太行山区物种生态位特征以及种间相关性对比分析
2016-09-26冯佳伟赵利新叶永忠
王 进, 冯佳伟, 牛 帅, 赵利新, 叶永忠
(1. 河南农业大学,河南 郑州450000; 2. 焦作市林场,河南 焦作454191)
太行山区物种生态位特征以及种间相关性对比分析
王 进1, 冯佳伟1, 牛 帅1, 赵利新2, 叶永忠1
(1. 河南农业大学,河南 郑州450000; 2. 焦作市林场,河南 焦作454191)
对河南省焦作市太行山1 hm2样地内的优势物种进行生态位宽度指数(B(sw)i)、生态位相似性比例指数(Cih)、生态位重叠指数(Lih和Lhi)等定量分析,并用点格局做种间空间格局分析。结果表明,(1)样地内优势物种生态位宽度的排序为:栓皮栎>刺槐>油松>黄栌>连翘>牡荆>扁担杆,生态位宽度值存在较大的差异,主要是植物本身生物生态学特性决定的。(2)样地内各优势物种种对的生态位相似比例值较高,生态位重叠值普遍较低。群落中生活习性相似的物种,具较高的生态位相似比例和生态位重叠。(3)样地内优势物种间存在负相关性的种对共32对,有6对物种的种间负相关尺度相加超过10 m,说明该群落优势物种在生长过程中存在某种竞争。(4)生态位重叠并不能反映种间竞争,点格局的分析方法弥补了生态位对于种间关系研究的不全面性,较全面地反映了群落中优势物种的种间关系。
优势物种;生态位;点格局;种间关系
群落内物种种间关系是物种空间分布、群落演替、物种共存机制的基础[1-4]。生态位理论是群落生态学研究的重要理论之一,生态位理论认为,不同的物种受到不同生境因子的影响,各物种通过占有不同的资源、时间和空间以实现共存,从而表现出一定的相关性[5-7]。因此,研究植物种群的种间相关性有助于认识植物的空间分布特点、种群利用环境资源的状况、植物种群的生态学过程及其与生境的相互作用等。杨宗慧等[8]对篦子三尖杉群落优势种群的生态位特征研究发现其群落中种群之间不存在较大的利用性竞争,张卓文等[9]对大别山自然保护区天然次生林的生态位特征研究表明,对于大多数演替阶段群落中的物种间的关系,以生态位重叠指数和物种生活特性作为主要的评价标准是可行的。以往的研究只是单独研究物种的生态位特征或物种的空间格局以及种间相关性,很少有研究将2种分析进行对比。本研究将生态位研究和空间格局研究相结合,综合分析样地内优势物种的生态位特征与种间相关性的关系,以期深入认识物种共存及演替的生态学过程,为该区生物多样性保护、植被恢复和次生林改造等提供理论参考。
1 材料与方法
1.1研究区概况
研究区域位于河南省焦作市太行山区,属大陆性季风气候。年均气温14.9 °C,1月份平均气温1.3 °C,7月份平均气温26.4 °C。年日照时数2 272 h,年日照率为51%,无霜期210 d。年降水量为534 mm。土壤以棕壤土、褐土为主,土层厚度为50~60 cm[10-11]。
1.2样地调查
2010—2014年,在焦作市中站区的太行山区域内选择木本植物群落保护较好的区域,参照BCI 50 hm2样地(CTFS)[12]的技术规范,建立1个100 m×100 m的样地,坐标为35°18′27′′N~35°18′31′′N、113°40′7′′E~113°40′8′′E。用全站仪将样地分成25个20 m×20 m的小样方。测量样方内所有胸径≥1 cm的木本植物的坐标、胸径、高度、冠幅等,定位并挂牌。该区域地带性植被以暖温带落叶阔叶林为主。研究样地内主要物种为栓皮栎(QuercusvariabilisBl.)、刺槐(RobiniapseudoacaciaLinn.)、油松(PinustabuliformisCarr.)、黄栌(CotinuscoggygriaScop.)等。灌木物种有连翘(Forsythiasuspensa(Thunb.) Vahl)、扁担杆(GrewiabilobaG.Don)、牡荆(VitexnegundoL.)等。样地内所有胸径大于1 cm的木本植物的平均高度为3.95 m,平均冠幅为1.54 m,平均胸径为4.83 cm。
1.3数据分析
1.3.1 重要值 重要值是表示群落中某一种群重要性的常用指标[13-14]。公式为:重要值(IV)=(相对密度+相对频度+相对显著度)/3,其中,密度指某样方内某种植物的个体数占样方面积的比例,相对密度=(某种植物的密度/全部植物的总密度)× 100%;频度指1个种在全部样方中出现的频率,重要值计算频度是以每个小样方内出现的次数进行计算,相对频度=(该种的频度/所有种的频度总和)×100%;显著度指样方内某种植物的胸高断面积除以样地面积,相对显著度=(样方中该种个体胸面积和/样方中全部个体胸面积总和)×100%。计算样地内各种群重要值,以重要值排在前7位的种群作为研究对象(表1)。
1.3.2 生态位特征 植物群落内优势物种的垂直生态位反映了不同物种在各高度资源单位内对资源的利用情况[15-16]。垂直生态位中资源位的划分是根据树木的高度划分,根据最大高度值划分合理的等级数,并确保每个资源等级为内的树木数量足够多,以保证计算准确性,一般以1 m或者2 m为1个级别。本研究中样地内胸径大于1 cm的树木数量不多,若以1 m高度为1个资源级划分,则个别资源级内出现的物种个体数量较少,会造成结果不准确,故以2 m高度为1个资源级,将高度划分为7个等级,保证每个资源位中物种个体数量较充分,以此为基础,对优势种群的垂直生态位进行计算。
(1)生态位宽度 本研究采用Shannon-Wiener多样性指标表征[17-19],公式为:
式中:B(sw)i表示物种i的生态位宽度,其域值为[0,1];S表示种群数;r表示资源位数;Pij表示物种i利用第j资源位的资源量占其利用全部资源位的比例。

表1 优势种群重要值Table 1 The importance value of dominant populations
式中:nij表示物种i在第j种资源位点的个体多度,Yi表示物种i所利用的全部资源位多度之和。
(2)生态位相似性比例 生态位相似性比例是指2个物种利用资源的相似程度,比值愈大,2个物种对资源的利用情况愈相近[20-21],公式为:
式中:Cih表示物种i和物种j的生态位相似程度,其域值为[0,1],且Cih=Chi;Pij表示物种i利用第j资源位的资源量占其利用全部资源位的比例;Phj表示物种h利用第j资源位的资源量占其利用全部资源位的比例。
(3)生态位重叠 生态位重叠是指在一定资源序列上,2个物种利用同等级资源而相互重叠的情况[22-23],公式为:

式中:Lih表示物种i重叠物种h的生态位重叠值;Lhi表示物种h重叠物种i的生态位重叠值,其域值为[0,1]。BL表示Levins生态位宽度指数。Lih、Lhi具域值[0,1],BL具域值[1/r,1]。
1.3.3 点格局分析 本研究采用双变量函数g(r)进行点格局分析。g(r) 函数是对全部定位的成对个体之间距离的格局分析,是距离依赖关联性函数。其公式为:
式中:n表示物种在样地内总的个体数,dij表示圆心点i和点j之间的距离,Wij表示以点i为圆心,dij为半径的圆的面积在样地面积A中所占的比例[24-26]。
点格局分析法要求慎重选择零假设模型。由于样方内存在空间异质性(如土壤、水流等)影响,因此本研究采用异质性泊松过程的零假设模型,分析不同物种间的空间关联性,以期得到真实准确的物种空间分布特征。异质性泊松过程模型运用圆形移动窗口在样方内进行插值计算,函数λ(x,y)依据单个物种具体位置的不同而改变,以此估计出λ(x,y)的值。其模型公式为:
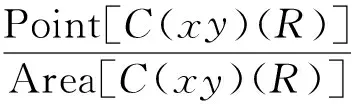
式中:C(x,y)(R)表示该圆形移动窗口以点(x,y)为圆心、R为半径,Point[X]代表区域X内的点数,Area[X]代表区域X的面积。
通过199次Monte-Carlo随机模拟计算上、下包迹线,得到置信度为99%的置信区间。若g(r)值高于上包迹线,表示2类格局为显著正相关,若g(r)值在上下包迹线之间,则符合随机分布,若g(r)值低于下包迹线,则表示2类格局为显著负相关。试验数据分析使用R语言软件完成。
2 结果与分析
2.1优势物种生态位宽度分析
生态位宽度表示种群在群落中所利用的各种资源的总和,以此来度量物种对资源多样性的利用水平。垂直生态位反映不同物种在各高度资源位内的资源占有量,同时也在一定程度上表征了森林生态系统的演替趋势。生态位宽度越大,表明物种的适应幅度越大,不同高度的个体在各个资源位内的分布也越均匀。由表2可知,样地内优势物种的生态位宽度排序为:栓皮栎>刺槐>油松>黄栌>连翘>牡荆>扁担杆。乔木物种和灌木物种受其本身的生物学特性制约,生态位宽度值有明显的区别。生态位宽度较大的是乔木、亚乔木,如栓皮栎、刺槐、油松、黄栌,反映其生态位宽度的Shannon-Wiener指数分别为0.789 3、0.703 3、0.625 9、0.564 2,这些种对垂直空间利用较充分,个体数多,树体较高且在垂直空间分布较均匀;生态位宽度较小的是灌木类,包括连翘(0.498 9)、牡荆(0.371 3)、扁担杆(0.291 6)。

表2 优势物种的生态位宽度Table 2 The horizontal niche of dominant species
2.2优势物种生态位相似性比例分析
生态位相似性比例通常与物种的数量及其自身的生物学、生态学特性相关。本研究优势物种生态位相似性比例见表3。表3中,Cih最小的是栓皮栎-扁担杆(0.091 2),种对扁担杆-刺槐(0.107 9)、牡荆-栓皮栎(0.162 4)、牡荆-刺槐(0.179 1)的生态位相似比例也较小,说明这些成对的单个种群对生境的要求差异性大。Cih值较大的多存在于同一生活型的种对中,如牡荆-扁担杆(0.924 1)、黄栌-油松(0.906 0)、栓皮栎-刺槐(0.777 2)等。优势种群间相似比大于0.6的种对占总对数的38.10%(图1);大于0.4的种对占总对数的52.38%;大于0.2的种对占总对数的80.95%,样地内优势物种生态位相似程度较大,表明在该林分中各优势物种对资源利用的相似性或物种间自身的生态学特性相似性程度较大。

表3 优势物种的生态位相似性比例值Table 3 The niche similarity of dominant species
注:1:栓皮栎;2:连翘;3:黄栌;4:牡荆;5:刺槐;6:油松;7:扁担杆。下同。
Notes: 1:QuercusvariabilisBl.; 2:Forsythiasuspensa(Thunb.) Vahl; 3:CotinuscoggygriaScop.; 4:VitexnegundoL.; 5:RobiniapseudoacaciaLinn.; 6:PinustabuliformisCarr.; 7:GrewiabilobaG.Don.The same as below.

图1 生态位相似比例分布范围Fig.1 The proportional allocation of niche similarity
2.3优势物种生态位重叠分析
群落中复杂的生态关系导致2个或更多的植物种群对某些资源产生共同的需求,使物种间出现不同程度的生态位重叠现象。种群间生态位重叠值高反映出种群对资源利用的相似程度较高,生态位重叠值低反映出种群间资源利用出现分化[27]。样地内优势物种生态位重叠值见表4。由表4可知,7个种群形成42个物种对,种树间均发生生态位重叠。优势物种间的生态位重叠值只有18对的值大于0.1,生态位重叠值较低,说明该林分中的主要物种生物学和生态学习性差异较大,能够较大程度地利用该生境中的全部资源。样地内优势物种所组成的42个种对中,连翘对扁担杆的Lih最大,为0.165 7,牡荆对刺槐的Lih最小,为0.016 0;连翘对油松的Lhi最大,为0.150 1,
栓皮栎对扁担杆的Lhi最小,为0.012 1。1个种对中,前1个种对后1个种的重叠值高,但反过来则不同,如栓皮栎对刺槐的垂直生态位重叠值为0.135 6,但刺槐对栓皮栎的垂直生态位重叠值为0.109 3。本研究中Lih大于Lhi的种对有14个,占33.33%。
所有种对的生态位均存在一定程度上的重叠。由图2可知,Lih和Lhi在0.03~0.05和0.13~0.15值域范围内比例相等。Lih小于0.09和大于0.09的种对所占比例相近,分别为52.38%和47.62%;Lhi小于0.09的种对占了较大比例,为61.90%,大于0.09的种对所占比例较小,为38.1%。
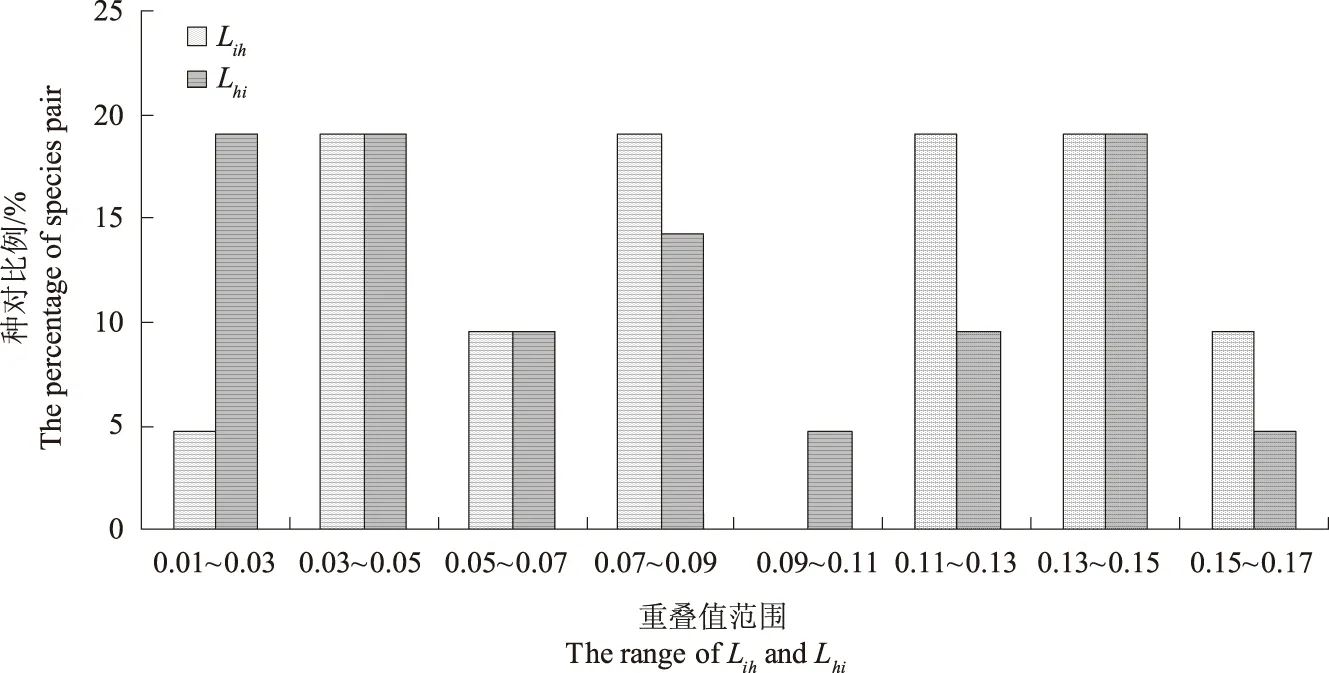
图2 生态位重叠比例分配Fig.2 The proportional allocation of niche overlaps
2.4优势物种空间关联性分析
种群的空间格局是物种与环境长期相互作用的结果,这与物种本身的生物学特性、生态环境和竞争排斥等密切相关,因此种间关联特征可能是物种间相互作用的结果,也可能是受物种生境趋同或趋异的影响[28-29]。在42个物种对中,物种间存在明显相关性的有32对,均为负相关,这表明样地内优势物种间在生长过程中存在某种竞争,另有11对物种在所有尺度上均无显著相关性,彼此之间不存在竞争和依赖。种间负相关性较显著的种对多出现在其余5个物种与刺槐、油松所组成的种对上,如扁担杆对油松、牡荆对刺槐等(表5)。
由图3知,种间负相关尺度相加超过10 m的有6对,占总对数的14.29%,具显著负相关;在5~9 m上存在空间相关性的有6对,占总对数的14.29%;在1~5 m尺度上存在空间相关性的种对有19对,占总对数的45.24%,关联性不显著;不存在空间相关性种对有11对,占总对数的26.18%。
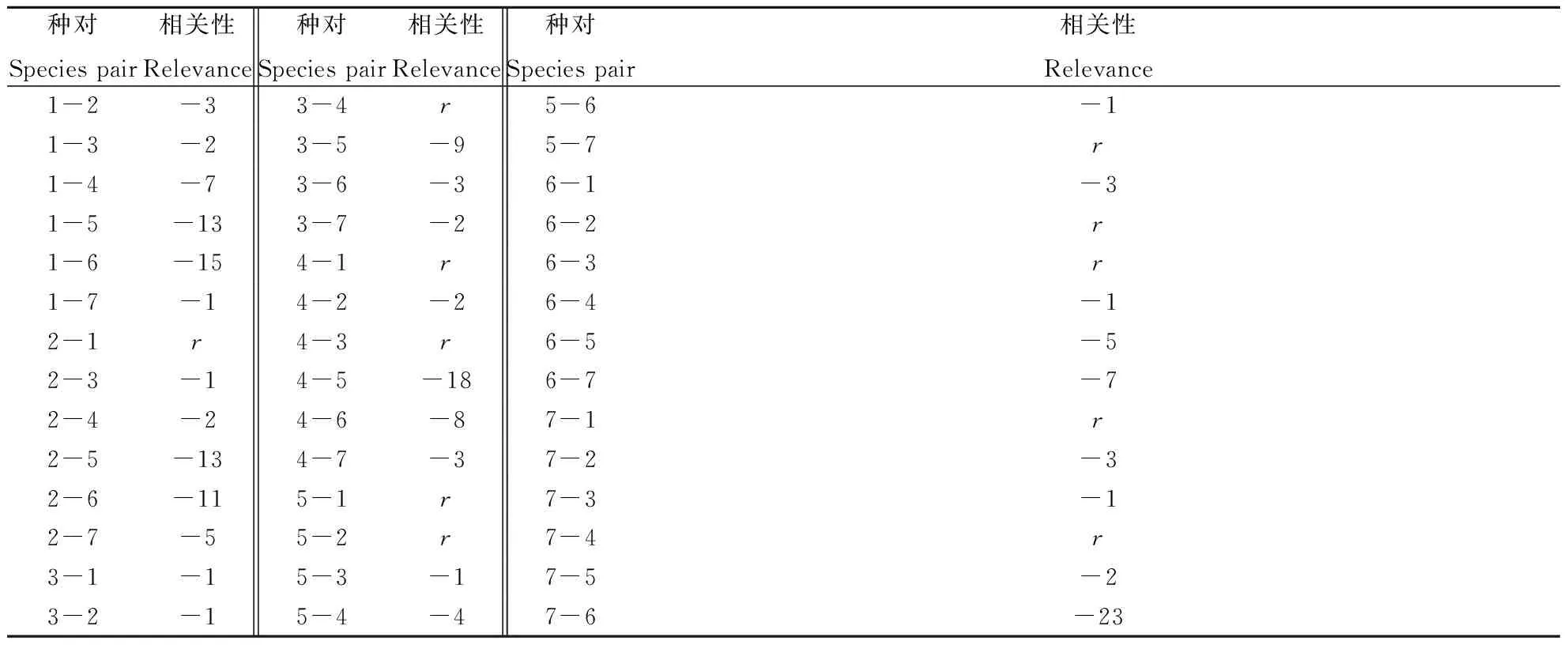
表5 优势物种间的空间格局Table 5 The spatial patterns of dominant species
注:“r”表示不存在相关性;“-”表示负相关;数字表示物种间相关性的尺度数,如-3代表物种1对物种2表现出负相关的尺度数为3 m。
Note: “r”represents no relevance; “-”represents negative corrleation; "Correlation" indicates that the number of species is significant correlation scale related to the number of scales, such as -3, which represents a negative correlation of 3 m between Species 1 and Species 2, 3 m.

图3 物种空间相关性分配比例Fig.3 The proportional allocation of spatial association
3 结论与讨论
3.1物种生物生态学特性决定其生态位宽度
在本研究的样地中,连翘、黄栌、扁担杆3个灌木类物种的生态位宽度值明显较低;栓皮栎、刺槐等乔木、亚乔木类的生态位宽度值相对较高且相差不大。种群的生态位宽度是由内外因素共同决定的,而植物的生物生态学特性起着决定性作用。栓皮栎的主要生态特征是耐寒、耐干旱瘠薄、抗风,喜生于阳光充足、通风的生境中,天然分布地段多为向阳的陡坡或山脊。故栓皮栎具有最广生态位,数量多,分布广,对资源环境的利用程度高,具最大竞争优势,也是焦作太行山南坡的主要优势物种之一,在这一时期的群落演替中起重要作用。与栓皮栎相比,生态位宽度较大的刺槐和油松也基本具备这些特征,但它们的最大高度通常小于栓皮栎的最大高度,在竞争阳光时能力弱于栓皮栎,因此是植物群落中的亚优势种。虽然从重要值来看,连翘的重要值大于除栓皮栎以外的其他乔木,但乔木物种的生态位宽度远大于灌木物种的生态位宽度,这是由于灌木种群自身的生物学特性(植物体矮小)使它们对以光因子为主导的资源利用率较低,在不同资源位分布不均匀,主要集中在较低的高度资源位中所致,因此其生态宽度低于在各资源位分布都较为均匀的乔木。
3.2生态位特征间的关系
从本研究的生态位相似值和生态位重叠值来看,样地内各优势物种种对的生态位相似比例值大于0.5的占52.38%,生态位相似比例值高;但生态位重叠值有54.14%在0.1以下,生态位重叠值较小。造成这种现象的主要原因是样地内的优势物种均为喜光物种,但因共处同一生境,物种间表现出在一定程度上对生态因子适应的互补性,部分物种增强其耐荫庇性能,同时对土壤以及地形因素的适应较为一致,因此各优势种群间的生态位相似比例较大,但由于对光需求的差异,不同物种生长在不同的林层中,这造成了生态位的分化,进而使物种之间的生态位重叠值一般都较小。此外,种群间整体上的生态位重叠值较小,与该区森林植物种类较少,群落结构简单也有关。总体上,生态位相似值与生态位重叠值保持一致,Cih大的值,其种对的Lih和Lhi均较大。
生态位宽度较大的物种,生态相似性比例值较大,与其他种群间的生态位重叠也较大,如栓皮栎在栓皮栎群落中生态位宽度最大,与其他种群的相似性和生态位重叠也较大。研究结果表明,虽然有些种群的生态位宽度值小,但种间的生态位相似比例值和生态位重叠值却较大,这是由于植物种类本身的生物学特性和生态学特性接近,故对环境资源的要求一致,这可以使环境资源得到充分利用。
3.3物种的种间相关性
通过点格局对群落内物种的种间相关性分析发现,栓皮栎林中优势物种之间的空间关系表现各异,大致分为负相关的组合和无明显空间关联性的组合2类。栓皮栎群落优势物种所组成的种对中,负相关的种对数占总种对数的76.19%,其中负相关尺度在5 m以上的有12对,占总种对数28.57%,栓皮栎、连翘、黄栌、牡荆、扁担杆这5个物种对刺槐、油松在多个尺度上都存在明显的负相关性,而这5个物种之间几乎不存在空间关联性,这说明该群落整体结构并不稳定,群落内物种还处在不断变化中。在群落的演替与发展过程中,物种出现互补的生理或生态特征,以此达到与环境和谐统一,与其他物种共存。
3.4生态位重叠与种间竞争并不保持完全一致
种群间的生态位重叠和种间负相关都是由于使用资源环境的重叠性而引起的,故生态位重叠与种间空间关联性存在一定的关系。一般来说,生态位重叠值较高的种对,对同一环境资源存在竞争,难以长期共存,其种间负相关性也较显著。如栓皮栎-刺槐、连翘-扁担杆等生态位重叠值较高且存在显著的负相关性。但生态位重叠值较高并不完全反映种间竞争程度,如刺槐-栓皮栎、扁担杆-牡荆等种对间,生态位重叠值较高,却无明显的种间竞争,这表明种对之间所需资源相对丰富,并随着群落演替与发展,相互适应并协同进化,在资源的利用上形成彼此促进的关系,从而减少种间竞争。
此外,不同生活型的物种之间如牡荆-刺槐、扁担杆-油松之间种间生态位重叠值并不高,但负相关最显著,主要原因是生态位反映物种对资源的利用情况,垂直生态位显示了物种对光照的需求。样地中刺槐、油松成年树体高大,利用光能力较强,在以光照为主的资源位划分中占据较强优势,灌木个体较小,不能与高大乔木竞争光照,但乔木幼树与灌木处于同一高度且耐荫庇能力不强,故受灌木物种的影响较大,种间负相关显著。而乔木幼树与灌木物种的竞争成功与否将决定小树能否成活,虽然灌木-乔木的生态位重叠为小,但灌木通过影响乔木物种的更新换代可以影响整个乔木物种种群。并且本研究发现,虽然相同生活型物种种对间的生态位相似比例值与生态位重叠值高,但其种间负相关性并不一定大;而不同生活型物种生态位重叠值整体偏小,但与种间相关性也无明显关系。这表明不论是相同生活型的物种还是不同生活型的物种的生态位重叠的大小与种间关系都无必然联系,在研究种间关系时不能完全依赖生态位重叠或生态位相似性比例。
综上所述,在焦作市太行山栓皮栎林养护过程中,不仅应该了解优势物种的生态位宽度、生态位相似性比例、生态位重叠等指数,也应充分考虑种群间的竞争关系,以保证存在竞争关系的种群间至少在某一维资源上不重叠,避免形成恶性竞争。同时,由于生态位重叠较大并且无显著竞争的几个物种之间在资源的利用上会形成彼此促进的关系,因此在森林养护过程中应对这几个物种同时采取保护措施,平衡种群关系,以提高森林生态系统的多样性水平。而对于生态位重叠较大并且存在竞争的几个物种,可以适当减少一部分与目标物种竞争的其他物种个体,从而保证目标物种在群落内的稳定发展。
[1] ZHU Y, GETZIN S, WIEGAND T, et al. The relative importance of Janzen-Connell effects in influencing the spatial patterns at the Gutianshan subtropical forest[J]. Plos One, 2013, 8(9): 1-8.
[2] 范娟, 赵秀海, 汪金松, 等. 江西九连山亚热带常绿阔叶林优势种空间分布格局[J]. 生态学报, 2012, 32(9): 29-39.
[3] DRUCKENBORD D L, SHUGART H H, Davies I. Spatial pattern and process in forest stands within the Virginia piedmont [J]. Journal of Vegetation Science, 2005, 16(1): 37-48.
[4] OLAGOKE A O, BOSIRE J O, BERGER U. Regeneration ofRhizophoramucronata(Lamk.) in degraded mangrove forest: Lessons from point pattern analyses of local species interactions[J]. Acta Oecologica, 2013, 50: 1-9.
[5] LEVINE J M, HILLERISLAMBERS J. The importance of niches for the maintenance of species diversity[J]. Nature, 2009, 461(7261): 254-257.
[6] WANG Z, CHEN Y, CHEN X, et al. Temporal-spatial niches of Chinese white wax scale insect (Ericerus pela) and its three dominant parasitoid wasps[J]. Acta Ecologica Sinica,2011,31(4):914-922.
[7] SUZAN-AZPIRI H, ENRIQUEZ-PENA G, MALDA-BARRERA G. Population structure of the Mexican baldcypress (TaxodiummucronatumTen.) in Queretaro, Mexico [J]. Forest Ecology and Management, 2007, 242(2): 243-249.
[8] 杨宗慧, 郎学东, 李帅锋, 等. 篦子三尖杉群落优势种群生态位和种间关系[J]. 林业科学研究, 2015, 28(4): 473-478.
[9] 张卓文, 王永健, 唐亚坤, 等. 大别山自然保护区天然次生林分类及其生态位特征[J]. 华中农业大学学报(自然科学版), 2013, 31(6): 686-692.
[10] 宋朝枢, 瞿文元. 太行山猕猴自然保护区科学考察集[M]. 北京: 中国林业出版社, 1996.
[11] 王育水, 权玉萍, 辛泽华, 等. 南太行山区黄刺玫资源调查及开发利用[J]. 焦作师范高等专科学校学报, 2014 (2): 77-79.
[12] CONDIT R. Tropical forest census plots: methods and results from Barro Colorado Island, Panama and a comparison with other plots[J]. Springer,1998: 1-224.
[13] 范娟, 赵秀海, 汪金松, 等. 江西九连山亚热带常绿阔叶林优势种空间分布格局 [J]. 生态学报, 2012, 32(9): 2729-2737.
[14] ZHANG J, HAO Z, SONG B, et al. Spatial distribution patterns and associations ofPinuskoraiensisandTiliaamurensisin broad-leaved Korean pine mixed forest in Changbai Mountains[J]. Chinese Journal of Applied Ecology, 2007, 8: 002.
[15] 陈璟. 衡阳盆地紫色土丘陵坡地自然恢复灌丛阶段植物群落主要种群生态位特征[J].中国农学通报2009,25(23):147-154.
[16] 王祥福, 郭泉水, 巴哈尔, 等. 崖柏群落优势乔木种群生态位[J]. 林业科学, 2008, 44(4): 6-13.
[17] 张金屯. 数量生态学[M]. 北京: 科学出版社, 2004: 78-177.
[18] 陈波, 周兴民. 三种嵩草群落中若干植物种的生态位宽度与重叠分析[J]. 植物生态学报, 1995, 19(2): 158-169.
[19] ROZAS V. Structural heterogeneity and tree spatial patterns in an old-growth deciduous lowland forest in Cantabria, northern Spain [J]. Plant Ecology, 2006, 185(1): 57-72.
[20] ZHIYAO S U. Niche characteristics of dominant populations in natural forest in north Guangdong[J]. Chinese Journal of Applied Ecology, 2003, 14(1): 25-29.
[21] 李菁,骆有庆,石娟,等. 阿尔山地区兴安落叶松林下植物种群生态位[J]. 林业科学研究, 2011, 24(5): 651-658.
[22] 曾岳明, 张超,周文春. 浙江莲都区公益林主要植物种生态位研究[J]. 山东林业科技, 2015, 45(3): 10-16.
[23] 谢琼中. 南方铁杉群落物种多样性及乔木优势种生态位初步研究[J]. 天津农业科学, 2011, 17(2): 133-136.
[24] 缪宁, 刘世荣, 史作民, 等. 青藏高原东缘林线杜鹃-岷江冷杉原始林的空间格局[J]. 生态学报, 2011, 31(1): 1-9.
[25] 范娟, 赵秀海, 汪金松, 等. 江西九连山亚热带常绿阔叶林优势种空间分布格局 [J]. 生态学报, 2012, 32(9): 2729-2737.
[26] WIEGAND T, A MOLONEY K. Rings, circles, and null-models for point pattern analysis in ecology[J]. Oikos, 2004, 104(2): 209-229.
[27] 刘巍, 曹伟. 长白山云冷杉群落主要种群生态位特征[J]. 生态学杂志, 2011, 30(8): 1766-1774.
[28] FEELEY K J, SILMAN M R. Biotic attrition from tropical forests correcting for truncated temperature niches[J]. Global Change Biology, 2010, 16(6): 1830-1836.
[29] 刘彤, 赵新俊, 崔运河, 等. 天山北麓中段拟南芥(Arabidopsisthaliana)与相邻物种的分布格局及相互关系[J]. 生态学报, 2008, 28(4): 1842-1849.
(责任编辑:朱秀英)
ComparativeanalysisofnichecharacteristicsofthespeciesandinterspecificcorrelationinTaihangmountains
WANG Jin1, FENG Jiawei1, NIU Shuai1, ZHAO Lixin2, YE Yongzhong1
(Henan Agricultural University, Zhengzhou 450000, China; 2. Forest Farm of Jiaozuo, Jiaozuo 454191, China)
Quantitative analysis was carried out on the niche breadth (B(sw)i) , niche similarity (LihandLhi) , and niche overlap (Cih) of the dominant species of 1 hm2plot in Taihang Mountains, were used by quantitative analysis with dominant species. In the mean time, point pattern analysis was carried out to study of the spatial distribution pattern. The results show that: (1) in the plot, the order of niche breadth values is:QuercusvariabilisBl.>RobiniapseudoacaciaLinn.>PinustabuliformisCarr.>CotinuscoggygriaScop.>Forsythiasuspensa(Thunb.)>VitexnegundoL.>VitexnegundoL. Because of the biological and ecological characteristic of plants, there exists bigger difference in the niche breath. (2) The niche similarity value of dominant species is high, while the niche overlap value is generally low. In the Quercus variabilis Bl. forest, the species with similar ecology characters have higher niche similarity and niche overlap values. (3) A total of 32 dominant species pairs have a negative correlation, and there are 6 species pairs negative scale added over 10m, which shows that the dominant species in the community have some competition to each other.(4) The niches overlap is not a reflection of interspecific competition. Point pattern analysis method can make up for the niche for comprehensive research on the relationship between species, and it accurately reflects the interspecific relationship of main species of Quercus variabilis Bl. community.
dominant species; niche ; point pattern; interspecific relationship
S 718.5
:A
2015-10-22
国家科技支撑计划项目(2008BAC39802)
王 进(1990-),女,河南方城人,硕士研究生,研究方向为植物生态学。
叶永忠(1957-),男,湖北黄冈人,教授,博士生导师。
1000-2340(2016)02-00181-08
