湖北木林子自然保护区大样地小叶青冈种群年龄结构及点格局分析
2016-10-12艾训儒张金龙
黄 伟,艾训儒*,黄 升,2,林 勇,陈 斯,张金龙
(1.湖北民族学院林学园艺学院,湖北恩施445000;2.恩施冬升植物开发有限公司,湖北恩施445000)
湖北木林子自然保护区大样地小叶青冈种群年龄结构及点格局分析
黄 伟1,艾训儒1*,黄 升1,2,林 勇1,陈 斯1,张金龙1
(1.湖北民族学院林学园艺学院,湖北恩施445000;2.恩施冬升植物开发有限公司,湖北恩施445000)
小叶青冈是我国亚热带常绿落叶阔叶混交林的主要优势树种之一,也是良好的用材树种和园林树种.本文采用胸径代替年龄和点格局分析(Rip1ey's K-Function)法对湖北木林子15 hm2大样地内小叶青冈种群的年龄结构和空间点格局进行了分析.结果显示:大样地内小叶青冈种群属增长型种群,其种群的存活曲线特征属DeeveyⅡ型,种群在幼树阶段死亡率较高,之后趋于稳定;种群整体及幼树、小树、中树和大树四个年龄段的个体在空间分布格局上随研究尺度的变化均呈集群分布,而老树年龄段的个体,其空间分布格局随研究尺度的增加由均匀分布向集群分布递变;在呈集群分布的尺度范围内,都存在随研究尺度的增加,聚集强度先减小后增大的规律.
小叶青冈;种群;年龄结构;点格局;生命表
种群是指分布在一定时空范围内的同种生物个体的集合[1],年龄结构是种群结构的重要组成部分,与种群的数量动态息息相关[2],分布格局是种群内个体在一定时空范围内的分布方式与配置特点[3],它呈现了特定时空下种群内不同个体之间以及不同种群个体之间、种群与环境之间的相互作用结果[4-5],采用不受研究尺度限制的点格局分析法对种群个体分布格局进行分析,有助于揭示种群的成因及群落生物多样性的维持机制.
小叶青冈(Cyclobalanopsis gracilis)为壳斗科青冈属的常绿阔叶大乔木,是我国亚热带常绿阔叶林和常绿落叶阔叶混交林等植被类型的主要优势树种之一[6],分布海拔一般在500~2600 m之间[7].其树皮灰褐色,幼枝被绒毛,新叶红色,树干通直,树冠整齐,树形优美,是良好的园林绿化树种,其材质坚重、耐腐、耐磨,为纺织工业、建筑业、交通运输业的上等用材,也是食用菌生产的优质用材[8].进行解释:
1 研究区域概况
木林子常绿落叶阔叶混交林大样地位于湖北木林子国家级自然保护区内,该保护区位于湖北省西南部恩施州鹤峰县境内,为云贵高原的东北延伸部分武陵山脉东北端石门支脉,地理坐标29°55'59″~30°10'47″N,109°59'30″~110°17'58″E,海拔范围1100~2096m.保护区面积20838 hm2,属亚热带湿润季风气候,年平均气温15.5℃,绝对最高温和最低温分别为39℃和-17.1℃,多年平均降水量1 733.7 mm,年均相对湿度82%,全年无霜期270~279d;山地黄壤、山地黄棕壤和山地棕壤为其主要自然土壤,800~1500 m为山地黄棕壤,1500m以上为棕壤,成土母质为绿色砂页岩、紫色砂页岩和泥质页岩.
2 研究方法
2.1 样地设置与调查
2013年6-9月,在木林子国家级自然保护区核心区内,选择地势相对平缓、内部地形相对一致的区域,按照热带林业科学中心(CTFS)的标准和技术规范,采用实时动态测量仪(RTK)从样地原点沿东西方向和南北方向每隔20m定点建立东西长300m、南北长500m的15hm2固定监测样地.将15 hm2大样地划分为375 块20m×20m的样地,样地4角用经过防腐处理的不锈钢管作永久标记,在每块20 m×20 m的样地内用插值法细分为4个10m×10m的样方共1500个和16个5m×5m的小样方共6000个.
在对样地生境因子(坡度、坡向、坡位、土壤、样地中心点经纬度和海拔等)调查的基础上,对样地内所有胸径(DBH)≥1.0cm的木本植物个体在胸径1.3 m处用红色油漆标记,对所有标记的植物个体用铜丝(5.0 cm≥DBH≥1.0cm个体)或镙纹钢钉(DBH>5.0cm个体)套挂制有唯一编号的特制铝牌.以样地为基本单元,对已标记挂牌的所有植物个体进行测树因子及坐标检测,主要包括物种登记与鉴别、胸径、树高以及在样地的坐标值(以样地西南角为原点,测定x轴和y轴).
2.2 种群年龄结构分析
2.2.1 种群径级结构划分 采用“径级代替龄级”[4,6,9-12]并参考相关径级结构的研究方法[9-11],结合大样地小叶青冈种群的实际生长情况,将其胸径划分为15个等级,根据径级结构划分将小叶青冈划分为5个年龄段并统计种群个体数.见表1.

表1 小叶青冈种群径级及年龄结构Tab.1 The DBH c1ass structure and age structure ofCyclobalanopsis gracilis popu1ation
2.2.2 种群静态生命表编制 统计各龄级小叶青冈的个体数,根据生命表各参数值编制种群静态生命表;以径级为横坐标,死亡率和标准化的存活个体数的自然对数为纵坐标,绘制死亡曲线[13-15]和存活曲线.
2.3 种群空间分布格局分析
采用点格局的分析方法,对种群整体及不同龄级在不同尺度下的空间分布格局进行分析.点格局分析的基本原理可用函数

式中:A为样地面积;n为总点数(植物个体数);uij为点i和点j之间的距离;r为研究尺度;当uij≤r时,Ir(uij)= 1,当uij>r时,Ir(uij)=0;Wij为边界影响修正参数.通常采用有直观线型效果的L(r)代替函数,其表达式为:
点格局分析中,先用Exce1对数据进行基础分类处理,然后用R软件的spatstat程序包中的L-function进行数据分析,分析返回值包括Lobs(r)和Ltheo(r),其中Lobs(r)为实际分析值,Ltheo(r)为完全随机分布下的理论值,两者进行比较:Lobs(r)位于Ltheo(r)左侧,表示集群分布;两者相交或重合,表示随机分布;Lobs(r)位于Ltheo(r)右侧表示均匀分布.
用Monte-Car1o拟合检验计算上下包迹线,拟合次数为100次,得99%的置信区间,若Lobs(r)在包迹线以内,则符合随机分布,若在上包迹线左侧,为集群分布;若在下包迹线右侧,为均匀分布.
3 结果与分析
3.1 种群径级结构
小叶青冈种群的年龄结构和径级结构见图1、图2.

图1 小叶青冈种群的年龄结构Fig.1 The age structure of Cyclobalanopsis gracilis popu1ation

图2 小叶青冈种群的径级结构Fig.2 The DBH c1ass structure of Cyclobalanopsis gracilis popu1ation
由表1和图1、图2可知,木林子大样地小叶青冈种群幼树、小树、中树、大树和老树的个体数依次为3555株、2008株、968株、305株和14株,分别占种群总个体数的51.9%、29.3%、14.1%、4.5%和0.2%.可见,木林子大样地小叶青冈种群幼树补给十分充足,老树个体相对较少,种群的出生率大于死亡率,为典型的增长型种群;由表1和图2知,木林子大样地小叶青冈种群径级结构中前两径级的个体数总数占种群总个体数的81.21%,这有助于整个种群在今后很长一段时间内的结构稳定性的维持.
3.2 种群静态生命表
由于木林子大样地小叶青冈种群径级结构中的第13-15级的个体数很少,仅3株(见表1),为满足生命表的编制要求,将此三个径级合并,编制静态生命表(见表2).
由表2知,小叶青冈种群的相邻龄级间隔期间的平均存活数(Lx)随年龄增加呈依次下降趋势,符合种群的一般生物学特性;小叶青冈种群的期望寿命(ex)在第3级达到最大值2.412,在最后一级降到最低值,总体上呈现出在一定范围内上下波动的规律;通过对比小叶青冈种群的期望寿命(ex)与死亡率(qx)的变化规律可知,两者都在一定范围内波动,且波动趋势刚好相反,即前者的上升(或下降)对应着后者的下降(或上升).因此,小叶青冈种群上一龄级的死亡率在一定程度上决定其种群在下一龄级的期望寿命,即种群通过自疏上一龄级的个体数量,为存活的下一龄级的个体留出生存空间,种群自疏[17]成为该种群应对生存资源限制的主要对策之一.
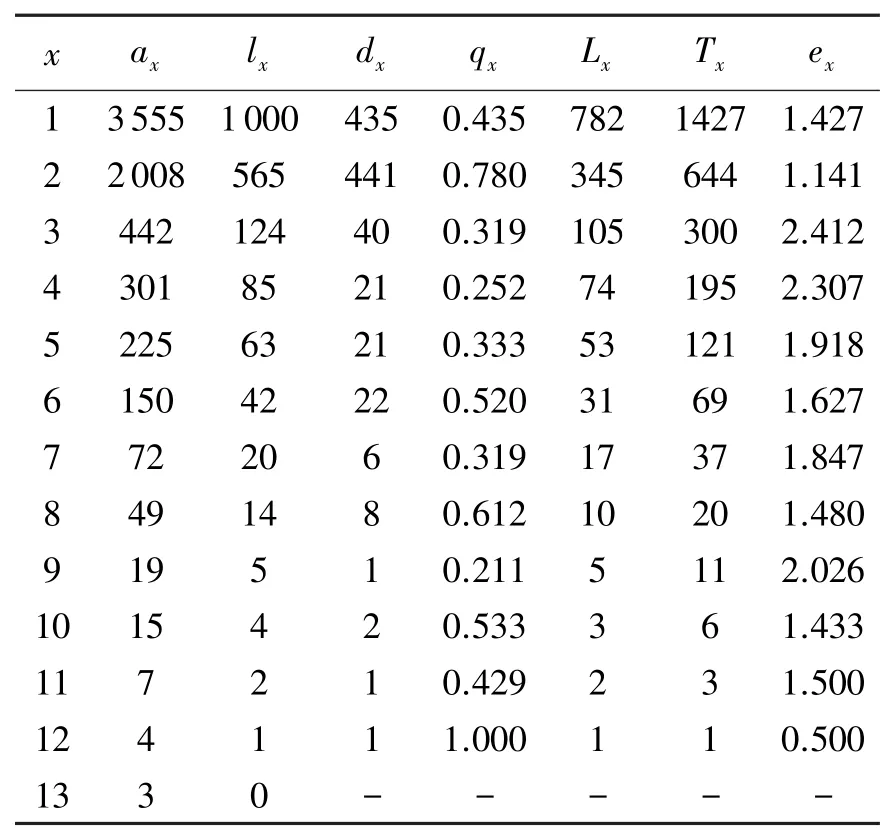
表2 小叶青冈种群的静态生命表Tab.2 Life tab1e of Cyclobalanopsis gracilis popu1ation in Wu1ashan Nature Reserve
3.3 种群死亡曲线与存活曲线特征分析
根据小叶青冈种群的静态生命表,可绘制死亡曲线(见图3)和存活曲线(见图4).
由图3可知,小叶青冈种群的死亡曲线整体呈现出“多峰值起伏状”特征.小树的死亡率较高,达80%;第2~4级,种群的死亡率持续下降,下降趋势逐渐减弱;自第5级后,种群的死亡曲线进入稳定状态.由于小叶青冈幼树生长旺盛[18],在短时间内大量消耗了生境内有限的资源,致使种内竞争急剧上升,为保证其种群的延续,种群经历了一次较高程度的自疏过程[19],为种群在第3~4级的生长预留了足够的空间,故在第2~4级,种群的死亡率持续下降;自第5级开始,小叶青冈种群应对生长过程中资源限制的自疏机制渐趋成熟,所以死亡率趋于稳定.由图4可知,小叶青冈种群的存活特征曲线呈对角线型,属于DeeveyⅡ[20],表明小叶青冈种群在其整个生活期内具有较稳定的存活率,种群结构较稳定.在当前生境条件不发生急剧变化的情况下,木林子小叶青冈种群结构能在很长一段时间内维持稳定.

图3 小叶青冈种群的死亡曲线Fig.4 Morta1ity ratio curve of Cyclobalanopsis gracilis popu1ation

图4 小叶青冈种群的存活曲线Fig.4 Surviva1 curve of Cyclobalanopsis gracilis popu1ation
3.4 种群点格局分析
小叶青冈种群整体和不同年龄段个体在木林子大样地的分布见图5至图10,其空间点格局分布见图11至图16.
由图5至图10可知:在木林子大样地(300m×500m)内,无论是小叶青冈种群个体的分布点图,还是不同年龄段个体的分布点图,都呈明显的集群分布,且分布趋势一致,但仅凭该图并不能看出空间分布格局与研究尺度的关系.在实际调查中发现,小叶青冈种群个体及不同年龄段个体在大样地的山谷、阴坡等地的分布密度明显较高,这主要与小叶青冈喜山谷、阴坡生境的生长习性相关[21].有研究表明,小叶青冈在群落中具有相对较宽的生态位[22],因此其种群个体在大样地内均有分布,只是分布密度存在大小差异.从幼树到老树,不同阶段的小叶青冈个体呈现出相同的分布趋势,表明小叶青冈种群同一年龄段内的种内竞争程度在样地内各处差异不大,即其种群会尽可能地利用生境内资源保证其种群的延续[22].木林子大样地生境的不均一性和小叶青冈种群与其环境适应对策的共同作用,使小叶青冈种群在木林子大样地内呈集群分布.
通过对小叶青冈种群个体及不同年龄段个体的空间点格局分析(图11至图16)可知,除老树的空间分布随研究尺度的增加呈现一定的变化外,其他年龄段的种群个体随研究尺度的增加均呈集群分布.小叶青冈种群整体和幼树、小树、中树、大树四个年龄段的种群个体,其空间分布格局基本一致,在10 m以下(0<r<10 m)的空间尺度范围内,聚集强度缓慢减小后缓慢增大,在10 m以上(r>10 m)的空间尺度内时,聚集强度的变化不是很明显.老树的点格局分析显示:在7m以下(r<7m)的空间尺度范围内,种群呈均匀分布,而在7 m以上(r>7 m)的空间尺度范围内呈集群分布,聚集强度也呈现出先下降后上升的趋势,这与种群整体分布格局的趋势相一致.
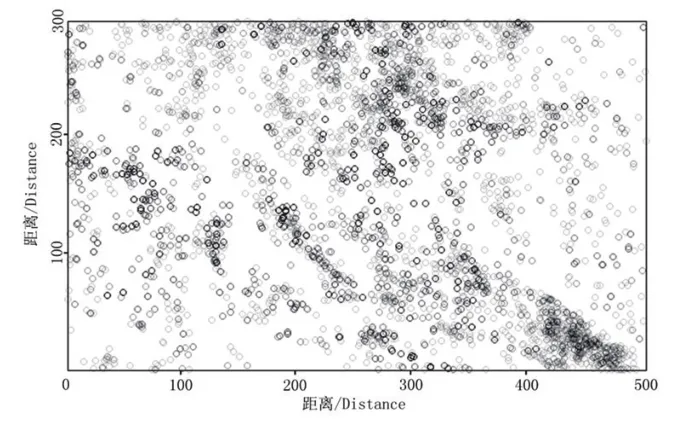
图5 小叶青冈种群空间分布点图Fig.5 Mapped point pattern of Cyclobalanopsis gracilis popu1ation

图6 小叶青冈幼树空间分布点图Fig.6 Mapped point pattern of the sap1ings

图7 小叶青冈小树空间分布点图Fig.7 Mapped point pattern of the young trees
4 结论与讨论
通过对木林子小叶青冈种群的年龄结构、静态生命表、死亡曲线和存活曲线的分析可知,木林子小叶青冈种群幼树的死亡率较高,但幼树的补给十分充足,该种群为增长型种群,且种群结构稳定;“自疏现象”是木林子小叶青冈种群应对有限的环境资源的主要对策.杨同辉[23]在对浙江天童山小叶青冈种群的研究也发现其种群幼树死亡率很高,且存在明显的低龄缺损现象,但种群结构依然稳定,只是为非增长型种群,故幼苗的补给与更新是小叶青冈种群得以稳定和延续的基础[24].在自然条件下,小叶青冈种群存在隔年结实的现象,该现象是否是小叶青冈种群的死亡曲线整体呈“多峰值起伏状”特征的原因之一,还有待进一步研究.
通过对木林子小叶青冈种群在大样地内的分布研究发现,其种群个体及不同年龄段的个体在大样地内均呈集群分布,且分布趋势一致,这是种群的环境适应性对策和所处生境不均一性的综合结果,生境的不均一性是该过程的外因,种群的生物学习性是内因,环境适应性对策是各年龄段分布趋势相一致的关键;通过小叶青冈种群的空间点格局分析发现,除老树个体群外,其种群个体及其他各年龄段个体群随研究尺度增加均呈集群分布,因此,种内竞争并非此空间格局形成的主导因子,但该过程的主导因子是源自环境的非生物因素还是小叶青冈种群所处群落的生物因素[25],还有待于进一步研究.

图8 小叶青冈中树空间分布点图Fig.8 Mapped point pattern of the midd1e aged-trees
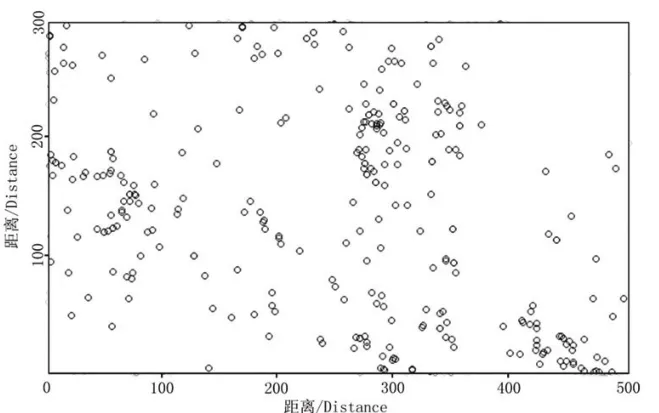
图9 小叶青冈大树空间分布点图Fig.9 Mapped point pattern of the mature trees

图10 小叶青冈老树空间分布点图Fig.10 Mapped point pattern of the o1d trees

图11 小叶青冈种群空间分布格局Fig.11 Spatia1 pattern of Cyclobalanopsis gracilis

图12 小叶青冈幼树空间分布格局Fig.12 Spatia1 pattern of the sap1ings

图13 小叶青冈小树空间分布格局Fig.13 Spatia1 pattern of the young trees
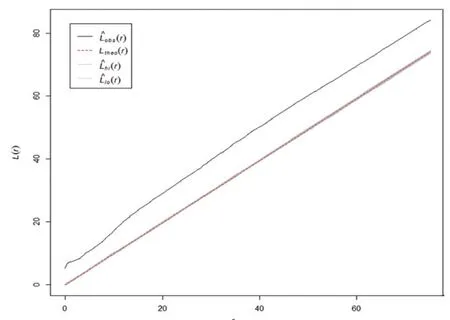
图14 小叶青冈中树空间分布格局Fig.14 Spatia1 pattern of the midd1e aged-trees

图15 小叶青冈大树空间分布格局Fig.15 Spatia1 pattern of the mature trees

Fig.16 Spatia1 pattern of the o1d trees图16 小叶青冈老树空间分布格局
对种群的年龄结构、数量动态和空间点格局的研究,有助于揭示种群的结构及其结构稳定性的维持机制和种群当前的空间分布格局及其格局成因,但种群的延续与更替,是以其无数种群个体相继完成其生活史[26]的过程为基础的,因此该过程具有一定的无限连续重复性,故在分析种群空间分布格局成因时,无法确定形成当前空间分布格局的原始起点,即究竟是原有母树的空间分布决定了种子的空间分布[27],进而决定了幼苗的空间分布格局,最终为整个种群的空间部分格局奠定基调,还是植物种子通过多种散布和传播方式[28],在一定程度上将种子的分布进行了均匀化和随机化后,环境变量[29]成为空间分布格局形成的初始主导因子,这有待于更大时空跨度的研究来揭示.
[1] 牛翠娟,娄安如,孙儒泳,等.基础生态学[M].2版.北京:高等教育出版社,2007:66-66.
[2] 李海东.雅鲁藏布江中游河岸带几种主要沙生植物种群点格局分析[J].植物生态学报,2011,35(8):834-843.
[3] YANG H X,ZHANG J T,WU B.Point pattern ana1ysis of Artemisia ordosica popu1ation in the Mu Us sandy 1and.Chinese Journa1 of P1ant Eco1ogy,2006,30(4):563-570.
[4] 李立.古田山常绿阔叶林优势树种甜槠和木荷的空间格局分析[J].植物生态学报,2010,34(3):241-252.
[5] 克平.试论生物多样性的概念[J].生物多样性,1993,1(1):20-22.
[6] 杨同辉.浙江天童小叶青冈种群结构和动态研究[J].福建林业科技,2010,37(1):26-29.
[7] 刘彬彬.浙江省小叶青冈生长过程的研究[J].浙江农林大学学报,2013,30(4):517-522.
[8] 王良衍.浙江天童山小叶青冈种群种子雨和地表种子库动态研究[J].福建林业科技,2011,38(3):8-10.
[9] 胡尔查.乌拉山自然保护区白桦种群的年龄结构和点格局分析[J].生态学报,2013,33(9):2867-2876.
[10] 张文辉.濒危植物太白红杉种群年龄结构及其时间序列预测分析[J].生物多样性,2004,12(3):361-369.
[11] 高贤明.北京山区辽东栎林的径级结构、种群起源及生态学意义[J].植物生态学报,2001,25(6)673-678.
[12] 韩路.不同生境胡杨种群径级结构与格局动态研究[J].北京林业大学学报,2010,32(1):7-12.
[13] 李豪.历山自然保护区秃山白树天然种群生命表[J].生态学杂志,2013,32(1):52-58.
[14] 张志祥.九龙山珍稀濒危植物南方铁杉种群数量动态[J].植物生态学报,2008,32(5):1146-1156.
[15] 陈玉华.篌竹无性系种群生态学特征研究[D].南京:南京林业大学,2004.
[16] 张金屯.植物种群空间分布的点格局分析[J].植物生态学报,1998,22(4):344-349.
[17] 黎磊.植物种群自疏过程中构件生物量与密度的关系[J].生态学报,2012,32(13):3987-3997.
[18] 刘彬彬.浙江省小叶青冈生长过程的研究[J].浙江农林大学学报,2013,30(4):517-522.
[19] REYNOLDS J H,FORD E D.Improving competition representation in theoretica1 mode1s of se1f-thinning:a critica1 review.Journa1 of Eco1ogy,2005,93:362-372.
[20] 牛翠娟,娄安如,孙儒泳,等.基础生态学[M].2版.北京:高等教育出版社,2007:109-109.
[21] 陈焕庸,黄成就.中国植物志第22卷[M].北京:科学出版社,1998:325-325.
[22] 林芳.小叶青冈群落主要种群生态位研究[J].北华大学学报,2014,15(5):684-688.
[23] 杨同辉.浙江天童小叶青冈种群结构和动态研究[J].福建林业科技,2010,37(1):26-29.
[24] 王良衍.浙江天童山小叶青冈种群种子雨和地表种子库动态研究[J].福建林业科技,2011,38(3):8-10.
[25] 牛翠娟,娄安如,孙儒泳,等.基础生态学[M].2版.北京:高等教育出版社,2007:6-75.
[26] 牛翠娟,娄安如,孙儒泳,等.基础生态学[M].2版.北京:高等教育出版社,2007:109-118.
[27] 杜彦君.森林种子雨研究进展与展望[J].生物多样性,2012,20(1):94-107.
[28] CLARK J S,FASTIE C,HURTT G,et a1.Reid's paradox of rapid p1ant migration[J].BioScience,1998a,48:13-24.
[29] 牛克昌.群落构建的中性理论和生态位理论[J].生物多样性,2009,17(6):579-593.
责任编辑:高 山
Age Structure and Point Pattern of Cyclobalanopsis Gracilis PoPulation in Mulinzi Nature Reserve of Hubei Province
HUANG Wei1,AI Xunru1*,HUANG Sheng1,2,LIN Yong1,CHEN Si1,ZHANG Jin1ong1
(1.Schoo1 of Forestry and Horticu1ture,Hubei University for Nationa1ities,Enshi 445000,China;2.Enshi Dongsheng P1ant Deve1opment Co.Ltd.,Enshi,445000,China)
Cyclobalanopsis gracilis is one of the dominant species of the subtropica1 evergreen and deciduous broad-1eaved mixed forest community in our country,and it is a1so a good timber species and 1andscape tree.This artic1e emp1oys the method of tree diameter serving as a proxy of tree age and point pattren ana1ysis to research the age structure and point pattern of the Cyclobalanopsis gracilis popu1ation of 15 hm2forest dynamics p1ot(300m×500m)in Mu1inzi Nature Reserve of Hubei Province.The resu1ts indicate:The Cyclobalanopsis gracilis popu1ation in the forest dynamics p1ot fo11ows an expanding popu1ation,and its survivorship curve fo11ows a DeeveyⅡindecating the morta1ity of this popu1ation is high first and then fo11ows stationary;the point pattern of the Cyclobalanopsis gracilis popu1ation,the sap1ings,the young trees,the midd1e aged-trees and the mature trees are of an aggregated distribution with the research sca1e,but the o1d trees'changes from a regu1ar pattern to an aggregated distribution;the aggregation intensity decreases first and then increases in a11 aggregated distribution.
Cyclobalanopsis gracilis;popu1ations;age structure;point parttern;1ife tab1e
Q145.1
A
1008-8423(2016)02-0173-06
10.13501/j.cnki.42-1569/n.2016.06.016
2016-04-07.
湖北省教育厅重点项目(D20122901).
黄伟(1990-),男(土家族),硕士生,主要从事森林生态与生物多样性保护理论的研究;*
艾训儒(1967-),男(土家族),博士,教授,主要从事森林生态与生物多样性保护理论的研究.
