东北堇菜属植物叶形态结构的研究及其分类学价值的探讨
2014-11-07孙雪芹刘玫孙天航张欣欣史传奇
孙雪芹,刘玫,孙天航,张欣欣,史传奇
(哈尔滨师范大学生命科学与技术学院,黑龙江 哈尔滨 150025)
堇菜属(Viola)是堇菜科(Violaceae)最大的属,约500余种,世界分布。中国约120种,东北42种、1变种及4变型[1-3]。本属植物用途广泛,可供药用,有清热解毒、消肿止痛及散瘀之功效[如紫花地丁(Violaphilippica)及鸡腿堇菜(Violaacuminata)],也可供观赏[如三色堇(Violatricolor)][4-5]。
堇菜属植物的柱头形态、花色和托叶常被作为分类的主要依据[1,6-7]。中国堇菜属分为4亚属,即堇菜亚属(Subgen.Viola)、二裂花柱亚属(Subgen.Dischidium)、须毛柱头亚属(Subgen.Chamaemelanium)及美丽堇菜亚属(Subgen.Melanium)[1]。孙坤等[8]对38种国产堇菜属植物进行了数量分类研究,认为须毛柱头亚属和二裂花柱亚属应作为同一亚属,堇菜亚属中裂叶堇菜组(Sect. Pinnatae)和阔鞘堇菜组(Sect. Vaginatae)应并入合生托叶组(Sect. Adnatae)。张春宇等[9]指出紫花地丁和早开堇菜(Violaprionantha)根的木质部、叶片的栅栏组织及海绵组织等结构明显不同。叶表皮的微形态特征不支持将早开堇菜及茜堇菜(Violaphalacrocarpa)合并到紫花地丁,不支持将总裂叶堇菜降为裂叶堇菜的变种[10]。堇菜属多数种类的花粉粒为三孔沟,其中三色堇的花粉为五孔沟,是本属内较为进化的类型[11-12]。一些学者指出堇菜属果实和种皮表面纹饰在种间差别明显,可作为种的鉴定依据[13-14]。而分子系统学研究显示美丽堇菜亚属与堇菜组和鸟嘴柱头堇菜组有较近亲缘关系,二裂花柱亚属与须毛柱头亚属近缘[15]。叶柄的解剖学特征对植物分类及系统演化研究,以及鉴定植物属种间的亲缘关系起到重要作用,对属及种的鉴定有着很大的参考价值[16-17]。然而,关于堇菜属植物叶柄结构的报道很少[18],且无系统的解剖学研究。本文的目的是通过深入研究东北堇菜属植物叶的形态结构,完善堇菜属形态学,为该属的分类及分子系统学研究提供形态学依据,并为进一步深入研究中国堇菜属及堇菜科提供了基础资料。
1 材料与方法
1.1 材料
实验材料为我国东北堇菜属28种及1变种的叶片及叶柄,它们分别隶属于堇菜属的4个亚属:美丽堇菜亚属及二裂花柱亚属各1种,须毛柱头堇菜亚属3种,其他均属于堇菜亚属。植物采自野外,经王臣教授鉴定,标本存放于哈尔滨师范大学植物标本馆(HANU),部分材料取自于黑龙江植物资源所标本馆(HNR)以及东北农业大学植物标本馆(NEAU)。物种名称及凭证标本(包括物种的采集人,采集号及存放的标本馆)的详尽信息见表1,分类地位见表2。

表1 东北堇菜属物种、凭证标本及采集地Table 1 Taxa of Viola in northeastern China, together with voucher specimen details and localities
1.2 方法
叶表皮撕取法:分别取5个叶片,用镊子撕取其上、下表皮,放在载玻片上,用刀片轻轻刮去残留的叶肉,直至可清楚地观察到上、下表皮细胞为止。用水冲去叶肉细胞,加入1~2滴50%甘油水溶液,盖上盖玻片。在显微镜下观察表皮细胞、气孔及毛状体的形态并记录。
GMA(Glycol methacrylate,乙二醇甲基丙烯酸酯)半薄切片法:将从标本上取的材料吸水膨胀固定于FAA(Formalin-acetic acid-alcohol mixture,福尔马林-醋酸-酒精混合液),新鲜材料直接用FAA固定,时间不少于24 h,取靠近托叶的叶柄,长约0.5 cm。采用Feder和O’Brien[19]的方法,材料经50%酒精(2次,每次4~6 h),100%酒精(2次,每次4~6 h),异丙醇(2次,每次6 h)和正丁醇(2次,每次6 h)脱水后,进入GMA渗透3次,第1和2次各为1~2 d,第3次不少于5 d,之后将材料及GMA装入胶囊,将其置于60℃温箱24 h。用Leica Ultralcut R切片,厚度约为2 μm。同样采用Feder和O’Brien[19]的方法,用考马斯亮蓝及希夫试剂染色,中性树胶封片。
所有观察结果经Olympus BX51显微镜观察,Olympus DP70照相。

表2 东北堇菜属物种的分类地位(中国植物志)Table 2 The taxonomic status of Viola in northeastern China (Flora of China)
2 结果与分析
2.1 叶及托叶的形态
东北堇菜属植物的叶具裂或无裂(表3),裂叶堇菜组的叶具裂,其他各组叶边缘较整齐。叶裂分为全裂,如掌叶堇菜(图1C),深裂(缺刻大于1/2叶片),如南山堇菜及裂叶堇菜(图1D),或浅裂(缺刻小于1/2叶片),如总裂叶堇菜(图1E)。后者的叶片形状变化较大,为心形或广卵形,如斑叶堇菜和鸡腿堇菜(图1A1,J1);肾形,如大黄花堇菜、兴安圆叶堇菜和双花堇菜(图1B);长卵形,如立堇菜、三色堇和紫花地丁(图1H,I,M);长三角形,如白花东北堇菜及宽叶白花堇菜(图1G)及匙形,如兴安堇菜(图1F)。
本文研究的堇菜属物种均具托叶(表3),但有的不明显。根据托叶与叶柄的基部着生方式可分为合生及离生两种类型。前者如鸡腿堇菜、兴安圆叶堇菜和掌叶堇菜(图1A1,B,C)。后者如立堇菜和三色堇(图1H,I)。托叶的形状主要为披针形及卵圆形。多数物种,如兴安堇菜、紫花地丁和斑叶堇菜的托叶为披针形(图1M,J2),少数物种,如黄花堇菜和大黄花堇菜的为卵圆形(图1L)。本文研究的物种仅三色堇的托叶具小裂片(图1I)。

图1 东北堇菜属植物叶及托叶绘图Fig.1 The leaves and stipules in Viola in northeastern China, showing the shape of leaves and stipules
示叶及托叶形状A1, A2. 鸡腿堇菜基生叶及茎生叶托叶;B. 兴安圆叶堇菜;C. 掌叶堇菜;D. 裂叶堇菜;E. 总裂叶堇菜;F. 兴安堇菜;G. 宽叶白花堇菜;H. 立堇菜;I. 三色堇;J1, J2. 斑叶堇菜;K. 堇菜;L. 黄花堇菜托叶;M. 紫花地丁。比例尺:图A1, A2, B, C, D, E, F, G, H, I, J1, K, M=1 cm;J2, L=0.5 cm。Basal leaf and stipule of stem leaf: A1, A2.V.acuminata; B.V.brachyceras; C.V.dactyloides; D.V.dissecta; E.V.fissifolia; F.V.gmeliniana; G.V.lactiflora; H.V.raddeana; I.V.tricolor; J1, J2.V.variegata; K.V.verecunda; L.V.xanthopetalastipule; M.V.yedoensis. Scale bar=1 cm in A1, A2, B, C, D, E, F, G, H, I, J1, K, M; 0.5 cm in J2, L.
2.2 毛状体及气孔的形态
除少数物种未见毛状体(如堇菜和宽叶白花堇菜)外,其余物种均有毛状体。毛状体多集中在叶脉及叶缘,少数叶柄也被毛。毛状体多为单细胞非腺毛,呈披针形,如茜堇菜、三色堇和双花堇菜(图2B,C)。仅在球果堇菜观察到两细胞非腺毛,呈披针形(图2A1,A2)。部分物种毛状体上具瘤状突起,如斑叶堇菜和三色堇(图2C)。气孔分布于叶的上、下两面及叶柄,下表皮气孔较多。绝大多数物种的气孔周围有3个副卫细胞,其中一个明显较其他的细胞小,为不等型,如早开堇菜和堇菜(图2D,E),部分物种气孔周围的细胞同其他表皮细胞形态相似,为不规则型,如深山堇菜、斑叶堇菜等(图2E)。堇菜属表皮细胞的细胞壁多呈波浪状弯曲,如堇菜(图2E),但有的物种较平滑,如总裂叶堇菜、早开堇菜(图2D)。
2.3 叶柄结构
堇菜属植物叶柄横切面多为半圆形(图3),如鸡腿堇菜、裂叶堇菜及总裂叶堇菜(图3A,C,D);三角形,如白花东北堇菜(图3F)。叶柄近轴面的特征在不同的物种有变化,分为较平,如总裂叶堇菜(图3D);中间凸起,如球果堇菜、茜堇菜及斑叶堇菜(图3B,H,J)。近轴面两侧向外延伸,形成翼,如鸡腿堇菜、球果堇菜和奇异堇菜(图3A,B,C);部分物种无明显翼,如总裂叶堇菜(图3E)。
叶柄表皮由一层近方形的细胞组成,无明显角质层,细胞排列紧密整齐,但东方堇菜叶柄一些表皮细胞的外壁明显突起(图2G)。此外,研究中叶柄的表皮细胞可见细小的绒毛状突起(如球果堇菜,图2C)。紧挨表皮有1~3层细胞构成的厚角组织,细胞排列紧密,且形态同表皮细胞相似。厚角组织以内的薄壁细胞较大,排列疏松,有明显的胞间隙,维管束规则分布在薄壁组织中,多数物种有3个维管束,大维管束在中间位置,小维管束位于两侧,如东北堇菜及紫花地丁(图3F,O)。有些物种的维管束多于3个,东北堇菜、白花东北堇菜和紫花地丁具4个维管束(图3F,O),兴安堇菜的维管束为6个,它们的排列方式均为中央一个较大,周围的较小。根据维管束形状不同,分为以下4种:新月形,如鸡腿堇菜和奇异堇菜(图3A,G);肾形,如裂叶堇菜、总裂叶堇菜和东北堇菜(图3C,D);箭头形,如茜堇菜和斑叶堇菜(图3H,J);马蹄形,如球果堇菜、堇菜和紫花地丁(图3B,F,I,K,L)。部分物种的韧皮部及木质部外侧具纤维,纤维为2~3层纤维细胞,如斑叶堇菜和双花堇菜(图2H,J),或几个纤维细胞聚集在一起,分散排列,如总裂叶堇菜及东北堇菜(图2I,3E,F),有些物种的韧皮部及木质部外侧无纤维,如鸡腿堇菜、球果堇菜和堇菜(图3A,C,N)。一些物种叶柄中央具溶生性通气组织,如堇菜和紫花地丁(图3K,L),而这种通气组织在多数物种中不存在。淀粉粒存在于一些物种靠近表皮的细胞,如东北堇菜及紫花地丁(图3O)。
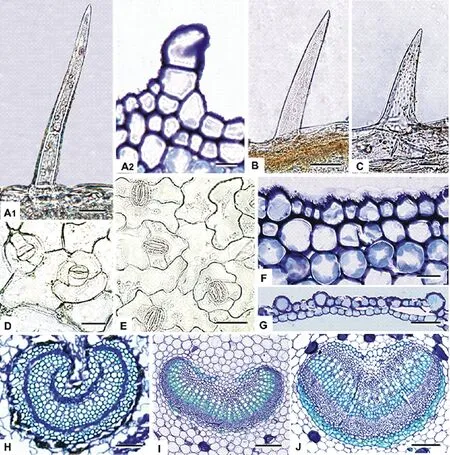
图2 东北堇菜属植物叶及叶柄Fig.2 The leaves and petiole in Viola in northeastern China
示毛状体、气孔、表皮、维管束、纤维。A1, A2. 球果堇菜的叶表皮毛;B. 双花堇菜的叶表皮毛;C. 三色堇的叶表皮毛;D. 早开堇菜气孔;E. 堇菜气孔;F. 球果堇菜叶柄表皮;G. 东方堇菜叶柄表皮;H. 双花堇菜叶柄维管束;I. 总裂叶堇菜叶柄维管束;J. 斑叶堇菜叶柄维管束。比例尺: 图A1, B, J=80 μm;A2, C, D, E, F=25 μm;G=100 μm;H, I=50 μm。Showing trichome, stomata, epidermis, vascular and fiber. Ttrichome on leaves: A1, A2.V.collina; B.V.biflora; C.V.tricolor; Stomata of leaves: D.V.prionantha; E.V.verecunda; Epidermis of petiole: F.V.collina; G.V.orientalis; Vascular bundle of petiole: H.V.biflora; I.V.fissifolia; J.V.variegata. Scale bar=80 μm in A1, B, J; 25 μm in A2, C, D, E, F; 100 μm in G; 50 μm in H, I.
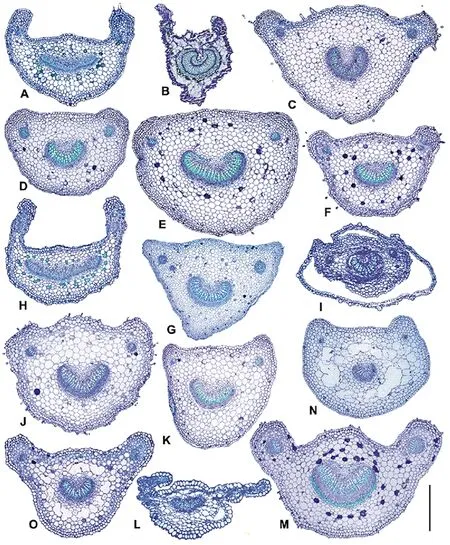
图3 东北堇菜属植物叶柄横切面绘图Fig.3 The transverse section of petioles in Viola in northeastern China
示横切面形状、维管束的分布及形状。A. 鸡腿堇菜;B. 双花堇菜;C. 球果堇菜;D. 裂叶堇菜;E. 总裂叶堇菜;F. 东北堇菜;G. 白花东北堇菜;H. 奇异堇菜;I. 东方堇菜;J. 茜堇菜;K. 早开堇菜;L. 三色堇;M. 斑叶堇菜;N. 堇菜;O. 紫花地丁。比例尺:图L=400 μm; A, B, H, I, O=500 μm; D, F, G, K, M, N=600 μm; C, E, J=670 μm。Showing the shapes of transverse sections, distribution and shapes of vascular bundles. A.V.acuminata; B.V.biflora; C.V.collina; D.V.dissecta; E.V.fissifolia; F.V.mandshurica; G.V.mandshuricavar.f.abliflora; H.V.mirabilis; I.V.orientalis; J.V.phalacrocarpa; K.V.prionantha; L.V.tricolor; M.V.variegata; N.V.verecunda; O.V.yedoensis. Scale bar=400 μm in L; 500 μm in A, B, H, I, O; 600 μm in D, F, G, K, M, N; 670 μm in C, E, J.
2.4 根据观察的堇菜属28种1变种叶的形态结构的特征,编制检索表如下

3 讨论
叶片作为植物体最重要的营养器官,是植物进化过程中对环境变化比较敏感且可塑性较大的器官,与植物抗旱性密切相关[20-23],易受到水热等微环境的影响而发生变化[24]。而叶柄是植物比较保守和原始的器官[25],在鉴定植物种属间的亲缘关系中起到重要作用[26]。叶柄维管束的数目、位置及排列等是属组间比较有分类价值的解剖形状[27]。研究结果表明堇菜属植物翼、叶柄近轴面的特征、通气组织及维管束的结构等在不同物种之间有明显区别,可以作为分类的依据,其中叶柄维管束排列方式基本一致,是属内稳定的特征。这与堇菜属的花粉外壁纹饰研究结果一致[12],表明了本属是一个较为自然的类群。
研究表明美丽堇菜亚属花粉外壁纹饰及种皮表面纹饰与堇菜属其他物种差异明显,须毛柱头亚属的花粉外壁穿孔极多也较为特殊,支持将美丽堇菜亚属及须毛柱头亚属作为亚属的分类处理[8,11,13],但其他关于堇菜属的数量分类学研究则认为应将须毛柱头亚属和二裂花柱亚属作为同一亚属的不同组处理[12]。本文研究的位于须毛柱头亚属的东方堇菜其叶柄的一些表皮细胞明显突起,位于美丽堇菜亚属的三色堇的托叶具小裂片。由于这些特征不存在于研究的其他物种,故支持须毛柱头亚属及美丽堇菜亚属的建立。
堇菜亚属种类多,而且结构变化较大,植物叶表皮微形态可为属种间的分类提供依据[28]。本研究中涉及堇菜亚属6个组,其中二细胞非腺毛仅见堇菜组,显示堇菜组是一个特化的类群,支持王庆瑞[1]关于堇菜组的建立。裂叶堇菜组与合生托叶组叶的结构相似,均具单细胞非腺毛,不等型气孔,肾形及马蹄形维管束,应将裂叶堇菜组并入合生托叶组,这一结论也与数量分类、分子系统学和其他学者的研究结果一致[8,15,29]。
黄勇等[30]根据叶形变化认为早开堇菜和紫花地丁在形状上无差别,应合并。而早开堇菜、茜堇菜和紫花地丁叶表皮微形态,根叶解剖结构,花粉形态及果实和种子的形态上均存在着明显的差异,应为3个不同的种[9-10,14,31]。本研究揭示的早开堇菜、茜堇菜和紫花地丁的气孔类型,维管束数量和形状,是否存在纤维等特征存在区别,故支持前人将他们作为独立的种。东北堇菜与白花东北堇菜的叶柄横切面形状存在着较大的差异,因此对后者是否为独立种需要进一步的深入研究。此外,叶柄维管束的数量及排列方式在组内具有稳定性,可作为划分组的形态依据。
我们的研究深入揭示了堇菜属植物叶,包括叶片、托叶及叶柄的形态结构,这些结构在不同物种间存在差异,在组内具有相似的特征,为种及组的划分提供了形态学依据。完善了堇菜属的形态学研究,为进一步深入研究中国堇菜属及堇菜科提供了基础资料,从而也为分子系统学对这一类群的研究提供了形态学依据。
