不同泡桐无性系光合特征参数差异分析
2016-09-26贾惠文吴雅文王艳梅
贾惠文, 郭 芳, 吴雅文, 姚 兵, 王艳梅, 刘 震
(河南农业大学林学院,河南 郑州 450002)
不同泡桐无性系光合特征参数差异分析
贾惠文, 郭 芳, 吴雅文, 姚 兵, 王艳梅, 刘 震
(河南农业大学林学院,河南 郑州 450002)
以1 a生1B、2B、3B、9501共4种泡桐优良无性系为材料,采用LCPRO+便携式光合测定系统,对其光合日均值和光合-光响应曲线进行测定研究。结果表明,1)4种泡桐无性系光合特征日均值差异显著,净光合速率日均值大小依次为9501>1B>3B>2B;2)4种泡桐无性系随光强变化的特性参数各不相同,综合各光合特征参数分析4种泡桐无性系光合速率随光强变化的大小依次为1B>9501>3B>2B;3)泡桐无性系1B不仅在弱光下光合能力强,在适中光强和高光强下,同样可以进行良好的光合作用;4)表观量子效率最大的泡桐无性系1B能高效的将光能转换为所需的化学能。
泡桐无性系;净光合速率;光合-光响应曲线;光合特征参数;表观量子效率
光合作用是地球上规模最大的把太阳能转变为可储存的化学能的过程,也是规模最大的将无机物合成有机物和释放氧气的过程[1]。它是蓄积能量和形成有机物的过程,植物生长所需的大部分营养和干物质,是通过光合作用直接或间接得来的[2]。泡桐是中国平原农区与农作物间作的重要速生用材树种,也是出口创汇的主要树种[3]。前人对毛白33、不同品种泡桐四倍体等的不同生长期光合日变化等进行了一定的研究[4-5],但对泡桐在不同光强下的光响应特点还未见报道。本研究利用美国ADC BioScientific生产的LCPRO+便携式光合测定系统对4种泡桐优良无性系的光合日均值和光合-光响应曲线进行了测定,探讨不同泡桐无性系的光合特性和差异,以及光合有效辐射与净光合速率、光补偿点和光饱和点的关系,试图为泡桐的良种选育奠定理论基础。
1 材料与方法
1.1试验地概况
试验地位于郑州市河南农业大学林业试验站,暖温带大陆性气候,四季分明,年平均气温为14.2℃,极端最高温度为43℃,极端最低温度为-17.9℃,≥10℃积温为4 717℃,无霜期为215 d,年平均降水量为650.1 mm,全年日照时数大约为2 400 h。土壤为潮土类的砂土,微碱性,pH值为8.0,有机质含量为7.6 g·kg-1,全氮含量为0.58 g·kg-1,全磷含量为1.34 g·kg-1,全钾含量为22.8 g·kg-1。
1.2试验材料
供试材料为1 a生4种嫁接泡桐无性系,分别为1B、2B、3B和对照无性系9501。其中,杂交泡桐“9501”系国家林业局泡桐研究开发中心“九五”科技攻关期间选育出的泡桐优良品种[6]。
1.3试验方法
1.3.1 试验材料 每种泡桐无性系随机选择3株生长状况良好,且高度一致的苗木作为测定对象,确定每株苗木高度2/3处东南方向的3个叶片为测定样本。测定时间为嫁接当年,即2015-08-01—2015-08-05的9:00 至 11:00,天气晴朗,光照充足,平均气温为(35.25±2.25)℃。
1.3.2 光合日均值的测定 选叶龄、叶位一致的成熟叶片,测其净光合速率(Pn),胞间CO2浓度(Ci),蒸腾速率(Tr),气孔导度(Gs)等光合指标。
1.3.3 光合-光响应曲线的测定 采用LCPRO+便携式光合测定系统,在0 ~ 2 000 μmol·m-2·s-1光合有效辐射(PPFD)范围内,利用红蓝光源(LED)设定2 000、1 800、1 500、1 200、1 000、500、200、100、50、20和0 μmol·m-2·s-1等11个梯度,并统一设定叶室温度为(25±0.5)℃、CO2浓度为(400±1) μmol·mol-1、叶面水分饱和亏缺为(1.0±0.1)kPa 和参考室空气湿度为(50±6.52)%。从强光到弱光测定各叶片的净光合速率和相关参数,并绘制光响应曲线。
不同无性系泡桐叶片的净光合速率Pn与PPFD之间的关系(光合-光响应曲线)采用经验方程[7-8]:
Pn=Pmax(1-C0e-aPAR/Pmax)
(1)
式中:Pmax为最大净光合速率(μmol·m-2·s-1);a和C0为度量弱光下净光合速率趋近于零的常数;Pn为特定光强下的净光合速率。
通过适合性检验,若方程良好(达到95%显著水平),则可利用(1)式分别计算光补偿点(LCP,μmol·m-2·s-1)、光饱和点(LSP,μmol·m-2·s-1)、表观量子效率(Φ,mol·mol-1)和光合幅度(PR,μmol·m-2·s-1)等[8]。
2 结果与分析
2.1不同泡桐无性系光合特性的差异
净光合速率是植物组织在有光的条件下,光合作用和细胞呼吸同时进行测得的数值,且Ci、Tr、Gs等光合特征值的高低都对植物的光合作用有很大的影响。植物的光合作用在一天内会随着时间、光照强度、空气温度等因素的影响而变化[9],不同泡桐无性系光合特性日均值同样存在差异。用SPSS软件Post Hoc多重比较LSD方差检验法分析泡桐无性系光合特性的差异,由图1可知,无性系1B和9501的Pn值无显著差异,4种泡桐无性系Pn日均值的大小依次为9501>1B>3B>2B,图2,图3中,Ci和Tr的大小依次为2B>3B>1B>9501,图4中,Gs的大小依次为2B>3B>9501>1B,表现出4种泡桐无性系Pn与Ci和Tr呈负相关Pn与Gs之间存在一定的负相关关系。

图1 净光合速率(Pn)日均值的比较Fig.1 Comparison of daily mean value of Pn

图2 胞间CO2浓度(Ci)日均值的比较Fig.2 Comparison of daily mean value of Ci
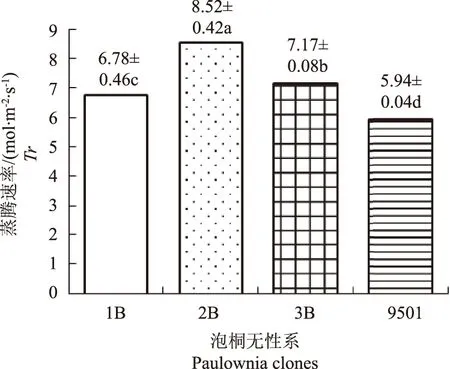
图3 蒸腾速率(Tr)日均值的比较Fig.3 Comparison of daily mean value of Tr

图4 气孔导度(Gs)日均值的比较Fig.4 Comparison of daily mean value of Gs
2.2不同泡桐无性系在不同光强下的光合速率差异
由图5可知,不同泡桐无性系的光合-光响应曲线的整体趋势大致相同,但不同无性系的光合速率在不同光强下存在差异。在光强500 μmol·m-2·s-1以下时,2B的光合速率较大,其他3种泡桐无性系的光合速率随光强变化的趋势基本一致,直到光强达500 μmol·m-2·s-1时,4种泡桐无性系的光合速率相差无几。光强大于500 μmol·m-2·s-1时,1B和9501的光合速率明显提高,3B的光合速率呈缓慢增长,2B的光合速率已基本稳定,但仍在缓慢增长。当光强大于1 500 μmol·m-2·s-1时,所有无性系的光合速率基本稳定在一定水平,表明这4种泡桐无性系对强光的适应能力较强,其净光合速率不受强光的抑制。
叶片的最大光合速率(Pn)表示了叶片最大光合作用的能力[10]。不同泡桐无性系对强光的适应能力不同,光强达2 000 μmol·m-2·s-1时, 其净光合速率较高的是1B和9501,说明无性系1B在强光下光合生产能力优于9501、3B和2B相对较弱。

图5 泡桐无性系光合-光响应曲线Fig.5 The photosynthetic-light response curve of Paulownia clones
2.3不同泡桐无性系光合-光响应曲线特征参数的差异
光补偿点与光饱和点是植物光合-光响应曲线过程中的重要特征指标。通常在光强很弱时,叶片的光合速率低于呼吸速率,随着光强的增大,光合速率等于呼吸速率,这时的光照强度称为光补偿点(LCP),当光强达到一定强度,光合速率基本稳定在一定水平,这时的光照强度称为光饱和点(LSP)[11]。植物光补偿点的高低反应了植物在弱光下的光合作用水平,补偿点越低,说明植物对弱光有较强的利用能力[12]。对按控制条件测得的4种泡桐无性系光合-光响应曲线采用公式(1)进行拟合,均能得到很好的结果(表1)。依此所计算的各无性系光合特征值见表2。
由表2可知,4种泡桐无性系拟合的最大净光合速率分别为24.60、19.42、21.20和23.30 μmol·m-2·s-1,与图5中光合-光响应曲线的表现一致。它们的光补偿点分别范围为18.37 ~ 44.89 μmol·m-2·s-1,其中3B的光补偿点最低,说明3B泡桐无性系对弱光有较强的光合作用能力,9501的光补偿点最高,说明9501对弱光的利用能力较弱,而1B和2B的弱光利用能力适中。

表1 光合-光响应曲线模型拟合参数Table 1 Model fitting parameters of the photosynthetic-light response curve

表2 光合-光响应曲线的特征参数Table 2 Characteristic parameters of the photosynthetic-light response curve
4种泡桐无性系光饱和点的范围为1 860.44 ~ 2 234.69 μmol·m-2·s-1,9501的光饱和点最高,为2 234.69 μmol·m-2·s-1,1B的光饱和点次之,表明9501和1B这2种无性系喜阳性更好,阳光充足时能很好的将光能转化为所需物质。光饱和点较弱的是2B和3B,反映其在强光下的光合作用相对较弱。
光饱和点和光补偿点之间的差值为光和幅度(PR),其差值越大,表明植物的光合作用范围越广,对自身的生长越有优势。从表2可看出,9501的光合幅度最广,表明其可利用的光强区域最宽,1B的光合幅度相对9501略小,但也能有足够的光照区域来进行光合作用。
2.4不同泡桐无性系半饱和点、近饱和点的差异
半饱和点是指光合速率为饱和光合速率50%时的光照强度,近饱和点是指光合速率为饱和光合速率95%时的光照强度[13]。从图5和表2的对比可以看出,在补偿点和半饱和点之间,光合速率整体呈加速增长趋势,在半饱和点和近饱和点之间,光合速率整体表现为缓慢增长,当光强大于近饱和点时,所有泡桐无性系光合速率的增加则更加微小。4种泡桐无性系中,1B的半饱和点最高,3B的半饱和点最低,表明3B的光合速率能在光强较弱的情况下加速增长,1B则需较高的光强加速其光合速率。近饱和点的比较中,1B、2B和9501较高,表明在高光强下,仍能高效利用光能进行光合速率,对光能的利用率较高。
2.5不同泡桐无性系表观量子效率的差异
表观量子效率(Φ)是植物对CO2同化的表观量子效率,其大小是植物在光合作用过程中对光的利用率的一种表现,特别是对弱光的利用效率[14]。由表2可知,1B的表观量子效率最高(0.057),表明1B对光的利用率很高,能较高的吸收光量子进行光合作用,3B的表观量子效率次之。这与1B和3B具有较低的光补偿点是一致的。
3 结论与讨论
3.1不同泡桐无性系光合特性的关系
4种泡桐无性系光合特征日均值差异明显,净光合速率日均值依次为9501>1B>3B>2B。而关于Pn与Ci,Tr,Gs之间的关系有很多不同的观点。本研究表明4种泡桐无性系Pn与Ci和Tr呈负相关。这与前人对一些植物叶片Pn与Ci呈负相关的研究结果一致[15-16],这可能是由于Pn随着光强增大的过程中,叶肉细胞光合活性增大的结果;Gs的减小也会导致Ci的降低[17],而本试验中,4种泡桐无性系Ci的大小顺序为2B>3B>1B>9501,Gs的大小顺序为2B>3B>9501>1B,且1B与9501的Gs无显著差异,表明4种泡桐无性系的Ci与Gs之间存在一定相关性,即Ci的降低可能起因于Gs的减小,越小的泡桐无性系,其Pn越大。4种泡桐无性系Pn与Tr的负相关关系,可能是由于蒸腾速率越大,叶片水分丧失越严重,导致叶片内与光合相关的酶活性下降,这可能是Pn与Tr呈负相关的原因[18]。
3.2不同泡桐无性系光合-光响应曲线特征参数的关系
不同泡桐无性系的光合作用能力不同,4种泡桐无性系Pmax的大小表现为1B>9501>3B>2B,其范围为19.42 ~ 24.60 μmol·m-2·s-1,表现出它们的光合能力较高。
通过Pmax与LCP、LSP和PR等特征参数的比较,可以看出,泡桐无性系1B有较大的Pmax、LSP和PR,与较小的LCP。一般情况下,LCP较低,LSP较高的植物对光环境的适应性较强[16],说明1B有较强的光适应性,不仅在高光强下能有效的进行光合作用,在弱光下也能发挥其光合作用的优势。3B也有较小的光补偿点,但其Pmax、LSP和PR也小,说明其虽然利用弱光进行光合作用的能力很强,但较小的光饱和点制约了其光照幅度的扩展,进而影响其最大净光合速率。2B的Pmax在4种泡桐无性系中相对最弱,与之对应的是较大的LCP与较小的LSP、PR,较大的LCP与较小的LSP共同缩短了2B的PR,不仅减少了2B可利用的光照范围,也降低了其光合速率。9501与1B有着幅度相近的PR,但1B的LCP低于9501,表明1B相对于9501在弱光下的光合作用效率更高。4种泡桐无性系从光补偿点到半饱和点直至近饱和点的变化可以看出,1B的半饱和点最高,说明其不仅在弱光下光合能力强,特别在高光强下能够加速光合作用的效率[13]。
3.3不同泡桐无性系表观量子效率的关系
一般情况下,植物的表观量子效率为0.03 ~ 0.07[19],本研究对按控制条件测得的4种泡桐无性系光合-光响应曲线进行拟合得到的表观量子效率范围为0.046 ~ 0.057 mol·mol-1,其大小表现为1B>3B>9501>2B。Φ值最大的1B能够说明1B有着较高的光合效率,这可能是由于无性系1B的叶片在PSII反应中心有较高的能量捕获效率[20-21],同时也有高效的光子吸收,高效的NADP+和ADP再生,以及高效的电子传递[22]。
综合4种光合特征参数的分析能很好地说明:泡桐无性系1B能最大范围地吸收光能并高效地将光能转换为所需的化学能,为泡桐的生长所需要。
[1] 李合生.现代植物生理学[M].2版.北京:高等教育出版社,2012:144.
[2] 李永华,武荣花,杨秋生,等.短期CO2加富对苗期红掌生长及光合作用的影响[J]河南农业大学学报, 2006, 40(6):607-610.
[3] 蒋建平.泡桐栽培学[M].北京:中国林业出版社,1990.
[4] 王艳梅,牛晓峰,刘震,等.泡桐生长停止和顶芽死亡过程中的光合特性研究[J].北京林业大学学报,2009,31(6):121-127.
[5] 张晓申,翟晓巧,赵振利,等.不同种四倍体泡桐光合特性的研究[J].河南农业大学学报, 2013, 47(4):400-404.
[6] 彭仁奎.优良杂交泡桐品种“9501”营造技术探讨[J].安徽农学通报,2010,16(7):163-165.
[7] BASSMAN J H,ZWIER J C.Gas exchange characteristics of Populus trichocarpa, Populus deltoides and Populus trichocarpa P.deltoides clones.[J].Tree Physiol,1991,8(2):145-59.
[8] 胡新生,刘建伟.四个杨树无性系在不同温度和相对湿度条件下净光合速率的比较研究[J].林业科学,1997,33(2):107-116.
[9] 张治安,杨福,陈展宇,等.菰叶片净光合速率日变化及其与环境因子的相互关系[J].中国农业科学, 2006, 39(3):502-509.
[10] 王广军,张彦妮,何叶.不同情况复叶械(Acer negundo L)叶片净光合速率的日变化[J].东北林业大学学报,2008,36(8):14-15.
[11] 叶子飘,于强.光合作用光响应模型的比较[J].植物生态学报,2008,32(6):1356-1361.
[12] 王雁,马武昌.扶芳藤、紫藤等7种藤本植物光能利用特性及耐荫性比较研究[J].林业科学研究, 2004, 17(3):305-309.
[13] 崔令军,王保平,孙向阳,等.不同泡桐无性系苗期光合特性的研究[J].安徽农业科学,2007,35(28):8874-8875.
[14] 高泽梅,权秋梅,赵刚,等.天全岩白菜光合特性初探[J].西华师范大学学报(自然科学版),2013,34(1):72-77.
[15] 翁晓燕,蒋德安,陆庆,等.影响水稻叶片光合日变化因素的分析[J].中国水稻科学,1998,12(2):105-108.
[16] 张治安, 杨福, 陈展宇,等.菰叶片净光合速率日变化及其与环境因子的相互关系[J].中国农业科学, 2006, 3(3):502-509.
[17] 陈根云,陈娟,许大全.关于净光合速率和胞间CO2浓度关系的思考[C].纪念殷宏章先生百年诞辰暨全国光合作用学术研讨会论文摘要汇编. 2008:64-66.
[18] 高景慧,张颖, 郭维,等.分枝期3个紫花苜蓿品种光合蒸腾日变化与相关因子的关系分析[J]. 西北农林科技大学学报(自然科学版), 2007, 10(10):29-34.
[19] 刘宇锋,萧浪涛,童建华,等.非直线双曲线模型在光合光响应曲线数据分析中的应用[J].中国农学通报,2005,21(8):76.
[20] RODRIGUEZ R E, ANABELLA L, POLI H O, et al. Transgenic Tobacco Plants Overexpressing Chloroplastic Ferredoxin-NADP(H) Reductase Display Normal Rates of Photosynthesis and Increased Tolerance to Oxidative Stress[J]. Plant Physiology, 2007, 143(2):639-649.
[21] 张变莉,王杨,刘荣宁,等.同源四倍体台湾泡桐体外植株再生系统的建立[J].河南农业科学,2014,44(12):119-123.
[22] CHIKAHIRO M, KATSUMI A, NAOMASA S, et al. Acclimation of Tobacco Leaves to High Light Intensity Drives the Plastoquinone Oxidation System--Relationship Among the Fraction of Open PSII Centers, Non-Photochemical Quenching of Chl Fluorescence and the Maximum Quantum Yield of PSII in the Dark[J]. Plant and Cell Physiology, 2009, 50(4):730-743.
(责任编辑:李莹)
ThevarianceanalysisofphotosyntheticcharacteristicparametersofdifferentPaulowniaClones
JIA Huiwen, GUO Fang, WU Yawen, YAO Bing, WANG Yanmei, LIU Zhen
(College of Forestry, Henan Agricultural University, Zhengzhou 450002, China)
The photosynthetic characteristic parameters and daily average photosynthetic and photosynthetic-light response curve of 4 kinds of 1-year-old excellent Paulownia clones, 1B,2B,3B, and 9501, were determined by using LCPRO+ portable photosynthesis determination system. The results showed that: 1) The 4 kinds of Paulownia clones had obvious differences in photosynthesis characteristic parameters, the daily average of the net photosynthetic rate in order is: 9501>1B>3B>2B; 2) The characteristic parameters of 4 kinds of Paulownia clones all come differently under different light intensities, taking all photosynthetic parameters to analyze, the photosynthetic rate order of 4 kinds of Paulownia clones with the light intensity changes was 1B>9501>3B>2B; 3) The Paulownia clone 1B had strong photosynthetic ability not only under low light, but also under moderate intensity and high intensity; 4) The Paulownia clone 1B with the maximum apparent quantum yield could efficiently convert the light energy to the necessary chemical energy.
Paulownia clones; net photosynthetic rate; photosynthetic-light response curve; photosynthetic characteristic parameters; apparent quantum yield
Q945.11;S 722.5
:A
2015-09-18
河南省平原地区森林抚育技术研究项目(201401);教育部高等学校博士学科点专项科研基金项目(2008046602)
贾惠文(1989-),女,河南郑州人,硕士研究生,主要从事森林培育与树木生理生态研究。
刘 震(1964-),男,河南沈丘人,教授,博士。
1000-2340(2016)02-0176-05
