNPs。此外,对TCS1中的SNP4318进行位点突变(C/T)可显著提高TCS1重组蛋白后可可碱合成酶(TS)和咖啡碱合成酶(CS)的活性,同时验证了SNP4318和咖啡碱含量的关系。
综上所述,众多研究者希望明确决定NMTs活性的关键位点。目前大多数研究者用定点突变的方法来进行功能验证(表2)。NMTs的结构与其活性关系的研究,将有助于我们对咖啡碱合成酶分子机制的明晰及高、低咖啡碱茶树选育具有重要意义。
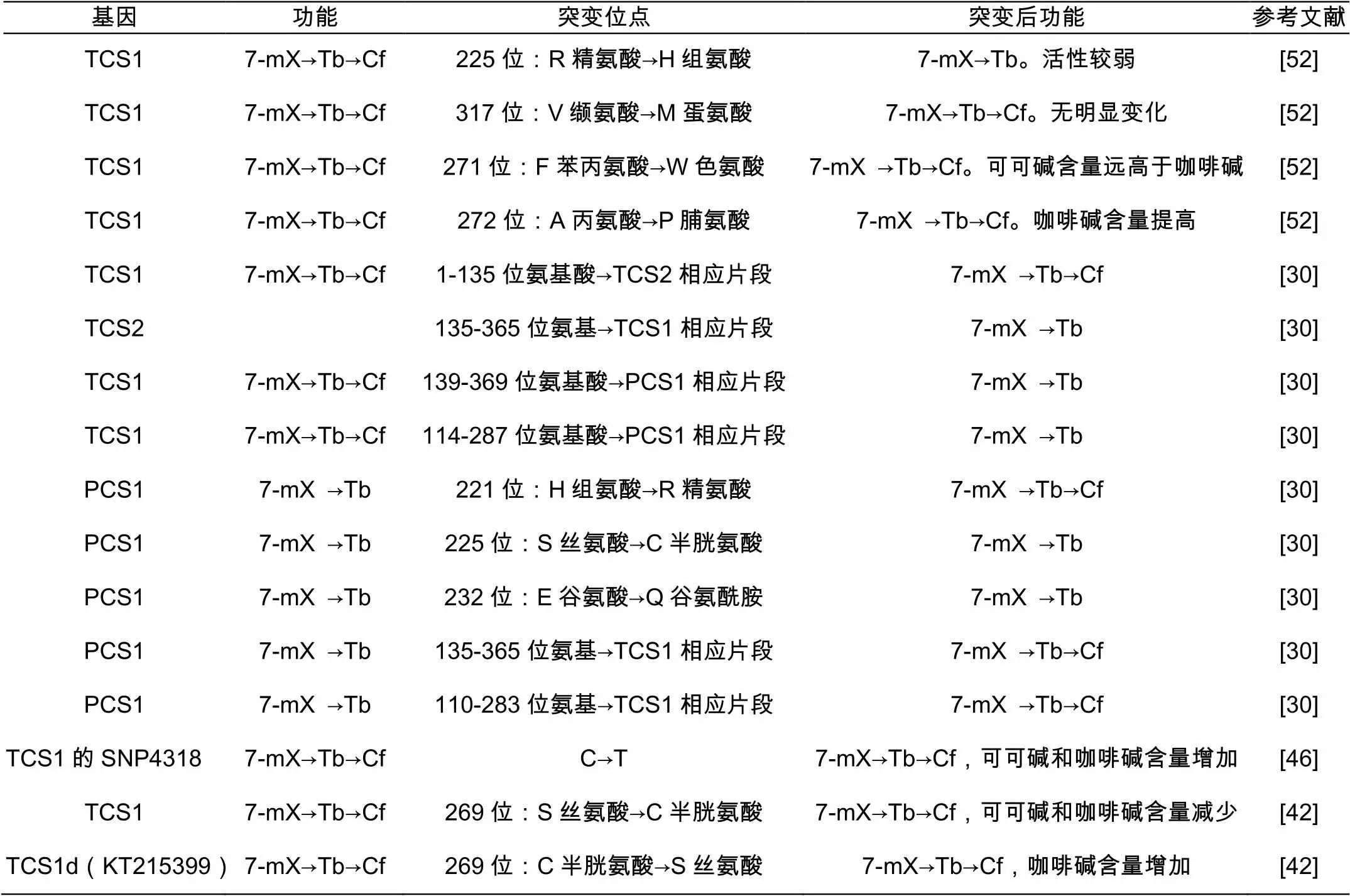
表2 定点突变研究NMTs的活性位点[5]
6展望
由于茶树NMTs直接参与咖啡碱合成茶树中与咖啡碱合成相关的NMTs基因的克隆、调控对培育高或低的咖啡碱茶树品种有重要意义。余有本[47]等通过构建TCS1的植物表达载体,将TCS1转入烟草发现TCS1在这些植株中均能表达。同时,通过提取转基因植株中粗酶液进行体外酶促反应发现,TCS1能催化7-甲基黄嘌呤和可可碱生成咖啡碱,由此证明转基因烟草植株中的TCSl具有催化活性。此外,张广辉[48]等构建了TCS基因的RNA干涉载体,从而降低茶树中的咖啡碱含量;一些研究者通过反义RNA等手段获得低咖啡碱含量的茶树品种。2016年,Jin[46]等通过分子标记辅助选择(MAS)的方法为选择高含量的咖啡碱茶树品种提供了强大高效分子标记实验基础,为加快育种进程、提高效率和选择精确提供策略。
根据已有报道,目前茶树中NMTs的研究需要进一步深入,要进一步完善茶树中NMT信息,了解NMTs在茶树中的调控机制,为完善咖啡碱合成代谢途径提供分子证据,从而实现茶树咖啡碱的有效靶标。
[1] Ashihara H, Sano H, Crozier A. ChemInform Abstract: Caffeine and Related Purine Alkaloids: Biosynthesis, Catabolism, Function and Genetic Engineering [J]. Phytochemistry, 2008, 69(4):841-856.
[2] Ashihara H, Gillies F M, Crozier A. Metabolism of Caffeine and Related Purine Alkaloids in Leaves of Tea (L.)[J]. Plant & Cell Physiology, 1997, 38(4):413-419.
[3] Crozier A. Biosynthesis of Caffeine in Leaves of Coffee [J]. Plant Physiology, 1996, 111(3):747-753.
[4] Ashihara H. Purine metabolism and the biosynthesis of caffeine in maté leaves [J]. Phytochemistry, 1993, 33(6):1427-1430.
[5] 晏嫦妤, 任秋婧, 陈小芳,等. 咖啡碱合成N-甲基转移酶研究进展[J]. 茶叶科学, 2014(6):531-540.
[6] 宛晓春,夏涛,等.茶树次生代谢[M]. 北京:科学出版社,2015,5:5-7.
[7] 李斌, 郑永球, 尹逸,等. 天然无咖啡碱茶叶资源的开发利用研究[J]. 食品科学, 2001, 22(7):33-35.
[8] Pare W. The effect of caffeine and seconal on a visual discrimination task [J]. Journal of Comparative & Physiological Psychology, 1961, 54(5):506-509.
[9] Brunyé T T, Mahoney C R, Lieberman H R, et al. Caffeine modulates attention network function [J]. Brain & Cognition, 2009, 72(2):181-8.
[10] Hartley T R, Sung B H, Pincomb G A, et al. Hypertension risk status and effect of caffeine on blood pressure.[J]. Hypertension, 2000, 36(1):137-41.
[11] Bode A M, Dong Z. The enigmatic effects of caffeine in cell cycle and cancer [J]. Cancer Letters, 2007, 247(1):26-39.
[12] 易超然, 卫中庆. 咖啡因的药理作用和应用[J]. 医学研究生学报, 2005, 18(3):270-272.
[13] Mackenzie T, Comi R, Sluss P, et al. Metabolic and hormonal effects of caffeine: randomized, double-blind, placebo-controlled crossover trial [J]. Metabolism-clinical & Experimental, 2007, 56(12):1694-1698.
[14] Shilo L, Sabbah H, Hadari R, et al. The effects of coffee consumption on sleep and melatonin secretion [J]. Sleep Medicine, 2002, 3(3):271-273.
[15] Robertson D, Frölich J C, Carr R K, et al. Effects of Caffeine on Plasma Renin Activity, Catecholamines and Blood Pressure [J]. New England Journal of Medicine, 1978, 298(4):181-6.
[16] Montes F, Cabrera M A, Salgar C, et al. The role of potassium channels in the vasodilatory effect of caffeine in human internal mammary arteries [J]. Vascular Pharmacology, 2008, 50(3-4):132-6.
[17] Weng X, Odouli R, Li D K. Maternal caffeine consumption during pregnancy and the risk of miscarriage: a prospective cohort study [J]. American Journal of Obstetrics & Gynecology, 2008, 198(3):1-8.
[18] Heaney R P, Recker R R. Effects of nitrogen, phosphorus, and caffeine on calcium balance in women [J]. Journal of Laboratory & Clinical Medicine, 1982, 99(1):46-55.
[19] Kato M, Mizuno K, Fujimura T, et al. Purification and characterization of caffeine synthase from tea leaves [J]. Plant Physiology, 1999, 120(2):579-86.
[20] Suzuki T, Ashihara H, Waller G R. Purine and purine alkaloid metabolism in, andplants [J]. Phytochemistry, 1992, 31(8):2575-2584.
[21] Negishi O, Ozawa T, Imagawa H. Biosynthesis of Caffeine from Purine Nucleotides in Tea Plant [J]. Bioscience Biotechnology & Biochemistry, 1992, 56(3):499-503.
[22] Stasolla C, Katahira R, Thorpe T A, et al. Purine and pyrimidine nucleotide metabolism in higher plants [J]. Journal of Plant Physiology, 2003, 160(11):1271-95.
[23] Negishi O, Ozawa T, Imagawa H. Guanosine Deaminase and Guanine Deaminase from Tea Leaves [J]. Bioscience Biotechnology & Biochemistry, 2014, 58(7):1277-1281.
[25] Keya C A, Crozier A, Ashihara H. Inhibition of caffeine biosynthesis in tea () and coffee () plants by ribavirin [J]. Febs Letters, 2003, 554(3):473-7.
[26]谢果, 何蓉蓉, 栗原博. 茶叶生物碱的生物合成与代谢的研究进展[J]. Chinese Journal of Natural Medicines, 2010, 8(2):153-160.
[27] Kato M, Mizuno K, Crozier A, et al. Plant biotechnology: Caffeine synthase gene from tea leaves [J]. Nature, 2000(6799):956-957.
[28] 许煜华, 文海涛, 赵亮,等. 英红九号cDNA文库的构建及NMT基因的筛选[C]// 中国农业工程学会农产品加工及贮藏工程分会学术年会暨华南地区农产品加工产学研研讨会. 2010.
[29] 金基强, 姚明哲, 马春雷,等. 合成茶树咖啡碱相关的N-甲基转移酶基因家族的克隆及序列分析[J]. 茶叶科学, 2014(2):188-194.
[30]Yoneyama N, Morimoto H, Ye C X, et al. Substrate specificity of N-methyltransferase involved in purine alkaloids synthesis is dependent upon one amino acid residue of the enzyme [J]. Molecular Genetics & Genomics, 2006, 275(275):125-35.
[31] Fujimori N, Ashihara H. Adenine metabolism and the synthesis of purine alkaloids in flowers of[J]. Phytochemistry, 1990, 29(11):3513-3516.
[32] 李叶云, 芦原坦. 茶树咖啡碱生物合成相关基因表达研究[C]// 2008茶学青年科学家论坛论文集. 2008.
当转发模式为GSF时,雷区中心位置和比例因子均为空。当转发模式为GSF时,需利用雷区转发数据包,进而平衡网络负载。因此,对凸包H进行相似变换,且从凸包H内随机选择中心位置,而比例因子ξ的定义如式(6)所示:
[33] Ishida M, Kitao N, Mizuno K, et al. Occurrence of theobromine synthase genes in purine alkaloid-free species of, plants [J]. Planta, 2009, 229(3):559-568.
[34]Mohanpuria P, Kumar V, Joshi R, et al. Caffeine biosynthesis and degradation in tea [Camellia sinensis (L.) O. Kuntze] is under developmental and seasonal regulation.[J]. Molecular Biotechnology, 2009, 43(2):104-11.
[35]陈丽萍. 南昆山毛叶茶等山茶属植物N-甲基转移酶基因克隆与表达研究[D]. 华南农业大学, 2006.
[36]李金, 魏艳丽, 庞磊,等. 茶树咖啡碱合成途径中TCS1、TIDH、SAMS的基因表达量差异及其与咖啡碱含量的相关性[J]. 江苏农业科学, 2013(10):21-24.
[37] Fujimori N, Suzuki T, Ashihara H. Seasonal variations in biosynthetic capacity for the synthesis of caffeine in tea leaves [J]. Phytochemistry, 1991, 30(7):2245-2248.
[38] Kato M, Kitao N, Ishida M, et al. Expression for Caffeine Biosynthesis and Related Enzymes in[J]. Zeitschrift Fur Naturforschung C, 2010, 65(4):245-256.
[39] Shinozaki K, Yamaguchi-Shinozaki K. Molecular responses to dehydration and low temperature: differences and cross-talk between two stress signaling pathways [J]. Current Opinion in Plant Biology, 2000, 3(3):217-223.
[40] Koshiishi C, Ito E, Kato A, et al. Purine Alkaloid Biosynthesis in Young Leaves of, in Light and Darkness[J]. Journal of Plant Research, 2000, 113(2):217-221.
[41] Kato A, Crozier A, Ashihara H. Subcellular localization of the-3 methyltransferase involved in caffeine biosynthesis in tea [J]. Phytochemistry, 1998, 48(5):777-779.
[42] Jin J Q, Yao M Z, Ma C L, et al. Natural allelic variations of TCS1 play a crucial role in caffeine biosynthesis of tea plant and its related species [J]. Plant Physiology & Biochemistry, 2016, 100:18-26.
[43]孔祥瑞, 杨军, 王让剑. 植物咖啡碱合成酶的生物信息学分析[J]. 福建农业学报, 2014(12):1211-1218.
[44] Mccarthy A A, Mccarthy J G. The structure of two-methyltransferases from the caffeine biosynthetic pathway [J]. Plant Physiology, 2007, 144(2):879-89.
[45] Jin J, Yao M, Chunlei M A, et al. Cloning and Sequence Analysis of the-methyltransferase Gene Family Involving in Caffeine Biosynthesis of Tea Plant[J]. Journal of Tea Science, 2014, 34(2):188-194.
[46] Jin J Q, Yao M Z, Ma C L, et al. Association mapping of caffeine content with TCS1, in tea plant and its related specie [J]. Plant Physiology & Biochemistry, 2016, 105:251-259.
[47] 余有本, 江昌俊, 宛晓春,等. 茶树咖啡碱合成酶基因cDNA在烟草中的表达[J]. 西北农林科技大学学报:自然科学版, 2007, 35(11):181-186.
[48] 张广辉, 梁月荣, 陆建良,等. 茶树咖啡因合成酶基因RNA干涉表达载体构建[J]. 茶叶科学, 2006, 26(4):243-248.
[49] Camellia sinensis TCSz Mrna for caffeine synthase, complete cds [DB]. http://www.ncbi.nlm.nih.gov/nuccore/AB031281.
[50] Camellia sinensis caffeine synthase mRNA, complete cds[DB]. http://www.ncbi.nlm.nih.gov/ nuccore/AY907710.
[51] Camellia sinensis caffeine synthase gene, complete cds[DB]. http://www.ncbi.nlm.nih.gov/nuccore/No. EF526217.
[52] 李萌萌, 邓威威, 金璐,等. 茶树咖啡碱合成酶基因TCS1的定点突变及体外表达分析[C]// 中国科协年会——分12茶学青年科学家论坛. 2014.
(责任编辑:蒋文倩)
2016-09-06
S571.1
A
1006-5768(2016)04-0177-08
