中国云南8种堇菜属植物的叶形态解剖特征及分类学意义
2023-05-21邵成艳段涵宁杨晨璇
田 琴 邵成艳 段涵宁 杨晨璇,4 李 璐*
(1.西南林业大学生物多样性保护学院,昆明 650224;2.西南林业大学林学院,昆明 650224 ;3.西南林业大学园林园艺学院,昆明650224;4.中国科学院昆明植物研究所,昆明 650201)
堇菜属(Viola)是堇菜科(Violaceae)最大的一个属,有525~600 余种,为世界广布属,几乎广泛生长在世界无霜区[1]。它起源于安第斯山脉,分布于世界温带地区以及热带亚热带的高海拔山区,集中在欧洲的阿尔卑斯山和地中海、亚洲的喜马拉雅和东亚地区,以及南美洲的安第斯山脉和巴塔哥尼亚等地[1-3]。该属为多年生草本植物,部分种类具有较高观赏价值[1-4],如:著名的三色堇(V.tricolor)和七星莲(V.diffusa),也有一些药用植物,如:紫花地丁(V.philippica)和荁(V.moupinensis)。
长期以来,由于物种多、分布广、生态型明显、兼具开放花和闭锁花,且普遍具有自然杂交现象,堇菜属的物种鉴定和属下分类系统争议较大。1823年,Gingins[5]根据柱头形态把堇菜属分为5个组。1925年,Becker[6]基于雌蕊的超微结构后把它细分为14 组28 亚组及7 系。紧接着,Clausen[7-10]在此基础上,不断扩充修订该分类系统,明确指出花瓣颜色、雌蕊形态、染色体数目及地理分布类型等都是属下分组的重要依据。1996 年,Ballard[11]根据形态学、染色体数目和自然杂交现象等提出堇菜属可以分为8 个组,并具相应的地理分布格局,同时指出哪些组是存疑类群。近年来,分子系统学为探讨堇菜属的分类学系统提供了新证据。Ballard 等[1]选择了堇菜属下大部分组的44 个代表种为材料,基于内转录组间隔区序列(ITS 序列)构建了分子系统树,证实该属起源于南美安第斯山脉,揭示了属下分组类群并非单系。基于8个叶绿体基因片段构建的韩国堇菜属分子系统学表明,该地区的二裂柱头堇菜组和须毛柱头堇菜组(sectionsDischidium&Chamaemelanium)为单系,而其他组为并系,建议根据形态特征在组下进一步细分[12]。同样地,中国堇菜属分类系统也存在类似问题。我国堇菜属先记录有111种,分为4亚属14组[2],后被修订为96种(含35个中国特有种),分为5 亚属,但未进一步分组[3]。目前,我国堇菜属的分子系统学研究资料不多,主要是基于ITS片段构建的分子系统树,并结合形态学证据展开讨论,认为中国堇菜属不宜分为4个亚属,堇菜亚属不是单系,讨论了部分组间的亲缘关系及形态特征演化趋势[13-14]。
现有资料表明,堇菜属的叶表皮细胞形态、毛被特征、气孔类型、叶和叶柄的横切面解剖结构等在种间差异显著,为近似种的划分和属内组间分类提供了形态学证据,但大部分研究集中在伊朗[15-17]、北美[18]和夏威夷地区[19]、澳大利亚[20-21]、我国东北及东部地区的一些种类[22-25]。这些研究说明对于种类较多、分布广,且分类困难的堇菜属,以某个地域的分布物种为研究对象,不断积累资料,对于全面理解其分类系统是有帮助的。云南是中国堇菜属物种最丰富的地区,占了全国近一半以上的种类,约有56 种及4 变种[26],但该地区的相关研究资料较少。野外调查中,作者发现云南常见的一些堇菜属植物的花形态较为相似,容易混淆,也常被列为鉴定困难的近似种,包括产于滇西北开黄花的灰叶堇菜(V.delavayi)和双花堇菜(V.biflora),在滇中地区开紫花的紫花地丁、早开堇菜(V.prionantha)和香堇菜(V.odorata)等。同时,笔者观察到叶型在这些常见种间变化明显,可用于物种鉴定,值得深入研究。例如,卵形叶最常见,见于紫花地丁、早开堇菜、香堇菜、七星莲和灰叶堇菜,但前4 种被放置在堇菜亚属(subgen.Viola),而灰叶堇菜为二裂花柱亚属(subgen.Dischidium)。肾形叶、戟形叶和三裂叶等叶型较为少见,分别见于双花堇菜、长萼堇菜(V.inconspicua)和南山堇菜(V.chaerophylloides),在当地堇菜属里物种辨识度较高。
综上所述,本研究选取上述云南常见的8种堇菜属植物为研究对象(见表1),采纳中国堇菜属分为4亚属14组的分类系统[2],通过醋酸离析法和石蜡切片技术,在显微镜下观察其叶表皮形态、叶片和叶柄横切面解剖结构,比较种间的叶形态差异,为该属的物种分类提供新资料。
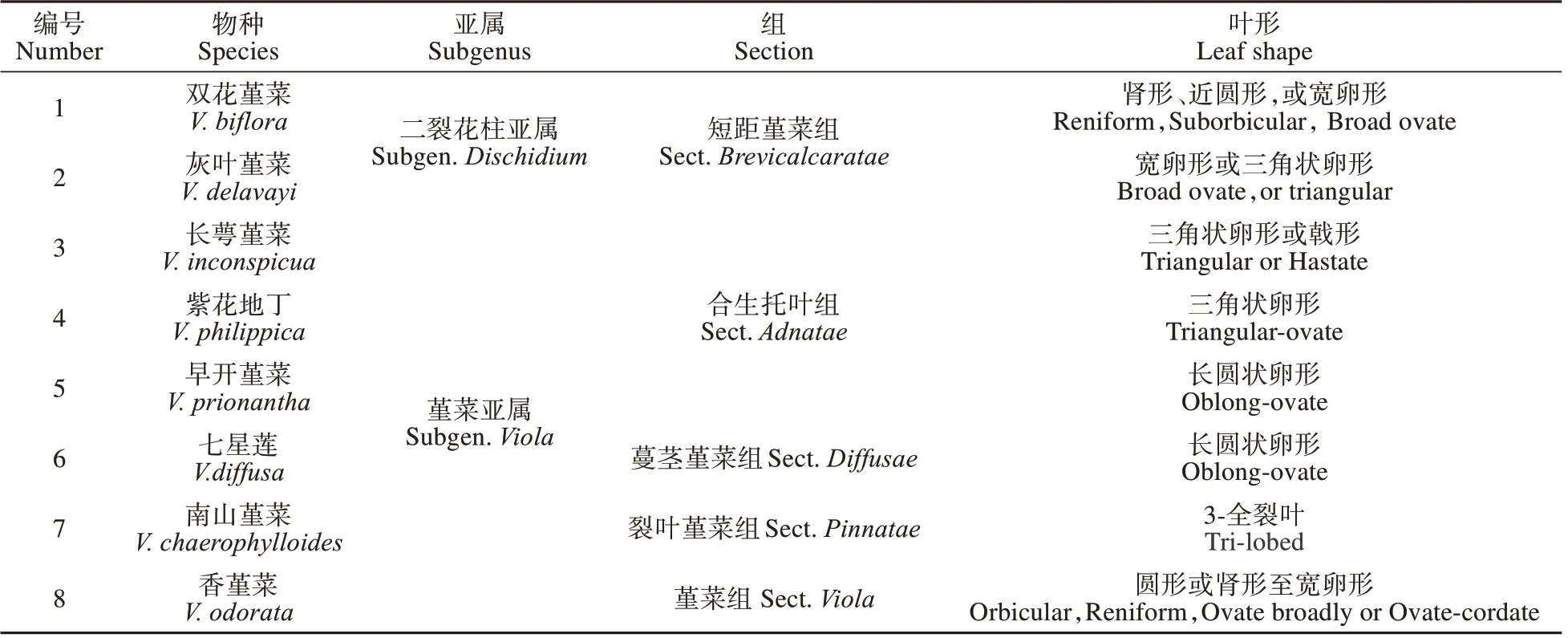
表1 8种堇菜属植物的基本信息[2]Table 1 Data of eight Viola species observed[2]
2 材料与方法
2.1 材料
8 种堇菜属植物(见表1)采集于2020—2021年春夏花期。野外观察拍照记录植株、花和叶片的形态特征,采集成熟叶片固定于50%乙醇,常温保存。其中,灰叶堇菜和双花堇菜属于二裂花柱亚属的短距堇菜组,分别采自大理市鸡足山和丽江市黑龙潭公园。其余6 种代表了堇菜亚属的4个组,均采自昆明植物园。值得说明的是,灰叶堇菜为中国特有种(仅分布于云南、贵州和四川),香堇菜为国外引种栽培多年的观赏植物,其余6种皆为北半球广布种,在云南均有分布,生长在温带和亚热带地区的林缘、灌木、草地和溪边等[2]。
2.2 方法
2.2.1 醋酸离析法观察叶表皮形态
取固定后的叶片,沿中脉取1 cm×1 cm 的小块,置于盛有适量解离液(V(30%过氧化氢)∶V(醋酸)=1∶1)[27]的小坩埚中。在烘箱中60 ℃加热3~5 h,待叶表皮与叶肉组织分离后取出(必要时可用毛笔清除叶肉残留组织),清水漂洗2~3 次后,置于洁净的载玻片上并滴1~2 滴1%番红染液,染色1~3 min。盖上盖玻片,洗去浮色,制作成临时装片,在尼康显微镜(DM750,日本)下观察拍照。观察上、下叶表皮的毛被类型、表皮细胞和气孔类型及密度,利用Image J 软件测量它们的大小,随机测量记录30 个数据,求平均值。其中,气孔指数=气孔数量/(气孔数量+叶表皮细胞数量)。专业术语参照文献[28]。
2.2.2 石蜡切片法观察叶中脉和叶柄横切面结构
取固定后的叶片,沿中脉处取叶片小块(0.5 cm×0.5 cm)和距叶柄基部约1 cm 处截取0.5 cm 长的叶柄段,每个物种各取5份样品。样品经酒精梯度脱水、二甲苯透明、浸蜡包埋、常规石蜡切片(手动轮转式切片机,Leica RM2125 RTS,德国),切片厚度为8 µm。切片烘干后,经番红-固绿染色,加拿大树脂胶封片,晾干后,在光学显微镜(Leica DM 750,德国)下观察叶片和叶柄横切面特征,包括上、下表皮层、维管束轮廓、叶肉和栅栏组织等;利用Image J软件测量它们的数据。
3 结果
3.1 8种堇菜属植物的植株形态
8 种堇菜属植物在开花期时的植株形态特征差异明显,包括生长习性、叶型及花形态等(图版Ⅰ)。首先,生长习性包括1年生和多年生草本,前者出现在七星莲(图版Ⅰ:6),后者出现在其余7个种。其次,成熟叶片几乎光滑无毛,叶形态在种间变化明显,可分为4 种:卵形叶(Ovate-shaped)、肾形叶(Reniform-shaped)、戟形叶(Hastate-shaped)和三全裂叶(Tri-lobed)。其中,卵形叶最普遍,出现在4 种:灰叶堇菜(图版Ⅰ:2)、紫花地丁(图版Ⅰ:5)、早开堇菜(图版Ⅰ:4)和香堇菜(图版Ⅰ:7)。相反,肾形叶、戟形叶和三全裂较为独特,分别见于双花堇菜(图版Ⅰ:1)、长萼堇菜(图版Ⅰ:3)和南山堇菜(图版Ⅰ:8)。再次,花色变异在组间有一定规律,可分为黄花、紫花和白花。黄花出现在二裂花柱亚属的双花堇菜和灰叶堇菜(图版Ⅰ:1~2);紫花出现在堇菜亚属的3组4种,包括长萼堇菜(图版Ⅰ:3)、早开堇菜(图版Ⅰ:4)、紫花地丁(图版Ⅰ:5)、七星莲(图版Ⅰ:6)和香堇菜(图版Ⅰ:7);而白花仅出现在南山堇菜(图版Ⅰ:8)。

图版Ⅰ 8种堇菜属植物的植株形态1.双花堇菜;2.灰叶堇菜;3.长萼堇菜;4.早开堇菜;5.紫花地丁;6.七星莲;7.香堇菜;8.南山堇菜Plate Ⅰ Plant morphology of eight Viola species1.Viola biflora;2.V.delavayi;3.V.inconspicua;4.V.prionantha;5.V.philippica;6.V.diffusa;7.V.odorata;8.V.chaerophylloides
3.2 8种堇菜属植物的叶表皮特征
3.2.1 毛被特征
肉眼观察下,8种植物在开花期成熟叶片几乎光滑无毛。但在显微镜下,除双花堇菜、长萼堇菜和香堇菜3 种的叶下表皮无毛外,其余5 种的叶片上下表皮均观察到长度不等、分布在叶缘和叶脉上的单列-单细胞-毛状体(uniseriately singlecelled trichome)(图版Ⅱ)。毛状体的长度在同种植物叶上下表皮的变化不大,但在种间差异明显,最长的出现在灰叶堇菜的上(903.69 µm)、下表皮(676.39 µm),最短的出现在南山堇菜的上(62.86 µm)、下表皮(136.94 µm)。根据长度,叶表皮毛被可分为三类:短柱状毛、中柱状毛和长柱状毛。第一类短柱状毛,是指毛状体长度小于150 µm,见于2 个种,包括南山堇菜的上、下表皮,密集在叶脉及叶缘(图版Ⅱ:1~2)和香堇菜的上表皮(67.14 µm)的叶缘处(图版Ⅱ:3)。第2类中柱状毛,是指毛状体长度在150~300 µm,见于3种植物,包括双花堇菜和长萼堇菜的上表皮(分别为249.45、177.61 µm),前者密集分布于叶缘细齿处(图版Ⅱ:5),后者均匀分布(图版Ⅱ:6),以及紫花地丁的上(219.61 µm)、下表皮(162.88 µm),集中分布于叶缘及叶脉(图版Ⅱ:4)。第3 类长柱状毛,是指毛状体长度大于300 µm,见于3 种,包括七星莲的上(310.82 µm)、下表皮(630.46 µm)(图版Ⅱ:8),灰叶堇菜的上(903.69 µm)、下表皮(676.39 µm)(图版Ⅱ:9)。但早开堇菜的上下表皮的毛被长度略有差异,下表皮为中柱状毛(201.49 µm),上表皮为长柱状毛(311.89 µm)(图版Ⅱ:7),密集分布在叶脉及叶缘。
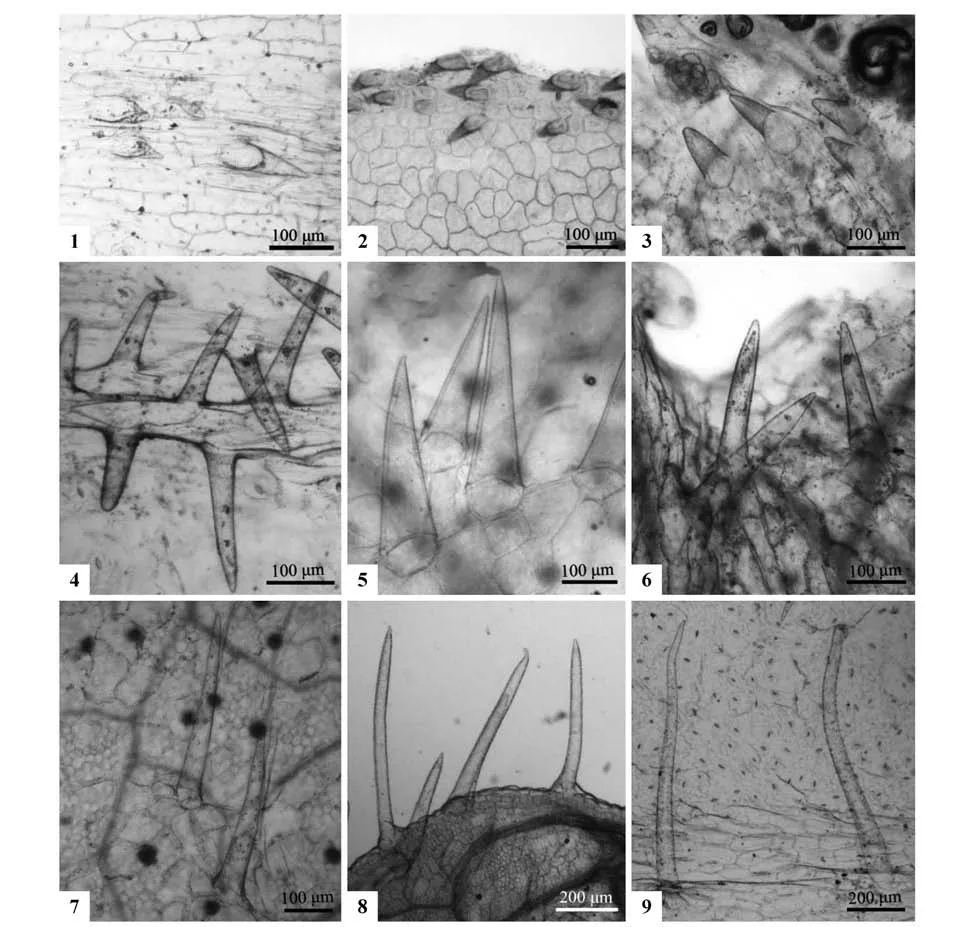
图版Ⅱ 8种堇菜属植物的3类叶表皮毛被类型1~3.短柱状毛(1,2.南山堇菜(中脉,叶缘);3.香堇菜);4~6.中柱状毛(4.紫花地丁;5.双花堇菜;6长萼堇菜);7~9.长柱状毛(7.早开堇菜;8.七星莲;9.灰叶堇菜)Plate Ⅱ Three types of leaf trichome in eight Viola species1-3.Short-column trichome(1-2.V.chaerophylloides(midrib,ridge);3.V.odorata);4-6.Medium-column trichome(4.V.philippica;5.V.biflora;6.V.inconspicua);7-9.Long-column trichome(7.V.prionantha;8.V.diffusa;9.V.delavayi)
3.2.2 叶表皮细胞和气孔特征
在显微镜下(图版Ⅲ,表2),同种植物叶表皮的上表皮细胞比下表皮细胞大,形态和垂周壁样式多为不规则形-浅波状或深波状,只有双花堇菜的为多边形-平直弓形(图版Ⅲ:1~2)。同种植物的上下表皮细胞在4 个种里相同,其余4 个种不同。相同的出现在包括双花堇菜在内的4种:长萼堇菜的为不规则浅波状(图版Ⅲ:9~10),紫花地丁和七星莲为不规则深波状(图版Ⅲ:11~12,13~14)。不同的见于另4种:早开堇菜和南山堇菜的上表皮为浅波状(图版Ⅲ:3,15),下表皮为深波状(图版Ⅲ:4);香堇菜和灰叶堇菜的上表皮为多边形和平直-弓形(图版Ⅲ:5,7),下表皮为不规则型和深波状(图版Ⅲ:6,8)。
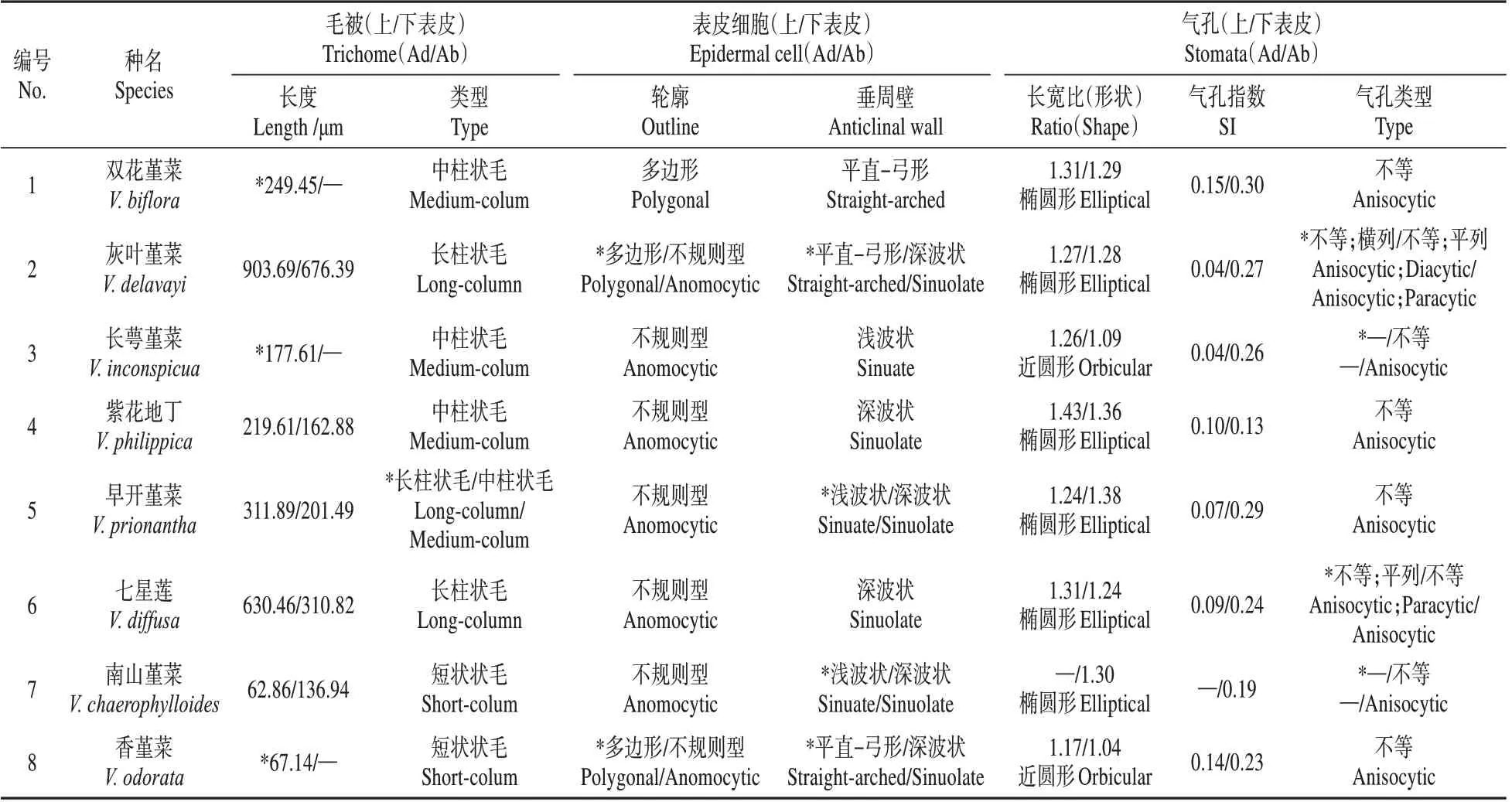
表2 8种堇菜属植物的叶表皮形态特征Table 2 Morphological characteristics of leaf epidermis in eight Viola species
8种植物均具不等型气孔,在灰叶堇菜和七星莲里还观察到少量的平列型和横列型(图版Ⅲ:7~8,13~14)。它们的气孔主要分布于叶下表皮,密度较高,为密集型气孔。除长萼堇菜和南山堇菜外,其余6 种的上表皮也观察到少量气孔,但密度较低为稀疏型。气孔有2种,除了长萼堇菜和香堇菜的气孔为近圆形(长宽比为1.04~1.20)外,其余6种都为椭圆形(长宽比为1.20~1.50)。
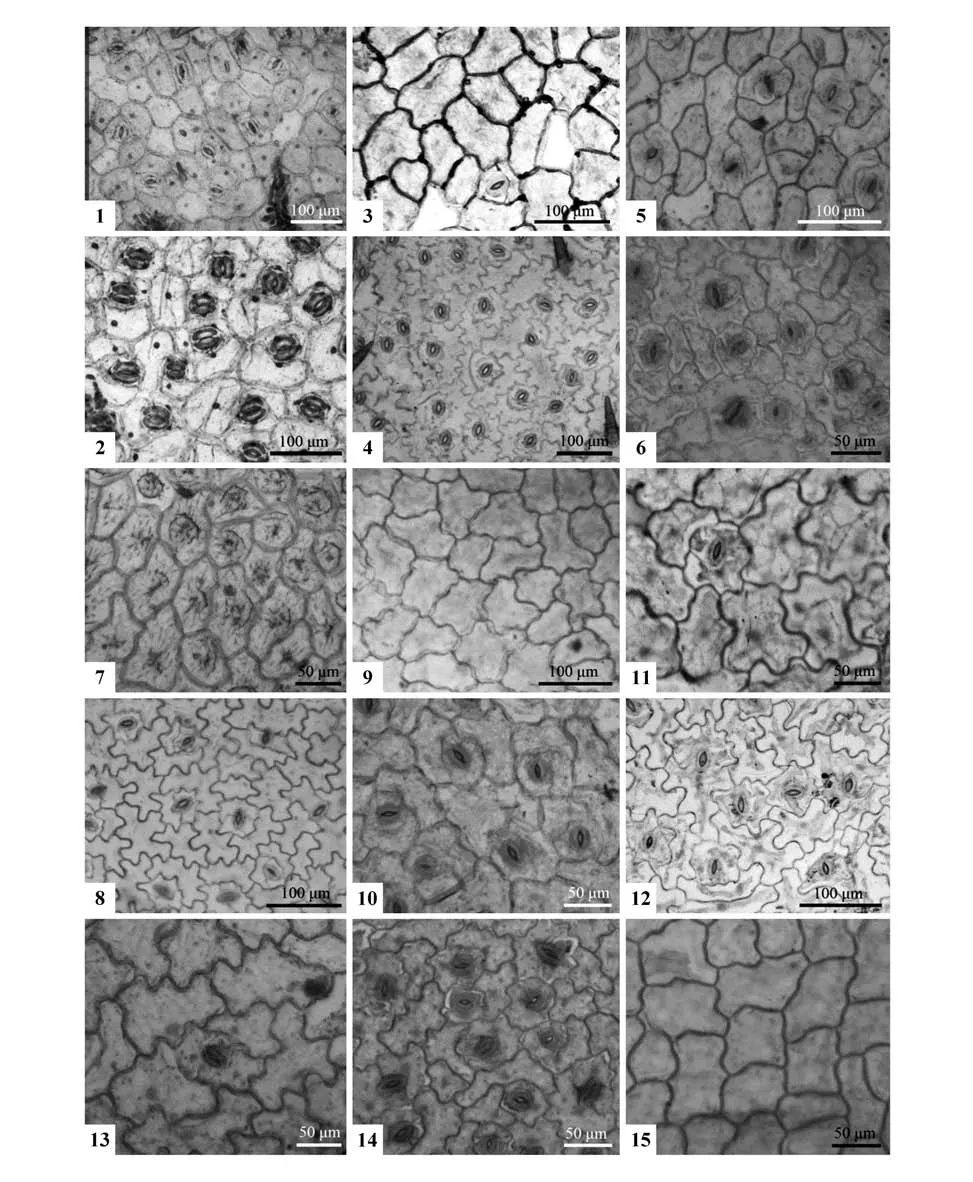
图版Ⅲ 8种堇菜属植物的叶表皮特征1~2.双花堇菜(上表皮,下表皮);3~4.早开堇菜(上表皮,下表皮);5~6.香堇菜(上表皮,下表皮);7~8.灰叶堇菜(上表皮,下表皮);9~10.长萼堇菜(上表皮,下表皮);11~12.紫花地丁(上表皮,下表皮);13~14.七星莲(上表皮,下表皮);15.南山堇菜(上表皮)Plate Ⅲ Morphological characteristics of leaf epidermis in eight Viola species1, 2.Viola biflora(Adaxial epidermis,Abaxial epidermis);3, 4.V.prionantha(Adaxial epidermis,Abaxial epidermis);5, 6.V.odorata(Adaxial epidermis,Abaxial epidermis);7, 8.V.delavayi(Adaxial epidermis,Abaxial epidermis);9, 10.V.inconspicua(Adaxial epidermis,Abaxial epidermis);11, 12.V.philippica(Adaxial epidermis,Abaxial epidermis);13, 14.V.diffusa(Adaxial epidermis,Abaxial epidermis);15.V.chaerophylloides(Adaxial epidermis)
3.3 8种堇菜属植物的叶中脉横切面解剖特征
8种植物的叶中脉横切面均由角质层、上下表皮层、栅栏组织、叶肉细胞和维管束组成(图版Ⅳ)。其中,上表皮细胞比下表皮的大,栅栏组织普遍分化不明显,部分物种存在方形晶状体,分布于栅栏和薄壁细胞之间,以双花堇菜最明显(图版Ⅳ:6),香堇菜(图版Ⅳ:1)和灰叶堇菜的零星可见(图版Ⅳ:5)。结果表明,叶中脉处的表皮突起和叶片厚度在种间差异明显(表3)。首先,叶中脉横切面主维管束处的上下表皮突起,来源于表皮下多了1层细胞形态相似的次生表皮细胞,由于种内和种间的上下表皮细胞大小不同,故形成大小相似的均等型和大小不同的不等型突起。前者指上下表皮突起微微向外隆起,大小形态一致、近似对称(图版Ⅳ:1~4),出现在4 种:香堇菜(图版Ⅳ:1)、紫花地丁(图版Ⅳ:2)、长萼堇菜(图版Ⅳ:3)和七星莲(图版Ⅳ:4)中。后者指上下表皮突起轮廓形态大小不一,见于另4种:灰叶堇菜(图版Ⅳ:5)、双花堇菜(图版Ⅳ:6)、早开堇菜(图版Ⅳ:7)和南山堇菜(图版Ⅳ:8)。再次,叶片厚度在种间也有差异,分为厚叶型(>200 µm)和薄叶型(≤200 µm),前者见于3个种:双花堇菜(267.81 µm)、紫花地丁(218.76 µm)和南山堇菜(208.76 µm);后者出现在5种:灰叶堇菜(109.84 µm)、长萼堇菜(158.35 µm)、早开堇菜(143.45 µm)、七星莲(151.66 µm)、香堇菜(195.88 µm)。其中叶片最厚的见于双花堇菜,最薄的出现在灰叶堇菜,两者厚度相差近2.5倍。
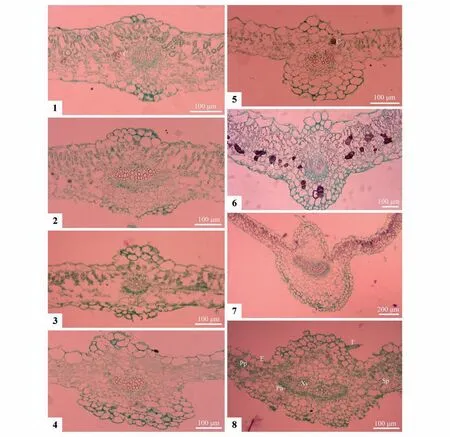
图版Ⅳ 8种堇菜属植物叶片主脉的横切面解剖特征1~4.均等型(1.香堇菜;2.紫花地丁;3.长萼堇菜;4.七星莲);5~8.不等型(5.灰叶堇菜;6.双花堇菜;7.早开堇菜;8.南山堇菜);C.晶体;E.表皮;Ph.韧皮部;Pp.栅栏组织;Sp:海绵组织;T.毛被;Xy.木质部Plate Ⅳ Cross sections of leaf midrib in eight Viola species1-4.Equa(l1.Viola odorata;2.V.philippica;3.V.inconspicua;4.V.diffusa);5-8.Unequal(5.V.delavayi;6.V.biflora;7.V.prionantha;8.V.chaerophylloides);C.Crystal;E.Epidermis;Ph.Phloem;Pp.Palisade parenchyma;Sp.Spongy parenchyma;T.Trichome;Xy.Xylem
3.4 8种堇菜属植物的叶柄横切面解剖特征
8种植物的叶柄横切面均由表皮层、厚角组织和维管束组织构成(图版Ⅴ)。表皮细胞较小、排列整齐,构成闭合结构,外壁有薄角质层。表皮内有4~10 层薄壁细胞构成的厚角组织,细胞大小不一、排列疏松,有明显胞间隙。主维管束位于中央,其形态与叶柄横切面形态相似。叶柄两侧近轴面有明显延伸生长的突起,构成带有1对侧生维管束的侧翼。比较分析结果表明,叶柄横切面轮廓、两侧近轴面的侧翼突起、主维管束形态和侧生维管束在同种植物里有一定的相关性,但在种间区别较大。叶柄横切面形态轮廓在8个种间差异明显,可分为3 类(表3):椭圆形、近圆形和圆形。椭圆形,指横切面较薄,其轮廓与主维管束的都呈椭圆状,近轴面表皮平直或中央微隆,无凹陷,远轴面为半弧形曲线;叶柄两侧外缘处带1对具次生维管束的延伸侧翼(图版Ⅴ:1~4);见于长萼堇菜、七星莲、早开堇菜和紫花地丁4 种。近圆形,指横切面较厚、近饼状,轮廓与主维管束的呈近圆形或深U 型(图版Ⅴ:6~8),近轴面两侧有1 对具侧生维管束的侧翼突起,与中央的突起构成了带有1 对侧翼的3 个脊状隆起(图版Ⅴ:6~8),见于灰叶堇菜、南山堇菜和香堇菜等3 种。圆形,指叶柄横切面厚,近轴面两侧有次生维管束不明显的侧翼突起,中央隆起呈短弧形,与远轴面的长弧形连接成带1 对侧翼圆周,与中央的圆形维管束构成同心圆或环形,只见于双花堇菜(图版Ⅴ:5)。较为特别的是,该种的主维管束形态完整,呈闭合环状,但韧皮部细胞出现分化,将其平分为4瓣,具有染色较浅的髓部。

表3 8种堇菜属植物叶中脉和叶柄的横切面解剖特征Table 3 Cross-sectional anatomical features of leaf and petiole in eight Viola species
3 讨论
本研究观察的云南常见8种堇菜属植物,分别来自2 亚属5 组[2],包括二裂花柱亚属的短距堇菜组(灰叶堇菜和双花堇菜)和堇菜亚属的4组,即合生托叶组(Sect.Adnatae)(长萼堇菜、紫花地丁、早开堇菜)、蔓茎堇菜组(Sect.Diffusae)(七星莲)、裂叶堇菜组(Sect.Pinnatae)(南山堇菜)、堇菜组(Sect.Viola)(香堇菜)。围绕着堇菜属存在的组间分类和物种鉴定困难,本研究讨论了它们的叶形态和解剖特征的分类学意义。
3.1 堇菜属区域内种间的叶型变化,可用于当地物种鉴定参考依据
长期以来,堇菜属的组和亚组的划分依据主要是花形态特征变异,尤其是柱头和花柱特征是主要的分类学依据[8]。在野外,堇菜属的花柱是否弯曲、柱头孔大小和位置等有分类学意义的显微特征不容易观察,导致了物种鉴定相对困难[3-4]。为此,本研究讨论了云南8种常见植物的叶形态分类学意义。首先,双花堇菜和灰叶堇菜均见于海拔2 000 m 以上的高山草地,花期相同,均为黄花,属于二裂柱头亚属的短距组,野外不易区别,两者仅在叶形态区别明显。双花堇菜的叶型多为宽卵形或肾形,其基部为深心形或心形;灰叶堇菜的叶片为三角状宽卵形,其叶基为浅心形或截型。其次,堇菜亚属的合生托叶组的3种(长萼堇菜、紫花地丁和早开堇菜),由于同具紫花、叶型相似,常被列为近似种,甚至合并为紫花地丁一个种[2,24]。但在野外,长萼堇菜为宽卵形叶,叶缘具粗齿、叶基心形、叶尖明显;早开堇菜宽卵形叶,近椭圆形,叶缘具细齿、叶基弧心形,叶尖不显著;紫花地丁叶片为狭卵型,仅长披针形,足以和前2 种区别。因此,这3 个种不宜合并,支持前人的观点[2]。此外,深裂叶片和肾形叶在云南常见的堇菜属里较为少见,在当地可以作为物种鉴定依据。深裂叶片是南山堇菜所在的裂叶堇菜组的主要分类学依据,稀不裂[2-3]。该组在我国有8 种,大部分都分布在东北,云南只有1 种,为羽裂堇菜(V.forrestiana),分布在滇西北和藏东南高山草地,但该种的叶片为三角状卵形或狭卵型,只在叶缘处有羽状浅裂[26]。同时,在国产的裂叶堇菜组的8 个种里,南山堇菜的三深裂叶片且开白花的形态特征也较为独特[2-3]。因此,对于云南常见堇菜属植物的物种鉴定而言,看到三深裂的羽状裂片叶,就可以将其初步判断为南山堇菜。肾形叶见于堇菜组的香堇菜,虽分布在欧洲、北非和西亚等地,但由于具有较高观赏价值,被广为栽培,其肾形叶或近圆形叶在园艺堇菜植物里也较为独特且少见。因此,肾形叶是香堇菜的物种鉴别特征之一。当然,叶型在物种多样性较高的堇菜属里变异较大,部分种类由于生境不同而产生了不同的变型或亚种,值得进一步研究。但在部分地域范围内、具明显地理分布格局的堇菜属区系范围,种间的叶型变化可作为云南常见种或近似种的鉴定依据之一[26]。
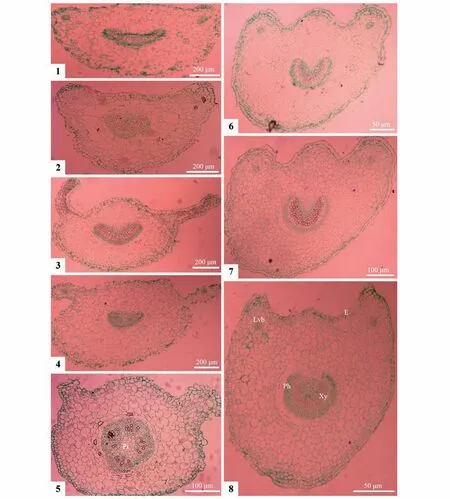
图版V 8种堇菜属植物叶柄横切面解剖特征1~4.椭圆形(1.长萼堇菜;2.紫花地丁;3.早开堇菜;4.七星莲);5.近圆形(双花堇菜);6~8.深U 形(6.灰叶堇菜;7.南山堇菜;8.香堇菜);E.表皮;Lvb.侧生维管束;Pi.髓部;Ph.韧皮部;Xy.木质部Plate V Cross section of leaf petiole in eight Viola species1-4.Elliptica(l1.V.inconspicua;2.V.philippica;3.V.prionantha;4.V.diffusa);5.Orbicula(rV.biflora);6-8.Deep-U shape(6.V.delavayi;7.V.chaerophylloides;8.V.odorata);E.Epidermis;Lvb.Lateral vascular bundle;Ph.Phloem;Pi.Pith;Xy.Xylem
3.2 堇菜属毛被特征和长度在种间的变化,可作为物种划分依据
关于堇菜属毛被特征的分类学意义,研究资料较少。早期研究表明,堇菜属的叶毛被类型为简单的单细胞毛或单列毛,并指出在叶缘有锯齿的种类里,其叶缘处的毛被为粗糙的腺体毛[29]。后来的研究也陆续证实了堇菜属的叶表皮毛普遍为单细胞毛,上下表皮毛被的有无及其分布特征都用于物种鉴定。例如,南京产的紫花堇菜(V.grypoceras)和球果堇菜(V.collina)的下表皮都密被单细胞毛[30];东北产的27种堇菜属植物里,除香堇菜为2 细胞单列毛外,其余26 种都为单细胞毛[25];伊朗产的3 个近缘种(香堇菜、白花堇菜V.alba&V.sintenisii)也都具有单细胞毛[16]。本研究观察的8种植物的叶表皮毛被类型都为单细胞毛,没有观察到单列毛,也未在叶缘有锯齿的种类里看到叶缘处的有腺体毛。值得注意的是,伊朗产的香堇菜单细胞毛只分布在上表皮,被列为区分近似种的鉴定依据[16]。该种的毛被特征与本研究观察的云南产的香堇菜一致。如上所述,单列毛在堇菜属里较为少见,只在堇菜组的球果堇菜观察到2细胞单列毛[24],目前尚未有其他类型的单列毛的资料。有意思的是,球果堇菜产自东北的植株有2 细胞单列毛,但长在南京的个体,其毛被为单细胞长柱状毛,且较为密集[30]。此外,香堇菜也属于堇菜组,但无论是产自伊朗或生长在云南的植株,其毛被都为单细胞毛。显然,毛被类型在堇菜属的分类学意义,需要更多新资料佐证。
如上所述,堇菜属的毛被类型和分布在种间差异明显,可作为物种划定依据。然而,毛被长度在种间的变化和分类学意义,鲜有人关注。辽东地区的15种堇菜属植物的毛被长度在种间差异明显[31],但缺乏深入研究的数据。本研究观察到毛被长度在8种植物里区别明显,将其分为3 类,即:短柱状毛、中柱状毛和长柔毛。其中,最容易鉴别的是短柱状毛,其长度仅为60 µm 左右,基部宽、顶部锐,状如宽短刺或盾鳞片,长度不及中柱状毛和长柱状毛的十分之一,而后者为基部宽而顶部尖的长单细胞毛。短柱状毛较少,仅见于具肾形叶的香堇菜(堇菜组)和具羽状深裂叶的南山堇菜(裂叶堇菜组)。其次,中柱状毛集中出现在堇菜亚属的合生托叶组的3 个种,而蔓茎堇菜组(七星莲)上表皮为长柱状毛,下表皮为中柱状毛。这意味着长柱状毛也许可以作为合生托叶组的分类学依据之一。同样地,二裂柱头堇菜亚属的灰叶堇菜和双花堇菜各具长柱状毛和中柱状毛,两类毛被的长度相差近4 倍,较容易区别。因此,毛被长度在种间的差异,可以作为物种鉴定依据。
3.3 堇菜属的叶表皮细胞类型和气孔形态可区别近似种
首先,堇菜属的叶表皮细胞形态和垂周壁样式在种间差异明显,多数为不规则型和浅波状或深波状,少数为多边形和平直-弓形[24,29]。本研究观察的8种植物,表皮细胞不规则型带波状垂周壁的出现在堇菜亚属的3 组5 种(南山堇菜除外),多边形和平直-弓形表皮细胞见于3 种,即二裂柱头亚属的双花堇菜、灰叶堇菜和南山堇菜。目前,关于叶表皮细胞类型的分类学意义,仅见于近似种间的划分[25]。本研究观察的近似种,如双花堇菜和灰叶堇菜虽然在叶型上区别明显,但叶表皮细胞形态差异也可将其分开,即:虽然它们的上表皮都为多边形,但后者的下表皮为不规则深波状,可以将2种分开。在长萼堇菜、紫花地丁和早开堇菜这3个近似种的鉴定里,前者上下表皮为浅波状与后2 种的深波状区别明显。紫花地丁的上下表皮皆为深波状,但早开堇菜的上表皮为浅波,下表皮为深波,也可作为鉴定参考依据。
其次,堇菜属的气孔普遍见于上下表皮,下表皮比上表皮常见,气孔呈椭圆形,或近圆形,气孔器类型普遍为不等型或横列型等[15,29,30]。本研究观察的8 种植物,除了南山堇菜的上表皮无气孔外,其余上下表皮都有不等型气孔,在灰叶堇菜和七星莲还兼具平列型、横列型或不等型。除长萼堇菜和香堇菜的气孔为圆形外,其余6种都为椭圆形。显然气孔形态在种内比较稳定,在种间差异明显,可用于物种划分。例如,根据气孔形态,可以把长萼堇菜与其近似种(紫花地丁和早开堇菜)区别开来,前者具圆形气孔,后两种为椭圆形气孔。然而,气孔器类型在同种和不同种植物里变化较大,气孔密度和气孔指数虽然种间也有差异,但难以比较其在种间或组间的分布规律,更多的与物种适应环境相关[28]。不过,也有研究认为,气孔类型可以作为堇菜属的组间界限划分依据。例如,以三色堇为代表的美丽堇菜亚属的气孔类型为围绕型,以双花堇菜为代表的二裂柱头亚属为极细胞型;裂叶堇菜组不具有四细胞型气孔器,因此支持其独立为组[23]。显然,仅以1个物种为代表来探究气孔类型在组间的分类学意义有其局限性,仍需更多资料来支持。
3.4 堇菜属叶中脉横切和叶柄横切面结构特征的种间异同可用于讨论近似种的关系
叶中脉和叶柄横切面轮廓、主维管束形态、次生维管束数目和晶状体的有无等特征在同种植物里是相对稳定而恒定的,具有重要的物种鉴定意义[32-34]。但关于堇菜属的叶中脉和叶柄横切面解剖特征的分类学研究的资料较少,本研究初步分析如下。
首先,关于叶中脉处结构特征的分类学意义有两点值得说明。第一,早期研究记录了堇菜属的部分种类叶中脉横切面栅栏组织化不明显,但在夏威夷分布的种类里分化为形态显著的栅栏和海绵组织[19,30]。该栅栏组织分化不明显的叶解剖特征,在伊朗产的堇菜属植物里也普遍[15],本研究观察的云南产的8个种的栅栏组织也不明显,说明该特征值得继续关注。第二,堇菜属的叶中脉横切面的上下表皮突起在种间多为凸起型,少数为平直型,可用于伊朗产的堇菜属近似种和亚种间的区别[16]。然而,他们观察的8 种植物都为凸起型,没观察到平直型,且叶中脉处的上下表皮突起在同种植物也有差异,可分为均等型和不等型,在种间差异明显,但对此没有深入分析[16]。本研究观察到的同种植物上下表皮突起是否等大的特征,可以较好地用于物种鉴定。例如:均等型突起出现在堇菜亚属的3 个近似种(香堇菜、紫花地丁和长萼堇菜)和七星莲。这证实了3个近似种在叶形态解剖特征方面具有相似性,但3种的主维管束的形态又各有不同,可以相区别,支持其为独立的近似种,并与七星莲还有一定的相关性。反之,不等型上下表皮突起见于其余4 种。虽然它们的主维管束都为典型的新月形,但各有区别。例如,除南山堇菜外,其余3 种的下表皮突起大于上表皮;双花堇菜和灰叶堇菜的维管束形态为近圆形,前者有大量的方形晶状体,后者有稀疏星状晶体;早开堇菜单细胞毛。
其次,关于堇菜属叶柄解剖特征的分类学意义,集中在叶柄横切面及主维管束的形态、次生维管束数目、上表皮两侧的翼状突起及中部的脊状突起数目等在种间差别明显[15-16,33]。与上述研究结果基本相似,本研究观察的8种植物的叶柄横切面均具有中央维管束和1 对带次级维管束的两侧翼状突起。但叶柄横切面和中央维管束形态相关,基本为椭圆形、近圆形和圆形等,在种间差异较大,可作为近似种之间的分析[16]。例如,双花堇菜和灰叶堇菜这对近似种,两者区别明显,前者叶柄横切面为圆形,后者呈具有缺口的近圆形。值得关注的是,双花堇菜的叶柄解剖特征很特别,主维管束形态为闭合状圆形,其中央分化出髓,这在现有资料里都是极少见的,值得进一步研究。不过,堇菜亚属的3个近似种(早开堇菜、长萼堇菜和紫花地丁)和七星莲,具有相似的叶柄横切面形态,均为椭圆形和具月牙形主维管束特征。因此,结合它们在叶中脉横切面解剖特征的相似性,进一步说明了这3个近似种为有近缘关系的独立种,同时还与七星莲有一定的亲缘关系,值得进一步研究。
再次,关于堇菜属的叶片晶状体的形态及其分类学意义资料较少。早期研究记录了堇菜属的叶肉和叶柄含有大量的单独或成簇的晶状体[16,21,25];在伊朗产的物种里,星状或方形的晶状体也普遍见于不同种,可作为属下分组的依据[15-17]。但本研究在8种植物的叶中脉和叶柄横切面结构上,仅在双花堇菜发现有密集的方形晶状体,在灰叶堇菜和香堇菜的叶中脉和叶柄横切面上有稀疏分布的晶体。
综上所述,堇菜属的叶型态、表皮毛被类型、叶表皮细胞形态、气孔形态和气孔器类型、叶中脉和叶柄横切面解剖特征在种间差异明显,可用于属内物种鉴定及近似种的划分[28-29]。根据叶形态、叶中脉和叶柄横切面解剖特征,编制了云南常见8种堇菜属植物的分类检索表,具体如下。
8种堇菜属植物检索表-主要根据叶形态解剖特征
Key to eight Viola species mainly based on the morpho-anatomy of leaf
1.花黄色。………………………………………………………………………………………………………………………………………(2)
1.花白色或紫色。………………………………………………………………………………………………………………………………(3)
2.卵形叶;叶下表皮细胞为不规则型,垂周壁深波状;长柱状毛;中脉处上下叶表皮突起为均等型,叶片薄;叶柄横切面近圆形,主维管束深U型……………………………………………………………………………………………………………………………灰叶堇菜V.delavayi
2.肾形叶;叶下表皮细胞多边形,垂周壁平直弓形;中柱状毛;中脉处上下叶表皮突起为不等型,叶片厚;叶柄横切面圆形,主维管束圆型……………………………………………………………………………………………………………………………………双花堇菜V.biflora
3.花白色;叶片三全裂………………………………………………………………………………………………南山堇菜V.chaerophylloides
3.花紫色,叶片非三全裂。………………………………………………………………………………………………………………………(4)
4.戟形叶……………………………………………………………………………………………………………………长萼堇菜V.inconspicua
4.卵形叶。………………………………………………………………………………………………………………………………………(5)
5.叶上下表皮细胞形态相同;叶中脉处上下表皮突起均等型。………………………………………………………………………………(6)
5.叶上下表皮细胞形态不同;叶中脉处上下表皮突起不等型。………………………………………………………………………………(7)
6.叶片薄;短柱状毛;叶柄横切面半圆形,主维管束深U形……………………………………………………………………香堇菜V.odorata
6.叶片厚;中柱状毛;叶柄横切面椭圆形,主维管束浅U形………………………………………………………………紫花地丁V.philippica
7.叶上表皮细胞垂周壁式样不规则浅波,下表皮为不规则深波状………………………………………………………早开堇菜V.prionantha
7.叶上下表皮细胞均为不规则深波状……………………………………………………………………………………………七星莲V.diffusa
