遮荫对斑叶堇菜和大叶铁线莲光合特性的影响
2016-09-25刘凯月杜绍华郭思佳
刘凯月, 杜绍华, 郭思佳, 赵 硕
(河北农业大学园林与旅游学院,河北 保定 071000)
遮荫对斑叶堇菜和大叶铁线莲光合特性的影响
刘凯月, 杜绍华, 郭思佳, 赵 硕
(河北农业大学园林与旅游学院,河北 保定 071000)
为探究遮荫对斑叶堇菜(Violavariegata)和大叶铁线莲(Clematisheracleifolia)光合特性的影响,分别对它们进行了30%、60%、90%的3个梯度的遮荫处理,以全光照作对照,研究了不同遮荫条件下2种植物的光合特性。结果表明,随着遮荫度的增加,斑叶堇菜和大叶铁线莲的净光合速率、气孔导度、蒸腾速率、胞间CO2摩尔分数、光饱和点、光补偿点、CO2饱和点、CO2补偿点降低,表现量子效率和羧化效率随遮荫度的增加而提高;不同遮荫条件下两种植物的净光合速率日变化、气孔导度日变化、蒸腾速率日变化均为双峰曲线,表现出明显的“午休”现象。表明斑叶堇菜和大叶铁线莲均具有较强的耐荫性。
斑叶堇菜;大叶铁线莲;遮荫;光合特性
乔灌草复层生态结构下层空间需要喜荫地被植物,但北方喜荫地被种类较少,常见的仅有玉簪等。因此,耐荫植物的筛选和应用在城市绿化中的地位日渐突出[1]。斑叶堇菜(Violavariegata)是堇菜科一种叶形和叶色都较别致的多年生矮小野生植物[2],是非常优秀的观叶植物,有耐寒、喜荫等优点。大叶铁线莲(Clematisheracleifolia)为毛茛科多年生灌木,蓝紫色聚伞花序观赏性很高,植株耐荫。野生状态下,2种植物均生长在庇荫的环境下,但关于不同遮荫条件对斑叶堇菜和大叶铁线莲光合特性的影响未见有报道。本试验采用人工遮光法,研究2种地被植物在夏季连续遮荫条件下的光合特性,以期为两种地被植物的园林应用提供依据。
1 材料与方法
1.1材料
试验材料为斑叶堇菜(Violavariegata)、大叶铁线莲(Clematisheracleifolia)。斑叶堇菜、大叶铁线莲均采于河北保定市易县蚕姑坨,为繁殖后的2 a生苗。
1.2方法
1.2.1 遮荫处理 采用不同透光率的黑色遮荫网进行遮荫,高度控制在1 m左右,设3个遮荫处理,遮光率分别为30%、60%、90%,以全光照(CK)为对照,遮荫时间自2015年4月初至2015年9月底。每处理30棵苗,试验重复3次。遮荫期间正常进行养护管理。
1.2.2 光合作用日变化测定 试验在2015-08-07进行,选择生长势旺盛且一致的植株作为测试样本,利用美国LI-COR公司生产的Li-6400便携式光合仪从8:00—18:00每隔2 h对2种地被植物各处理的光合作用日变化进行测定,每次每个处理5次重复,取其平均值作为该时刻的测定值。Li-6400 便携式光合作用系统记录叶片瞬时净光合速率值(Pn)、蒸腾速率(Tr)、气孔导度(Gs)、胞间 CO2摩尔分数(Ci)、叶面水汽压差(Vpdl)、大气 CO2浓度(Ca)、光合有效辐射(PAR)、空气气温(Ta)、叶片温度(Tl)、相对湿度(RH)等参数。水分利用效率WUE=Pn/Tr。光能利用效率QUE=Pn/PAR。
1.2.3 光响应曲线和CO2响应曲线测定 试验在2015年8月晴朗无风的天气进行,选择生长势旺盛且一致的植株作为测试样本,于上午9:00—11:00进行,重复3次。光响应曲线在2015-08-08测量,利用Li-6400光合仪将参比室的CO2摩尔分数稳定在380 μmol·mol-1,手动设置人工光源光照度(PAR)从高到低为1 500、1 200、1 000、800、600、400、200、150、100、70、50、30、20、0 μmol·m-2·s-1共14个梯度。利用叶子飘等[3]的光合计算软件对光响应曲线进行拟合,并根据曲线求得光饱和点(LSP)、光补偿点(LCP)、最大净光合速率(Pn)、暗呼吸速率(Rd)、最大净光合速率(Amax)以及表现量子效率(AQY)。 CO2响应曲线在2015-08-09测量,利用人工光源设置饱和光强(根据光响应曲线确定),CO2的摩尔分数从高到低为1 600、1 400、1 200、1 000、800、600、400、200、150、100、50 μmol·mol-1共11个梯度。利用叶子飘等[3]的光合计算软件对CO2响应曲线进行拟合,并根据曲线求得CO2饱和点(CSP)和CO2补偿点(CCP)以及羧化效率(CE)。
1.3数据处理
试验数据用EXCEL处理,方差分析用DPS7.05处理。
2 结果与分析
2.1遮荫对净光合速率日变化的影响
净光合速率是指植物的总光合量减去呼吸作用所消耗的量[4]。净光合速率日变化可以反映植物在不同的遮荫环境下的生长状况,净光合速率高,植物生长好,观赏价值高;净光合速率低,说明植物受到光胁迫,观赏价值降低[5]。图1表明,不同遮荫条件下斑叶堇菜和大叶铁线莲的叶片的净光合速率均表现出先上升后下降之后再上升,最后又下降的“双峰”曲线的变化规律,在12:00时2种植物各处理都表现出明显的光合“午休”现象。2种植物各处理的第一个最大值均出现在10:00;但是第二个最大值出现时间略为不同,斑叶堇菜各处理和大叶铁线莲60%遮荫和90%遮荫的第二个峰值均出现在14:00;大叶铁线莲全光照条件和30%遮荫处理的第二个峰值则出现在16:00。且各处理的第二个峰值均比第一个峰值低。随着遮荫度的增加,斑叶堇菜和大叶铁线莲各处理的峰值和日均光合速率也都呈现出了下降的趋势。斑叶堇菜在全光照下的两个峰值和日均光合速率分别比90%遮荫处理的高出了44.75%、53.94%、50.8%;大叶铁线莲则分别高出了35.82%、52.8%、46.01%。说明2种植物适应并且利用弱光的能力都很强。
2.2遮荫对气孔导度日变化的影响
气孔导度是植物气孔传导CO2和水汽的能力,气孔导度越大,代表气孔开闭程度越大;反之,气孔阻力越大,气孔开闭程度越小[6]。气孔导度对环境因子的变化十分敏感,凡是影响植物光合作用和叶片水分状况的各种因素都有可能对气孔导度造成影响[7]。由图2可以看出,不同遮荫条件下2种植物的气孔导度日变化与净光合日变化基本一致,也均表现出先上升后下降之后再上升,最后又下降的“双峰”曲线的变化规律。且各处理2个峰值出现的时间也与净光合速率的一致。在8:00和18:00时,因为光线较弱,气孔张开较小,气孔导度较低,而在12:00时由于高温强光使得气孔导度降低。斑叶堇菜和大叶铁线莲各处理的峰值和日均气孔导度随着遮荫度的增加也都呈现出了降低的趋势。斑叶堇菜在90%遮荫条件下的2个峰值和日均气孔导度分别比全光照的低了46.72%、47.85%、48.5%;大叶铁线莲则分别低了30.76%、42.13%、34.35%。
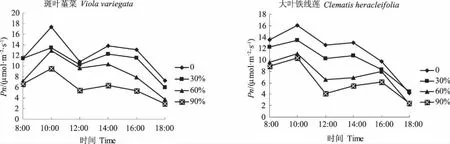
图1 遮荫对净光合速率日变化的影响Fig.1 Effects of shading on the diurnal variations of net photosynthetic rate

图2 遮荫对气孔导度日变化的影响Fig.2 Effects of shading on the diurnal variations of stomatal conductance
2.3遮荫对蒸腾速率日变化的影响
从图3可知,斑叶堇菜和大叶铁线莲在不同遮荫条件下的蒸腾速率日变化与净光合速率、气孔导度日变化趋势相似。且遮荫后蒸腾速率受到了显著影响。斑叶堇菜和大叶铁线莲的各处理在10:00时的蒸腾速率都达到了1 d中的最大值,而在12:00时蒸腾速率都最低,这可能是因为气孔导度对于蒸腾速率有较大影响。2种植物各处理的蒸腾速率都随着遮荫度的增加而呈现出了减小的趋势。这说明斑叶堇菜和大叶铁线莲通过降低蒸腾速率来适应弱光环境。
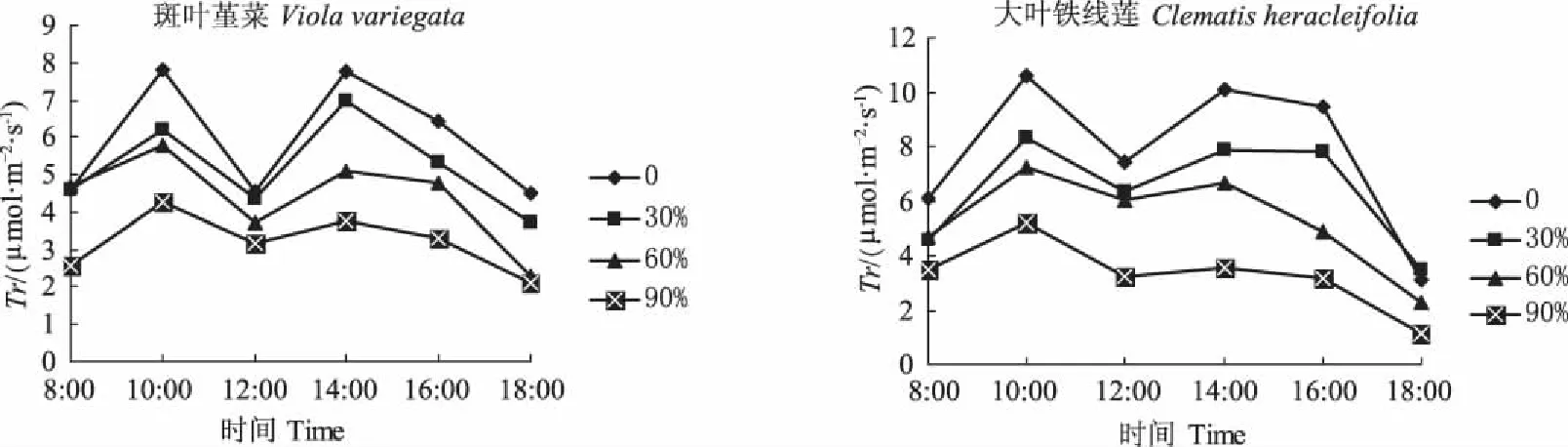
图3 遮荫对蒸腾速率日变化的影响
2.4遮荫对胞间CO2摩尔分数日变化的影响
斑叶堇菜和大叶铁线莲的胞间CO2摩尔分数日变化与各自的净光合速率日变化相比,呈现出了不同的变化趋势,各处理均表现出了先下降后上升的趋势,在12:00时,胞间CO2摩尔分数达到了一天中的最低值。且随着遮荫度的增加,2种植物在各遮荫处理条件下的值均比全光照依次降低(图4)。
2.5净光合速率与气孔导度以及蒸腾速率的关系
Pn与Gs间和Pn与Tr间均表现为线性关系。
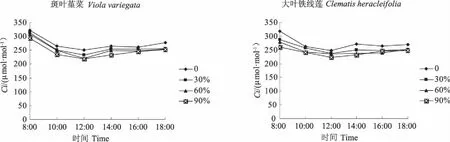
图4 遮荫对胞间CO2摩尔分数日变化的影响
不同的遮光条件对2种地被植物的Pn与Gs之间和Pn与Tr之间的相关性都没有影响,大叶铁线莲的Pn与Gs,Pn与Tr均呈显著相关(P<0.05);斑叶堇菜的Pn与Tr也呈显著相关(P<0.05),但其Pn与Gs却没有明显的相关性。大叶铁线莲的Pn与Tr相关性较强,全光照与30%、60%、90%遮荫条件下分别达到 0.97、0.92、0.99、0.82(表1)。
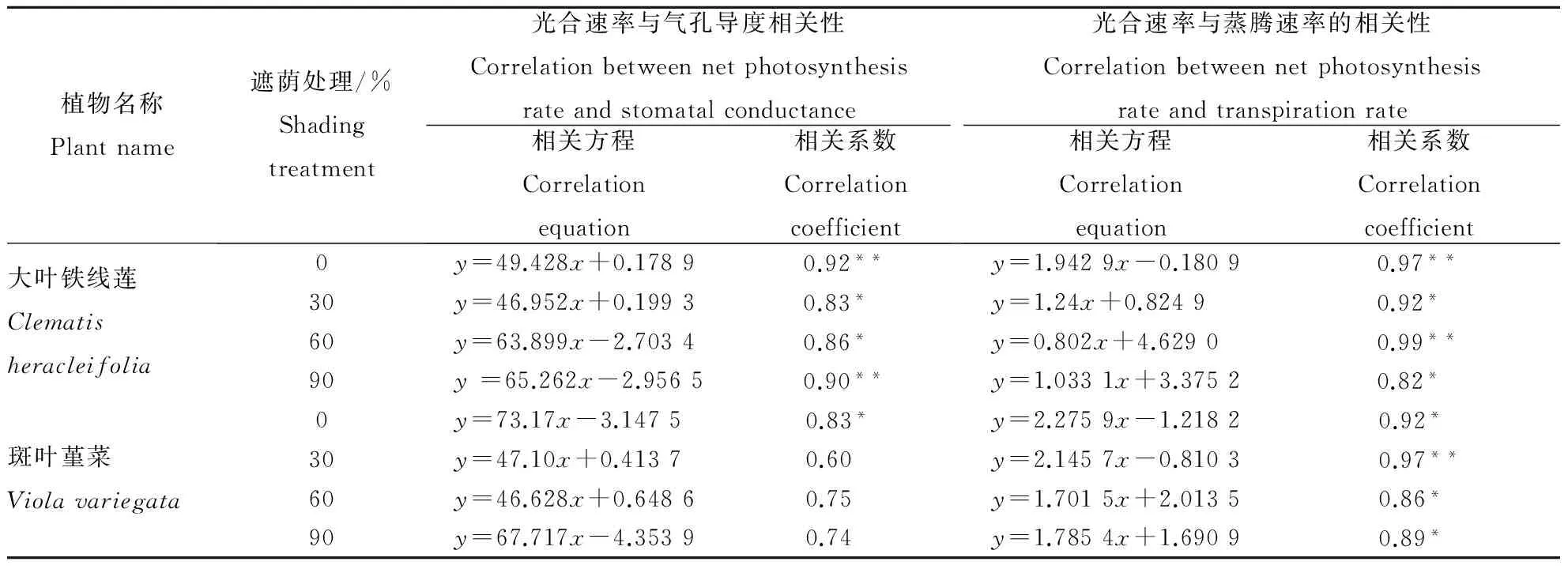
表1 光合速率与气孔导度和蒸腾速率之间的相互关系Table 1 Correlation between net photosynthesis rate and the stomatal conductance and transpiration rate
注:*表示显著性相关(P<0.05),**表示极显著性相关(P<0.01)。
Note:*Correlation is significant at the level of 0.05,**Correlation is extremely significant at the level of 0.01.
2.6遮荫对光响应曲线的影响
由图5可知,各遮荫处理下大叶铁线莲和斑叶堇菜的净光合速率对光强的响应规律基本与全光照处理下的一致,均有快速增加、缓慢上升与下降阶段:在光强小于500 μmol·m-2·s-1时,净光合速率随着光合有效辐射的增加而快速增加;在光强介于500 μmol·m-2·s-1和各自的光饱和点之间时,净光合速率的增加程度随着光合有效辐射的增加逐渐变缓;当光合有效辐射达到各自的光饱和点时,净光合速率的值达到最大;之后当光合有效辐射继续增加,叶片因为过强的光照而产生了光抑制,从而导致了净光合速率的下降。且随着遮荫度的增加,大叶铁线莲和斑叶堇菜各遮荫处理的曲线与对照相比依次降低,光合速率也均依次低于全光照下的。
从表2可以看出,遮荫对2种地被植物的光饱和点、光补偿点、光饱和时的最大净光合速率以及暗呼吸速率均有较为显著的影响。大叶铁线莲和斑叶堇菜的光饱和点、光补偿点、最大净光合速率以及暗呼吸速率均随着遮荫度的增加而依次减小。其中,大叶铁线莲在遮荫90%时的光饱和点、光补偿点、最大净光合速率以及暗呼吸速率与全光照的相比分别下降了32.07%、13.26%、35.88%、19.91%;斑叶堇菜则分别下降了36.18%、51.91%、44.27%、46.79%。而2种植物的表现量子效率随着遮荫度的增加则呈现了逐渐增加的趋势。这说明在弱光条件下,2种地被植物可通过降低光饱和点、光补偿点和暗呼吸速率,提高光量子效率的方式来提高利用弱光的能力,同时降低呼吸消耗,提高光合效率,从而保证植株能够正常生长,以表现出较强的适应弱光的能力。
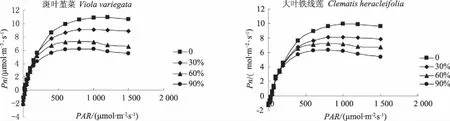
图5 遮荫对光响应曲线的影响Fig.5 Effects of shading on the light response curves
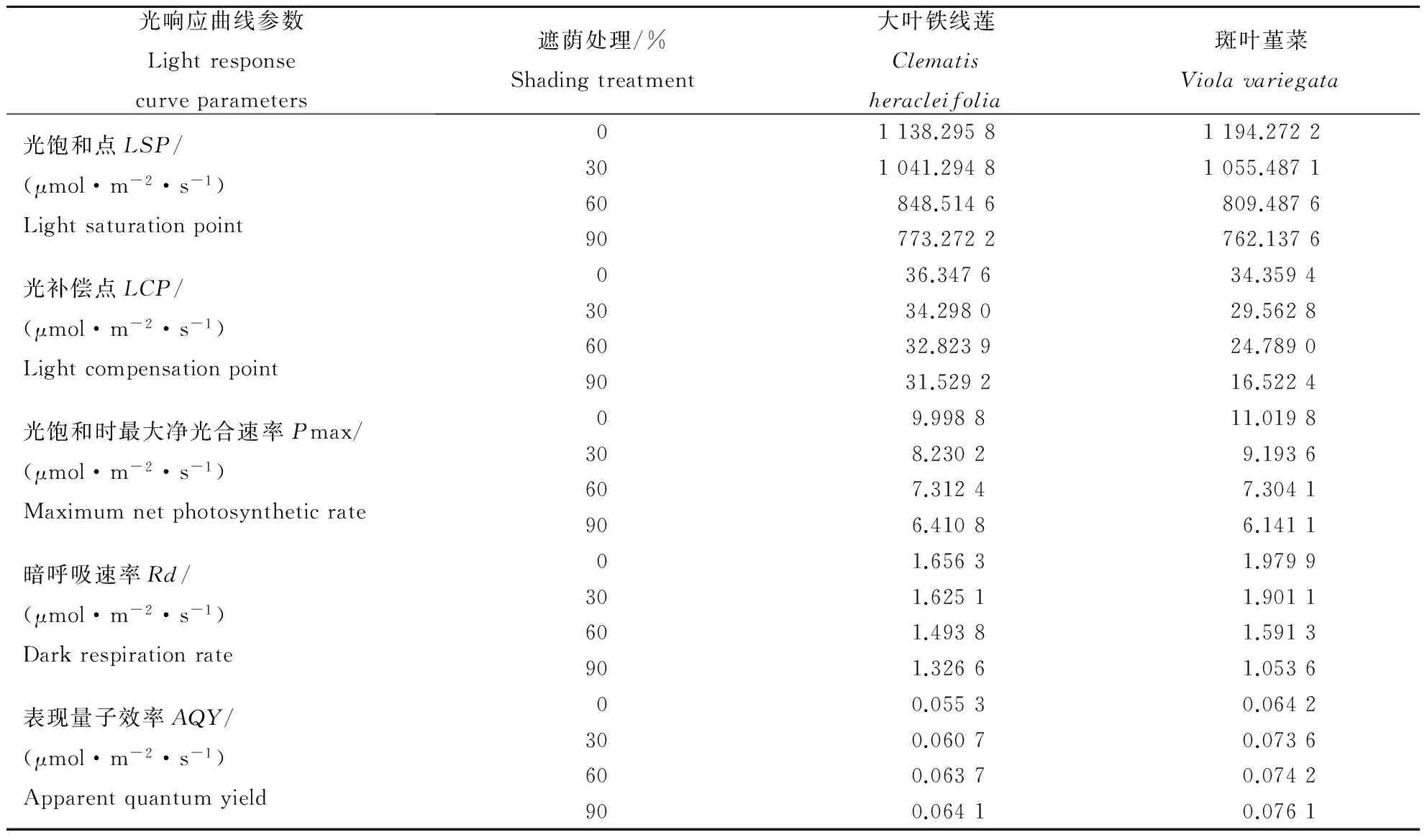
光响应曲线参数Lightresponsecurveparameters遮荫处理/%Shadingtreatment大叶铁线莲Clematisheracleifolia斑叶堇菜Violavariegata光饱和点LSP/(μmol·m-2·s-1)Lightsaturationpoint01138.29581194.2722301041.29481055.487160848.5146809.487690773.2722762.1376光补偿点LCP/(μmol·m-2·s-1)Lightcompensationpoint0 36.3476 34.35943034.298029.56286032.823924.78909031.529216.5224光饱和时最大净光合速率Pmax/(μmol·m-2·s-1)Maximumnetphotosyntheticrate0 9.9988 11.0198308.23029.1936607.31247.3041906.41086.1411暗呼吸速率Rd/(μmol·m-2·s-1)Darkrespirationrate0 1.6563 1.9799301.62511.9011601.49381.5913901.32661.0536表现量子效率AQY/(μmol·m-2·s-1)Apparentquantumyield0 0.0553 0.0642300.06070.0736600.06370.0742900.06410.0761
2.7遮荫对CO2响应曲线的影响
图6表明,在经过遮荫处理后,斑叶堇菜和大叶铁线莲对CO2的响应都与它们在全光照条件下的响应基本一致。随着CO2浓度的增加,2种地被植物在CO2没有到饱和点以前,各处理的净光合速率均逐渐上升,达到CO2饱和点后,净光合速率又有所下降,且随着遮荫度的增加,大叶铁线莲和斑叶堇菜各遮荫处理的曲线与对照相比依次降低,光合速率也均依次低于全光照下的。
表3进一步说明,遮荫对2种地被植物的CO2饱和点、CO2补偿点、CO2饱和时的最大净光合速率影响均较为显著。大叶铁线莲和斑叶堇菜的CO2饱和点、CO2补偿点、最大净光合速率、光呼吸速率随着遮荫度的增加均依次减小。其中大叶铁线莲在遮荫90%时的CO2饱和点、CO2补偿点、CO2最大净光合速率与全光照的相比分别下降了23.04%、23.19%、25.78%;斑叶堇菜则分别下降了20.38%、24.66%、22.03%。而2种植物的羧化效率随着遮荫度的增加则呈现了逐渐增加的趋势。这说明在弱光条件下,2种地被植物可通过降低CO2饱和点、CO2补偿点,抑制光呼吸和提高羧化效率的方式来提高利用弱光的能力,并提高光合效率,从而保证植株能够正常生长,以表现出较强的适应弱光的能力。
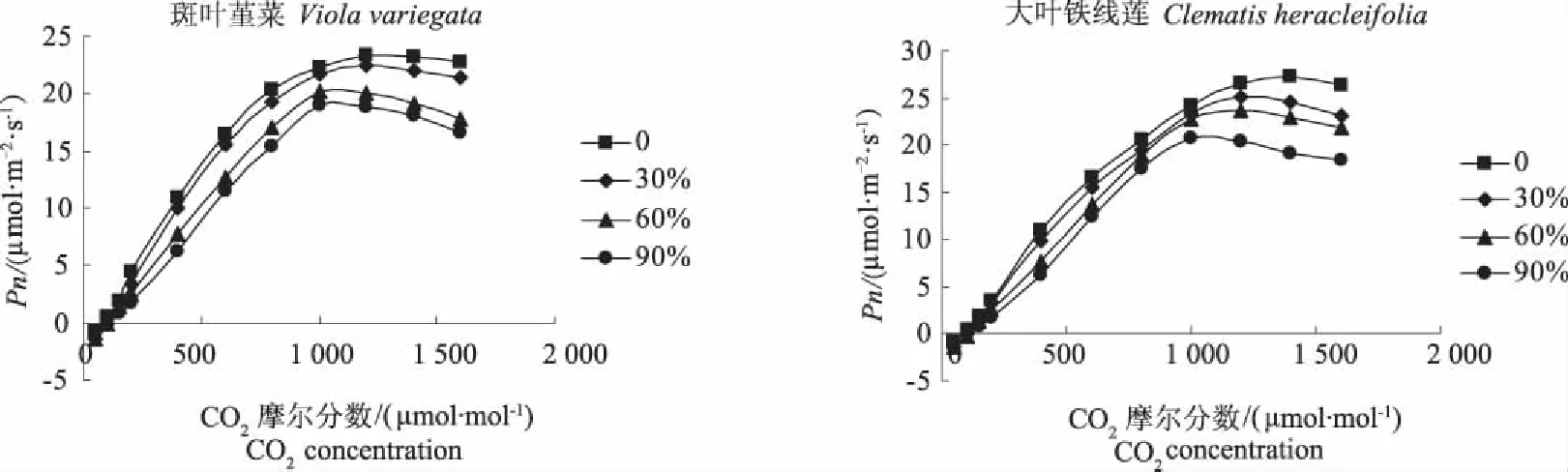
图6 遮荫对CO2响应曲线的影响Fig.6 Effects of shading on the CO2 response curves
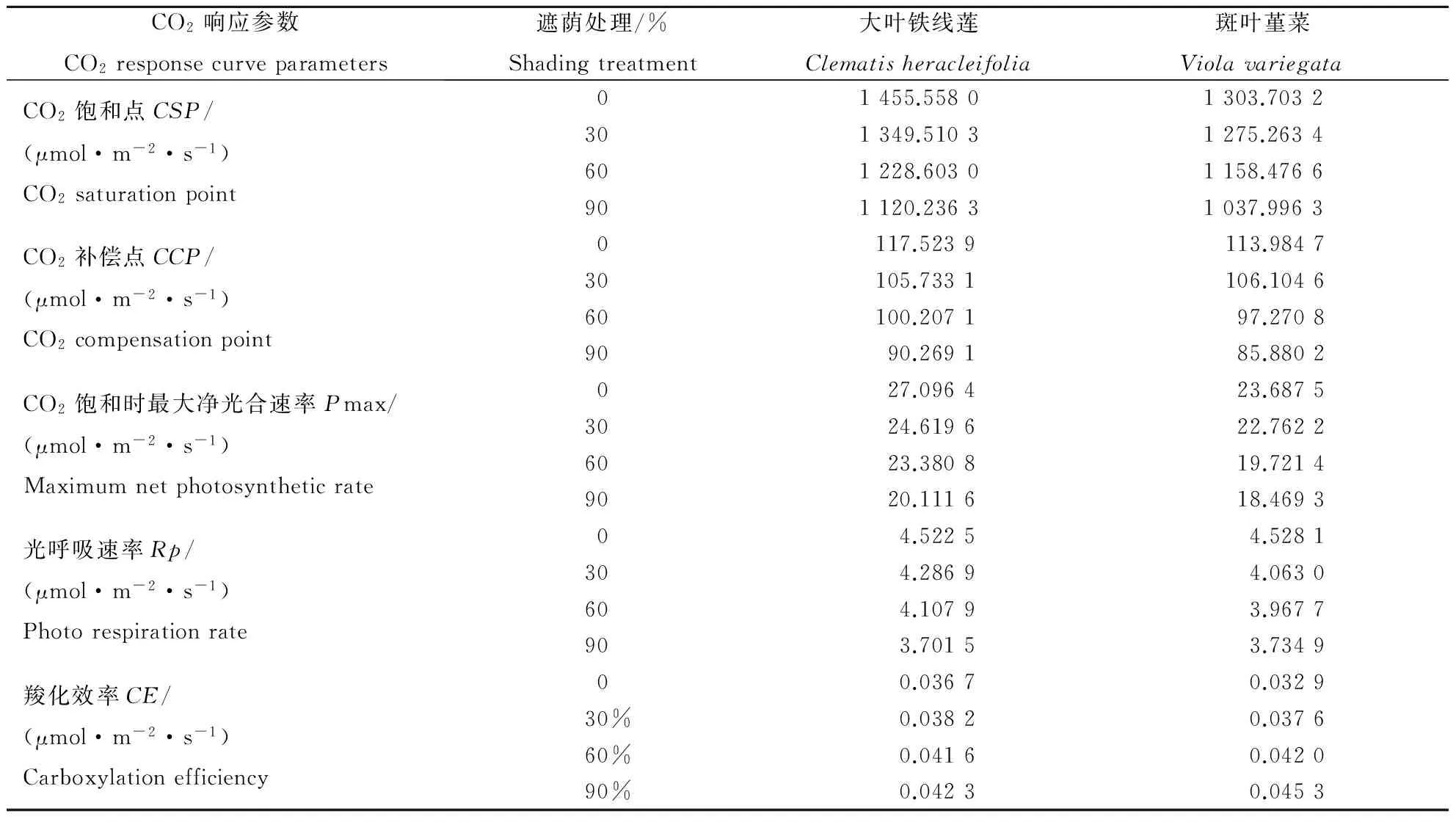
CO2响应参数CO2responsecurveparameters遮荫处理/%Shadingtreatment大叶铁线莲Clematisheracleifolia斑叶堇菜ViolavariegataCO2饱和点CSP/(μmol·m-2·s-1)CO2saturationpoint01455.55801303.7032301349.51031275.2634601228.60301158.4766901120.23631037.9963CO2补偿点CCP/(μmol·m-2·s-1)CO2compensationpoint0117.5239 113.984730105.7331106.104660100.207197.27089090.269185.8802CO2饱和时最大净光合速率Pmax/(μmol·m-2·s-1)Maximumnetphotosyntheticrate0 27.0964 23.68753024.619622.76226023.380819.72149020.111618.4693光呼吸速率Rp/(μmol·m-2·s-1)Photorespirationrate0 4.5225 4.5281304.28694.0630604.10793.9677903.70153.7349羧化效率CE/(μmol·m-2·s-1)Carboxylationefficiency0 0.0367 0.032930% 0.03820.037660% 0.04160.042090% 0.04230.0453
3 结论与讨论
随着遮荫度的增加,斑叶堇菜和大叶铁线莲的净光合速率、气孔导度、蒸腾速率、胞间CO2浓度、光饱和点、光补偿点、CO2饱和点、CO2补偿点降低,表现量子效率和羧化效率随遮荫度的增加而提高。不同遮荫条件下,2种植物的净光合速率日变化、气孔导度日变化、蒸腾速率日变化均为双峰曲线,表现出明显的“午休”现象。在弱光条件下,2种地被植物均可通过自身的调节来提高利用弱光的能力。综合分析可得:斑叶堇菜和大叶铁线莲均具有较强的耐荫性。
植物光合作用变化的不同程度是不同植物所具有的特性[8-9]。通常,净光合速率日变化可分为“单峰”型、“双峰”型和“三峰”型等类型。本研究中,不同遮荫下斑叶堇菜和大叶铁线莲的净光合速率日变化均表现为典型的“双峰”型变化规律,出现了明显的“午休”现象,可见遮荫只改变了每日各时刻的净光合速率数值,并未改变净光合速率日变化规律,这与严海燕等[10]的研究结果一致。许大全等[11]把非气孔限制和气孔限制作为了植物出现光合“午休”现象的两大因素。本研究中,2种植物的胞间CO2浓度和净光合速率在10:00—12:00间的变化相同,都呈现下降趋势,由此可知,斑叶堇菜和大叶铁线莲出现光合“午休”现象是由气孔限制因素导致的。
光补偿点和光饱和点作为植物耐荫性评价的重要指标,其高低直接反映了植物对弱光的利用能力[12]。一般LCP和LSP均较低是典型的耐荫植物,能充分地利用弱光进行光合作用[13]。本研究中,斑叶堇菜和大叶铁线莲的光饱和点和光补偿点随着遮荫度的增加而逐渐下降,说明2种植物对弱光的利用能力较强,具有较强的耐荫性。表观量子效率是植物利用光能情况的反映。表观量子效率越大,表明植物对弱光的利用能力强,吸收与转换光能的色素蛋白复合体多[14]。薛建平等[15]研究表明,遮荫后叶片的表观量子效率与全光照相比显著上升。本研究也表明,遮荫度的增加提高了斑叶堇菜与大叶铁线莲的表现量子效率。
CO2作为光合作用的原料,也在较大程度上影响着植物的光合作用,通常认为达到光饱和点之后,对植物光合速率影响较大的因素是CO2[16]。董如磊[17]研究表明,白辛树在经过遮荫处理后,CO2补偿点降低。本研究中,斑叶堇菜和大叶铁线莲的CO2补偿点也随着遮荫度的下降而降低,说明遮荫环境提高了对2种植物利用CO2的能力,从而使有机物在较低的CO2浓度下就开始有效积累。羧化效率是植物同化CO2的能力的反映,羧化效率越大,则表示在较低的CO2浓度下有较高的光合速率[18]。本研究中,斑叶堇菜和大叶铁线莲在遮荫条件下的羧化效率要大于全光照条件下的羧化效率,说明遮荫后,2种植物在低浓度的CO2条件下有较高的光合速率,适应弱光的能力较强。
[1] 缴丽莉,路丙社,周如久,等. 遮光对青榨槭光合速率及叶绿素荧光参数的影响[J].园艺学报,2007,34 (1):173-178.
[2] 鲍平秋.斑叶堇菜[J].中国花卉盆景,2005(4):4.
[3] 叶子飘,王建林.植物光合-光响应模型的比较分析[J].井冈山学院学报(自然科学版),2009,30(4):9-13.
[4] 习朱万泽,土金锡,薛建辉.台湾桤木引种的光合生理特性研究[J]西北植物学报,2004,24(11): 2012 -2019.
[5] 王瑞,丁爱萍,杜林峰,等.遮荫对 12 种阴生园林植物光合特性的影响[J].华中农业大学学报(自然科学版),2010,29(3):369-374.
[6] 赵玲珍,朱首军.三种树莓光合特性的研究[J].西北林学院学报,2006, 21(4):39-42.
[7] 王玉辉,周光胜.松嫩草地羊草叶片光合作用生理生态特征分析[J].应用生态学报,2001,12(1):75-79.
[8] BJORLANAN O, HOHNGREN P. Adaptability of the photosynthetic apparatus to light intensity in ecotypes from exposed and shaded habitat[J].Physoil Plantarum,1963, 16( 4):889-914.
[9] GAUL E. Differential photosynthetic performance ofSolanumdulcamaraecotypes form shaded and exposed habitat[J].Carnegie Inst Yearbook,1967,67:482-487.
[10] 严海燕,董然,金光勋,等.遮荫对富贵草光合特性的影响[J].东北林业大学学报,2013,41(5):46-50.
[11] 许大全.光合作用气孔限制分析中的一些问题[J].植物生理学通讯,1997,33(4):241-244.
[12] 蔡仕珍,陈其兵,潘远智,等.遮光对花叶细辛光合特性和荧光参数的影响[J].四川农业大学学报,2004,22(4):326-331.
[13] 冷平生,杨晓红,胡悦,等.5种园林树木的光介和蒸腾特性的研究[J].北京农学院学报,2000,15(4):13-18.
[14] 潘瑞炽,王小菁,李娘辉.植物生理学[M].北京:高等教育出版社,2004:87-90.
[15] 薛建平,王兴,张爱民,等.遮荫对半夏光合特性的影响[J].中国中药杂志,2008,33(24):2896-2900.
[16] 蔡仕珍,李西,潘远智,等.不同光照对蝴蝶花光合特性及生长发育研究[J].草业学报,2013,22(2):264-272.
[17] 董如磊.东京野茉莉耐荫性研究[D].南京:南京林业大学,2010.
[18] 刘金祥,陈伟云,肖生鸿.黑籽雀稗的光介生理特性研究[J].草业学报,2009,18(6): 254-258.
(责任编辑:蒋国良)
EffectsofshadingonthephotosyntheticcharacteristicsofViolavariegataandClematisheracleifolia
LIU Kaiyue, DU Shaohua, GUO Sijia, ZHAO Shuo
(College of Horticulture, Forestry and Tourism, Hebei Agricultural University, Baoding 071000, China)
To explore the influence of shading on photosynthetic characteristics inViolavariegataandClematisheracleifolia, with full illumination as the control, these two plants were treated at three shading densities: 30%, 60% and 90%. The results showed that with the rise of shading density, the net photosynthetic rate, stomatal conductance, transpiration rate, CO2concentration between cells, light saturation point, light compensation point, CO2saturation point and CO2compensation point of Viola Variegata and Clematis Heracleifolia decreased. But the performance quantum efficiency and carboxylation efficiency increased with the rise of shading density. The day range of net photosynthetic rate, stomatal conductance and transpiration rate of these two plants show bimodal curve and obvious “midday depression” (under different shading densities). This indicates that the shade endurance ofViolavariegataandClematisheracleifoliais strong.
Violavariegata;Clematisheracleifolia; shading density; photosynthetic characteristics
2016-04-23
河北省科技厅科技攻关项目(15227534,14236001D,16236901D-5)
刘凯月(1991-),女,河北石家庄人,硕士研究生,从事园林植物应用方面的研究。
刘冬云(1971-),女,河北赞皇人,副教授,博士。
1000-2340(2016)06-0778-07
S687
:A
