棉花脱水素GhDHN1的克隆及其表达
2016-09-16王俊娟陆许可陈修贵王德龙樊伟丽阴祖军郭丽雪叶武威喻树迅
王俊娟,穆 敏,王 帅,陆许可,陈修贵,王德龙,樊伟丽,阴祖军,郭丽雪,叶武威,喻树迅
(中国农业科学院棉花研究所/棉花生物学国家重点实验室/农业部棉花遗传改良重点开放实验室,河南安阳 455000)
棉花脱水素GhDHN1的克隆及其表达
王俊娟,穆 敏,王 帅,陆许可,陈修贵,王德龙,樊伟丽,阴祖军,郭丽雪,叶武威,喻树迅
(中国农业科学院棉花研究所/棉花生物学国家重点实验室/农业部棉花遗传改良重点开放实验室,河南安阳 455000)
【目的】通过对棉花脱水素基因结构特征及其在低温胁迫下表达模式进行分析,探讨脱水素在棉花响应低温过程中的功能,为棉花抗冷育种提供理论基础。【方法】以棉花抗冷品种豫2067为试验材料,根据棉花陆地棉基因组序列查找已知脱水素(dehydrin,Dhn)基因的CDS序列,利用Primer5软件设计引物,克隆该基因,并命名为GhDHN1;采用生物信息学方法分析其蛋白质性质、氨基酸含量特征、功能结构域、系统进化树;选择XbaⅠ和SmaⅠ酶切位点对植物表达载体pBI121∶∶GFP进行双酶切,采用In-Fusion连接技术构建融合蛋白瞬时表达载体pBI121-GhDHN1∶∶GFP;分析其在洋葱表皮细胞中的瞬时表达,进行亚细胞定位;利用抗冷材料豫2067在三叶期对低温处理(4℃,24 h)前后的叶片和根系进行转录组测序,筛选差异表达基因;在三叶期时分别对豫2067(抗冷品种)和衡棉3号(冷敏感品种)进行低温(4℃,24 h)处理,利用实时荧光定量方法分别比较根、茎、叶中GhDHN1表达量,对该基因在2个抗冷差异材料叶中的表达量进行比较;对豫2067进行不同时间低温(4℃)处理,分析GhDHN1在叶片和根中的动态表达模式。【结果】该基因全长为726 bp,开放阅读编码框为636 bp,编码211个氨基酸,预测分子量为23.79 kD,等电点为5.04,富含谷氨酸(26.10%)和赖氨酸(19.40%),不含色氨酸,半衰期为 30 h,蛋白呈酸性,带负带荷,带负电荷的残基总数为60%;GhDHN1的二级结构α螺旋(Alpha helix)包含116个氨基酸残基,占54.98%,组成该蛋白的主体结构,无规则卷曲(Random coil)的氨基酸残基有87个;GhDHN1位于陆地棉D亚组第9染色体(Dt_chr9)上,在cDNA的259—348位置上含有一个长度为90 bp的内含子,2个外显子长度分别为258 和378 bp;SMART和CDD分析表明该氨基酸序列含有2个保守的富含赖氨酸的K片段和1个保守的富含丝氨酸S片段,具有亲水素蛋白结构域pfam00257,表明该蛋白为K2S型脱水素;系统进化树分析表明,陆地棉亲水素GhDHN1与可可亲缘关系最近;洋葱表皮细胞中瞬时表达分析表明,GhDHN1蛋白主要定位在细胞膜附近。转录组分析表明,该基因在棉花三叶期叶片和根中,受低温处理后上调表达;荧光定量PCR分析表明,GhDHN1 在4℃低温胁迫24 h后,在叶片、茎、根中均上调表达,在叶中上调表达倍数最大,在低温处理4 h和24 h时叶片中有2个表达高峰,在低温处理6 h和12 h时在根中有2个表达高峰,在抗冷材料叶片中的表达是冷敏感材料的 2.47倍,说明该基因可能参与了棉花对低温的适应性调控。【结论】陆地棉GhDHN1属于典型的K2S型脱水素,与可可亲缘关系最近。该基因响应低温胁迫,在抗冷材料和冷敏感材料中表达差异显著,其表达量与棉花的抗冷性呈正相关,可以作为筛选不同抗冷材料的标记,同时可以作为重要的候选基因来培育棉花抗冷新材料。
棉花;脱水素;亚细胞定位;低温胁迫;转录组分析;实时荧光定量PCR
0 引言
【研究意义】低温胁迫是严重影响植物生长和存活的主要逆境之一[1]。棉花的整个生育期均对低温比较敏感,特别是苗期更容易遭受低温冷害,低温对棉花生长的影响远大于其他作物[2]。因此,挖掘棉花抗冷相关功能基因,对开展棉花抗冷分子育种具有重要意义。【前人研究进展】植物具有很高的能力适应外界环境条件的变化,在面临低温胁迫时会产生一系列的生理生化和基因表达变化,植物响应低温胁迫的最初反应是细胞质中钙离子的释放[3];其次是伴随着大量基因的表达,细胞内会诱导合成一系列的功能蛋白,以保护其免受胁迫的伤害[4],植物应对细胞脱水的一条主要途径是积累亲水性蛋白。LEA(late embryogenesis abundant proteins)蛋白,即胚胎发育后期丰富蛋白[5],是一类重要的植物细胞脱水保护蛋白。大量研究表明,LEA蛋白在植物响应各种逆境胁迫中起着重要作用,其首先在棉花子叶中被发现[6],根据其氨基酸序列及其保守域特征,将 LEA蛋白分为 6类(Ⅰ—Ⅵ),其中,脱水素(dehydrin,Dhn)属于第Ⅱ类,又名LEAⅡ蛋白,研究表明脱水素具有很强的热稳定性,在非生物逆境胁迫条件下对维持植物细胞的正常代谢和细胞膜结构稳定性具有重要的保护功能[7]。脱水素最基本的功能是清除细胞内自由基,保护细胞能正常行使功能[8],甚至对植株整个蛋白质组进行保护[9]。近年来,研究人员对植物响应低温胁迫过程中脱水素的功能开展了一系列的研究,KOSOVÁ等[10]研究表明经过低温胁迫后,10个大麦品种中Dhn5的积累量与其抗冻能力成正相关;郭鹏等[11]利用 Northern杂交分析表明,番茄脱水素基因SlDHN2b受冷胁迫处理后在番茄根、茎、花蕾、果实、叶中的表达量均高于其常温(25℃)条件下在相应组织的表达量。在小麦的脱水素研究中发现,小麦脱水素基因TaDHN-1通过依赖ABA非生物胁迫响应路径发挥功能,参与了小麦对低温等逆境胁迫的调节过程[12];PUHAKAINEN等[13]在拟南芥中同时过量表达几个拟南芥脱水素基因后,在转基因植株中积累了大量的脱水素,与野生型对照相比,转基因植株在面对冻害胁迫时表现出较低的半致死温度和较高的存活率,同时证明了脱水素基因通过保护细胞膜来提高植物的抗冷性。高粱脱水素基因 SbDhn2在高粱遇到低温胁迫时能够保护细胞内乳酸脱氢酶行使正常功能,增强其他蛋白酶的抗逆性[14]。在啤酒酵母(Saccharomyces cerevisiae)中过表达马铃薯Y2K型脱水素 LEA4后,酵母的抗冻性大大增强[15]。SWIRE-CLARK等[16]在酵母中异源表达LEA/DHN后,发现 LEA/DHN的主要功能是通过增强抗渗透胁迫能力来提高冷胁迫的能力。【本研究切入点】在笔者前期研究中,系统分析了棉花叶片低温转录组差异基因,获得了一批与抗冷性密切相关的候选基因,在这些候选基因中,有一个被注释为脱水素基因(CotAD_58358),关于该基因在棉花中对低温等非生物胁迫的响应及调控机制鲜见报道。【拟解决的关键问题】本研究分析棉花脱水素基因及其编码蛋白的结构特性,构建植物表达载体,研究其亚细胞定位,在转录组测序水平和实时荧光表达水平检测该基因受低温胁迫过程中在根、茎叶中的表达模式,为在棉花抗冷分子育种中进一步应用奠定基础。
1 材料与方法
1.1试验材料及其培养和低温处理
所用材料为陆地棉品种豫2067和衡棉3号,由中国农业科学院棉花研究所提供。于2015年9月在中国农业科学院棉花研究所抗逆鉴定实验室恒温光照生长箱中进行育苗,采用沙培法育苗(28℃,14 h光照/10 h黑暗),正常水分条件(沙土相对含水量约23%)下进行棉花的培养,在三叶期进行4℃处理0 h(CK,即28℃)和24 h;对抗冷材料豫2067分别取根、茎、叶,其中根取样部位是须根,茎取样部位是下胚轴,叶片取样部位是真叶的倒一叶,对冷敏感品种衡棉 3号只取叶片,取样部位为倒一叶,每部位均取若干重复,剪碎,迅速放入液氮中速冻,然后放入-80℃超低温冰箱中保存备用。衡棉3号的叶片用于实时荧光定量分析,豫2067的一部分根、茎、叶样品用于实时荧光定量分析,在棉花生物学重点实验室进行;另一部分的叶片和根用于转录组测序,由深圳华大基因公司完成。
同时对豫2067三叶期进行低温不同时间(4℃,0、0.5、2、4、6、8、12和24 h)处理,其中0 h为对照,取样部位为倒一叶和根系,用于实时荧光定量分析。
1.2RNA的提取和全长GhDHN1的CDS序列克隆
利用北京艾德莱生物科技有限公司生产的EASYspinPlus植物RNA快速提取试剂盒对以上所取样品进行RNA提取,利用Nanodrop2000核酸分析仪测定总RNA的浓度和纯度,A260/280=2.0,同时进行1% (m/v)琼脂糖凝胶电泳检测RNA的完整性。在棉花基因组 Cotton Genome Project (CGP,http://cgp. genomics.org.cn/page/species/index.jsp)中搜索到的脱水素(dehydrin,Dhn)基因(CotAD_58358)的CDS序列,利用Primer5软件设计特异引物,上游引物为GhDHN1-F:5′-ATGGCCGAGGAGCATACCAGTA-3′,下游引物为GhDHN1-R:5′-TCAAGCCTTTTCTTTT TCTTCA-3′。利用 PrimeScript RT Reagent Kit with gDNA Eraser(Perfect Real Time)试剂盒(TaKaRa,China)将豫2067叶片RNA反转录合成第一链cDNA,以反转录获得的棉花 cDNA为模板,扩增 GhDHN1全长cDNA序列。所用的PCR程序为94℃ 5 min;94℃40 s,54℃ 40 s,72℃ 40 s,30 cycle;72℃ 10 min。扩增产物采用1%琼脂糖凝胶电泳(m/v)进行检测。PCR产物采用Omega PCR产物纯化试剂盒进行纯化。然后将目的基因与PMD-19载体连接,转化至大肠杆菌感受态细胞 DH5α,倒置培养过夜,挑取白色单克隆,进行PCR检测,并且测序进行验证。测序由苏州金唯智科技有限公司北京分公司完成。
1.3棉花GhDHN1的生物信息学分析
1.3.1棉花GhDHN1蛋白质的等电点、分子量及结构域的预测 用Protparam(http://web.expasy.org/ protparam/)在线程序分析蛋白质的分子量和等电点,ProtScale(http://web.expasy.org/protscale/)预测亲疏水性,利用 ScanProsit、SMART(http://smart.emblheidelberg.de/)、CCD(http://www.ncbi.nlm.nih.gov/ Structure/cdd/wrpsb.cgi)、PROSITE(http://prosite. expasy.org/)网站对基因进行蛋白质结构域预测,确定该基因所属蛋白家族,SOPMA(https://npsa-prabi. ibcp.fr/cgi-bin/secpred_sopma.pl)分析蛋白的二级结构,NetPhos 2.0 Server(http://www.cbs.dtu.dk/services/ NetPhos/)程序预测磷酸化位点。
1.3.2棉花GhDHN1与其他几个物种DHN1进化树分析
在NCBI上查找葡萄(Vitis vinifera,NP_001268221.1)、可可(Theobroma cacao,XP_007017965.1)、拟南芥(Arabidopsis thaliana,CAA62449.1)、柑橘(Citrus sinensis,NP_001275806.1)、高粱(Sorghum bicolor,AGS16688.1)、苹果(Malus domestica,AFG33211.1)、玉米(Zea mays,NP_001105419.1)、山杨(Populus davidiana,ABS12334.1)、白骨壤(Avicennia marina,A8CVF3.1)、油茶(Camellia oleifera,ACF72673)、小麦(Triticum aestivum,AF453444_1)、葡萄柚(Citrus ×paradisi,AAN78125.1)等12个物种DHN1的蛋白序列,分别命名为VvDHN1、TcDHN1、AtDHN1、CsDHN1、SbDHN1、MdDHN1、ZmDHN1、PdDHN1、AmDHN1,CoDHN2、WZY1-1、CpDHN,利用在线工具ClustalW2和MEGA5.0对不同物种的DHN1全长蛋白质序列进行多序列比对,使用邻接(Neighbor-Joining,NJ)法建树,用MEGA软件生成系统进化树。
1.3.3棉花GhDHN1染色体定位以及基因结构分析
分别在棉花基因组中获得GhDHN1(CotAD_58358)的CDS、cDNA全长序列和基因组序列,将cDNA序列作为查询序列与整个陆地棉AD基因组进行比对,获得GhDHN1在染色体中的位置;利用在线软件Gene Structure Display Server(GSDS2.0,http:// gsds.cbi.pku.edu.cn/)将GhDHN1的cDNA与其对应的基因组DNA序列进行比对,确定GhDHN1的外显子/内含子结构。
1.4pBI121-GhDHN1::GFP荧光瞬时表达载体的构建及洋葱表皮细胞中的亚细胞定位分析
利用http://bioinfo.clontech.com/infusion在线设计In-Fusion引物(上游引物为InGhDHN1-F:5′-CACGG GGGACTCTAGAATGGCCGAGGAGCATACCAGTA -3′,下游引物为InGhDHN1-R:5′-AGGGACTGACCA CCCGGGTCAAGCCTTTTCTTTTTCTTCA-3′,下划线为酶切位点),以GhDHN1质粒为模板进行扩增。选择的酶切位点为 XbaⅠ和 SmaⅠ,对植物表达载体pBI121:GFP进行双酶切,采用In-Fusion连接技术构建融合蛋白瞬时表达载体 pBI121-GhDHN1:GFP,转化至大肠杆菌DH5α感受态细胞,挑选阳性克隆进行测序,同时选用限制性内切酶BglⅡ和XhoⅠ验证插入位点后提取质粒。
将洋葱切成1—2 cm的小块,用基因枪GDS-80(购自美国 Wealtec Corp)活体转化技术将pBI121:GFP质粒和pBI121-GhDHN1:GFP质粒分别轰入洋葱内表皮细胞,把转化后的洋葱贴在MS培养基上,暗处室温过夜培养。用镊子轻轻撕下转化后的洋葱内表皮放在载玻片上,在激光共聚焦显微镜FV1000(日本Olympus)下观察。
1.5基于转录组测序的GhDHN1表达分析
GhDHN1受低温胁迫后的转录表达分析在陆地棉豫2067三叶期的真叶和根中进行,低温处理为4℃,24 h,对照为4℃处理0 h(28℃),取样部位是倒一叶和根系。为了评价基因表达水平,将获得的GhDHN1序列上的序列读数转换为RPKM[17],公式如下:
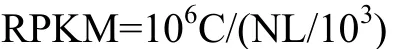
式中,C指的是唯一比对到转录组的读长的数量,N是指唯一比对到一个特定样品上的读长的总数,L是指转录本中的碱基数。
1.6三叶期棉花幼苗低温胁迫后GhDHN1的实时荧光定量分析
利用PrimerPremier5.0设计GhDHN1的荧光定量引物(上游引物为GhDHN1-F:5′- GTTAGCGGTGAA GGAGCAGT-3′,下游引物为GhDHN1-R:5′- ACTCGG TTACGATCACCTCC-3′),以 Gossypium hirsutum Histone-3(Accession No.:AF024716)作为反应中的内参基因进行实时荧光定量 PCR,PCR程序设定为94℃ 30 s;94℃ 5 s,55℃ 34 s,72℃ 34 s,共40个循环,所用仪器为7500 Real Time PCR System。实时荧光定量PCR结果分析参考AFRIN等[18]所报道2-ΔΔCt方法,每处理3个生物学重复,3个技术重复,结果用于平均数统计和方差分析。
2 结果
2.1GhDHN1的克隆
在前期的转录组试验中,发现一个注释为脱水素的基因受 4℃低温胁迫后在棉花叶片和根中均上调表达,推测其与棉花的抗冷性有一定的相关性。根据棉花基因组Cotton Genome Project(CGP,http://cgp.genomics. org.cn/page/species/index.jsp)序列中搜索到的脱水素(dehydrin,Dhn)基因的CDS全长序列,在其两端设计引物,以棉花豫2067叶片的cDNA为模板,扩增出GhDHN1的完整CDS序列(图1)。得到的阅读框长度为636 bp,测序结果正确。
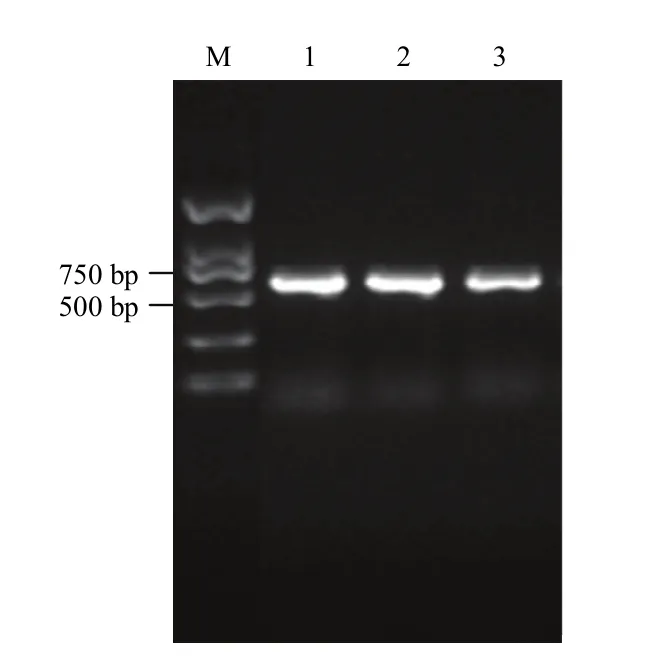
图1 GhDHN1的扩增PCR产物Fig. 1 The product of PCR amplification of GhDHN1 gene
2.2棉花GhDHN1蛋白质的等电点、分子量及二级结构域的预测
GhDHN1蛋白质编码211个氨基酸,预测分子量为23.79 kD,等电点为5.04,富含谷氨酸(26.10%)和赖氨酸(19.40%),不含色氨酸,符合植物脱水素的一般特性(图 2-A)[19]。带负电荷的酸性氨基酸(Asp+Glu)有 61个,带正电荷的碱性氨基酸(Arg+Lys)有43个,该蛋白呈酸性且带负带荷,半衰期约为30 h,不稳定指数为70.27。分析GhDHN1蛋白的亲疏水性发现,蛋白中亲水性氨基酸明显多于疏水性氨基酸(图2-B),亲水性平均系数(GRAVY, Grand average of hydropathicity)为-1.53,属亲水性蛋白。SOPMA分析结果(图2-C)表明,α螺旋(alpha helix)包含116个氨基酸残基,占54.98%,组成该蛋白的主体结构;无规则卷曲(random coil)的氨基酸残基有87个,占41.23%;β片层(β-sheet)包含氨基酸残基5个,占2.37%;β-转角(beta turn)包含3个氨基酸,占 1.42%,推测该蛋白的结构功能域可能主要由α螺旋构成。
2.3棉花GhDHN1的染色体定位、基因结构分析和功能结构域分析
在棉花基因组 Cotton Genome Project (CGP,http://cgp.genomics.org.cn/page/species/index.jsp)中对陆地棉AD基因组信息分析,结果表明,GhDHN1位于陆地棉D亚组第9染色体(Dt_chr9)上的54183731 —54184456(+),全长726 bp;基因结构分析表明,GhDHN1在cDNA的259—348位置上含有一个长度为90 bp的内含子,2个外显子长度分别为258 bp和378 bp(图3)。
利用SMART(http://smart.embl-heidelberg.de/)、CCD(http://www.ncbi.nlm.nih.gov/Structure/cdd/wrpsb. cgi)、PROSITE(http://prosite.expasy.org/)对棉花GhDHN1进行功能结构域分析可知,该氨基酸序列在C端含有 2个保守的富含赖氨酸片段 PS00823 (KIKEKLPG,位于151—158;KIKEKLPG,位于192 —199),在N端一个保守的富含丝氨酸片段PS00315 (SSSSSDEEEGEGEEKKKKKK,位于84—103),具有亲水素蛋白结构域pfam00257(位于14—160和143—201),属于K2S型脱水素蛋白的典型结构。
2.4棉花GhDHN1编码蛋白的磷酸化位点预测
利用在线软件NetPhos 2.0 Server预测GhDHN1编码蛋白的磷酸化位点(图4)。结果表明,GhDHN1蛋白主要含有16个丝氨酸(Serine)磷酸化位点(分别在第7、14、17、24、79—88、127、167、183、202、204位点)、5个苏氨酸(Threonine)磷酸化位点(第6、48、48、126、142位点)、1个酪氨酸(Tyrosine)磷酸化位点(第207位点)。由此推测GhDHN1蛋白的活性可能与其磷酸化调控有关。
2.5棉花GhDHN1编码蛋白与其他植物亲水素蛋白的进化分析
在NCBI上查找到12种植物的亲水素蛋白序列(详见方法1.3.2),采用MEGA5.0邻接法与陆地棉GhDHN1蛋白序列对比并建立无根系统进化树,结果表明,陆地棉GhDHN1蛋白序列与可可亲缘关系最近,其序列一致性为78.70%,其次与柑橘、葡萄柚、山杨的脱水素蛋白亲缘关系也比较近,一致性分别为61.10%、61.40%和63.40%;与拟南芥、玉米、高粱、葡萄、苹果、小麦、油茶、白骨壤等物种的亲水素蛋白的亲缘关系较远(图 5),推测棉花脱水素基因的功能可能与木本植物中脱水素基因功能相似。

图2 GhDHN1蛋白结构分析Fig. 2 Protein stucture analysis of GhDHN1 protein
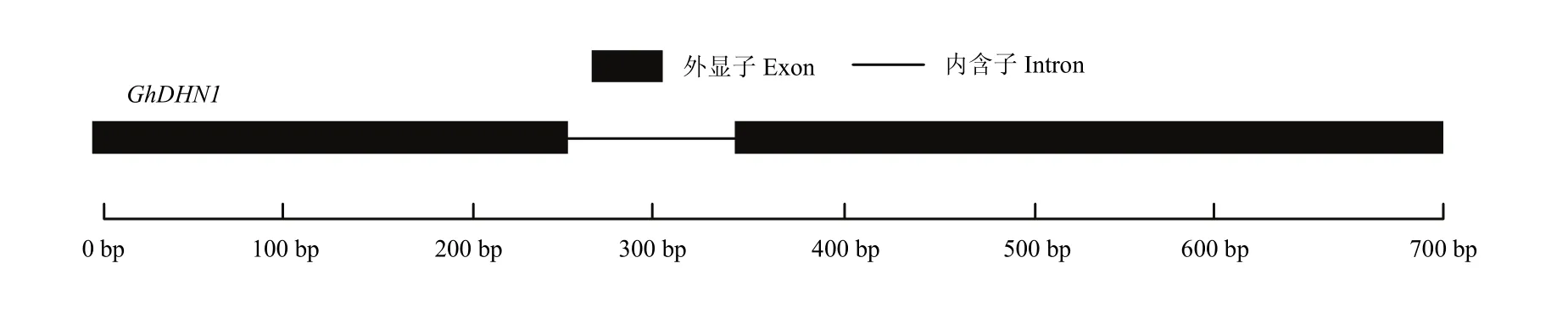
图3 棉花GhDHN1内含子-外显子结构分析Fig. 3 The intron-exon organization of cotton GhDHN1 gene
2.6pBI121-GhDHN1::GFP荧光表达载体的酶切验证
利用pBI121作为瞬时表达载体,利用限制内切酶XbaⅠ和 SmaⅠ,将 GhDHN1插入植物表达载体pBI121:GFP中,构建融合蛋白瞬时表达载体,转化至大肠杆菌DH5α感受态细胞,挑选阳性克隆进行测序,序列比对结果正确。选用限制性内切酶BglⅡ和XhoⅠ酶切验证插入正确(图6),表明表达载体构建成功,命名为pBI121-GhDHN1:GFP。

图4 棉花GhDHN1蛋白的磷酸化位点预测Fig. 4 Protein phosphorylation sites prediction of GhDHN1 protein
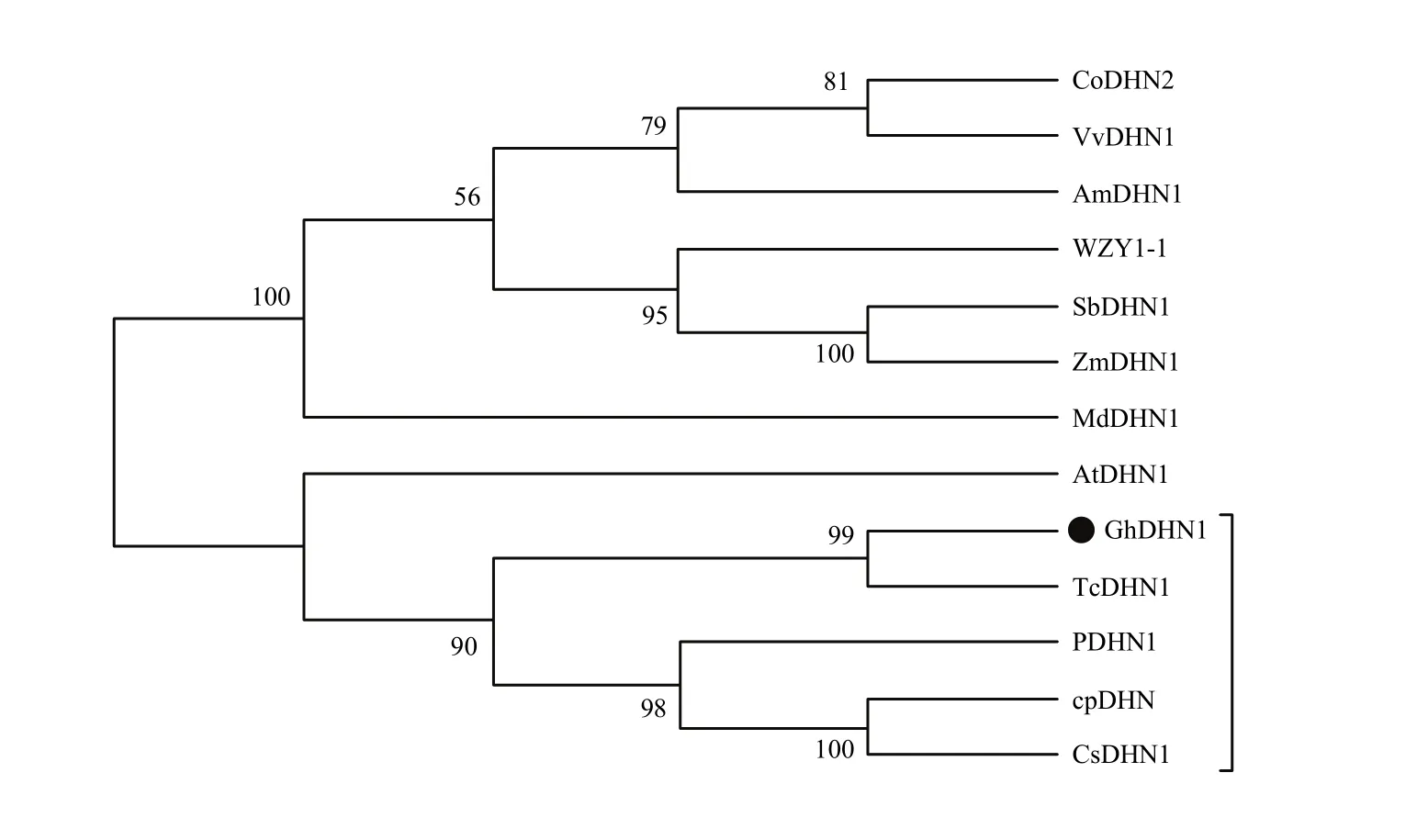
图5 棉花GhDHN1蛋白与12个植物DHN1蛋白的进化树分析Fig. 5 Phylogenetic tree analysis of GhDHN1protein and 12 plant DHN1 proteins
2.7棉花GhDHN1蛋白的亚细胞定位分析
通过瞬时表达系统分析 GhDHN1蛋白亚细胞定位,对照PBI121:GFP的绿色荧光分布在整个细胞中(图7-A—图7-C),而PBI121-GhDHN1:GFP融合蛋白的绿色荧光信号主要集中在细胞膜附近(图 7-D—图7-F),推测GhDHN1蛋白主要在细胞膜附近发挥作用。该基因所编码的蛋白质在洋葱表皮中得到了表达,证明了该基因能够正常表达功能蛋白,为下一步使用基因枪活体转化技术获得转基因拟南芥和转基因棉花材料提供理论依据。
2.8低温胁迫条件下GhDHN1的转录表达分析
基于棉花转录组测序分析表明(图 8),棉花三叶期在受4℃低温处理24 h后,与对照相比,GhDHN1在叶中和根中表达均差异显著,且上调表达,上调倍数分别为2.57和1.29倍,说明GhDHN1棉花响应低温胁迫中起正调控作用。
2.9低温胁迫条件下GhDHN1实时灾光定量分析
实时荧光定量分析表明(图9-A)可知,GhDHN1在受4℃低温处理24 h后,与对照相比,在叶、茎均上调表达,且差异达到极显著,表达倍数分别为14.28 和4.30,在根中也上调表达,且与对照差异显著,上调倍数为1.33,说明GhDHN1在响应低温胁迫过程中,在叶中的作用最大,其次是在茎中和根中,这与转录组测序结果是一致的;4℃低温处理不同时间后,与对照相比,GhDHN1在在叶片中基因表达,除了在0.5 h处理时差异显著外,在其他时间均差异达极显著水平,在4 h和24 h时有2个表达高峰(图9-B);在根中受不同时间低温处理后,与对照相比,在 0.5、6、8 和12 h上调表达且差异达极显著,同时在6 h和12 h 有2个表达高峰(图9-C);GhDHN1在受4℃低温处理24 h后,与对照相比,抗冷材料豫2067和衡棉3号的叶片均上调表达,且与对照间达差异极显著,表达倍数分别为14.28和5.77,在抗冷材料中的表达量远大于冷敏感材料,其表达量是冷敏感材料的2.47倍,二者表达倍数差异极显著(图9-D)。
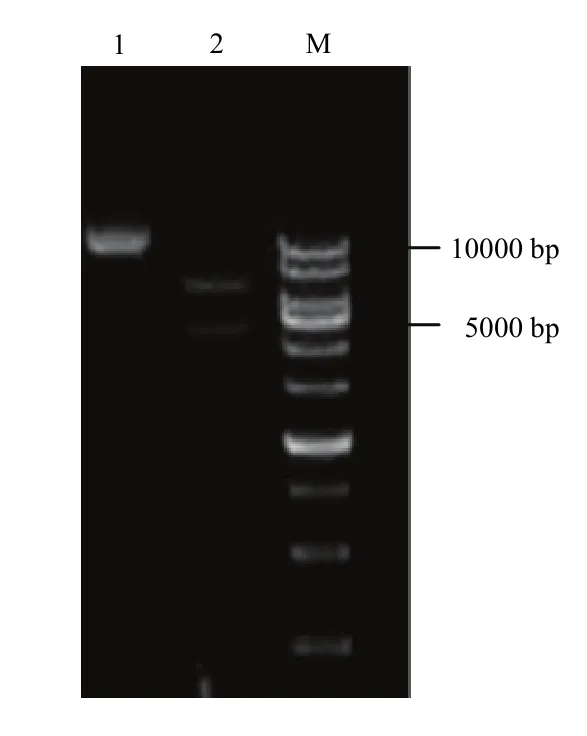
图6 pBI121-GhDHN1::GFP表达载体酶切验证Fig. 6 Enzyme digestion of expression vector pBI121-GhDHN1:GFP
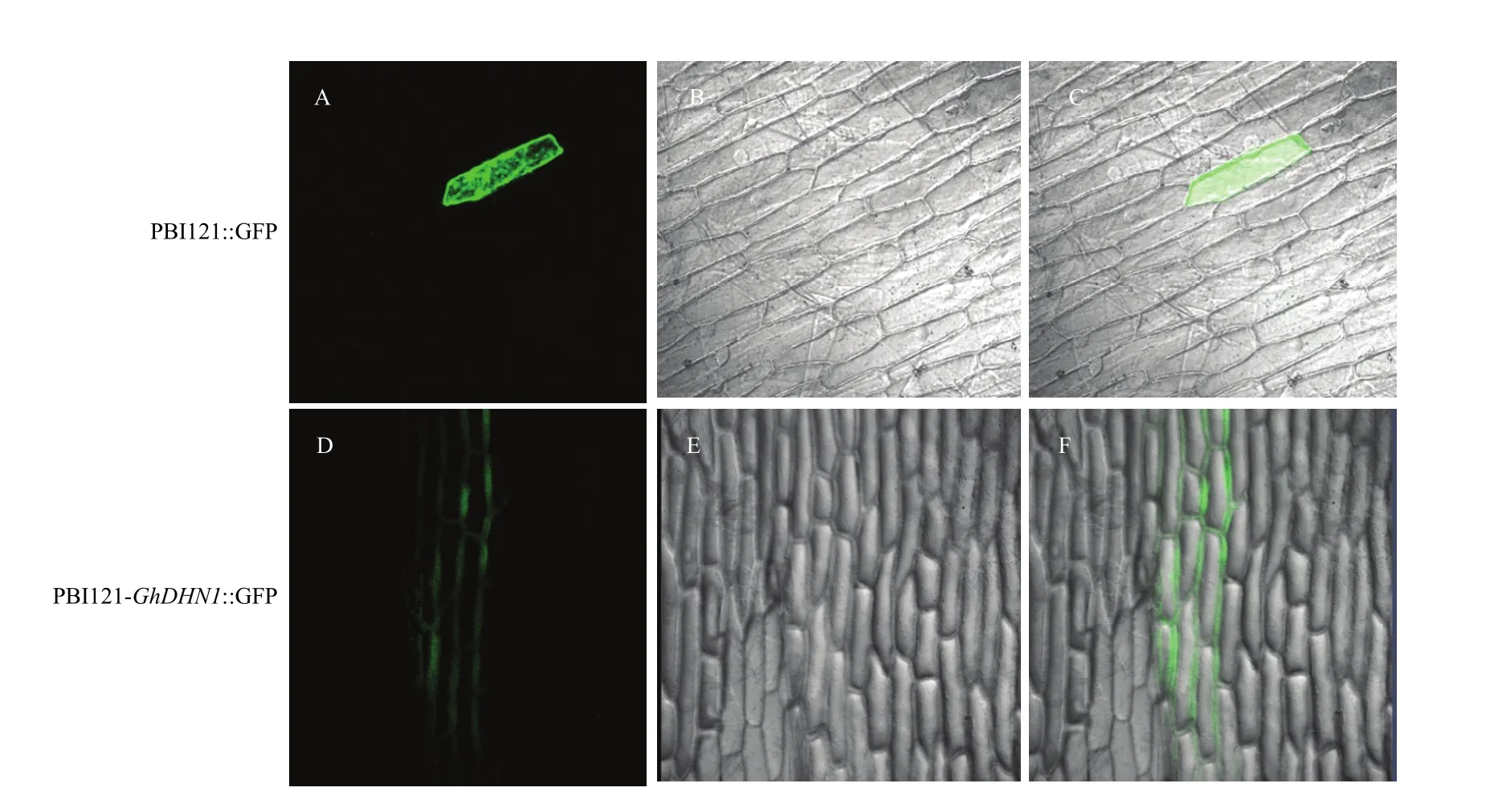
图7 棉花GhDHN1蛋白的亚细胞定位Fig. 7 Sub-cellular localization of GhDHN1 protein
3 讨论
植物在生长发育发过程中面临大量的环境威胁,如生物的和非生物的逆境,面对这些环境的不断变化,它们必须进化出相应的适应机制,才能度过这些不利环境,继续生存下去。合成一系列逆境响应蛋白,小分子代谢物和一些渗透保护物质等可以有助于植物在不同的逆境胁迫条件下存活。积累脱水素是植物在应对逆境胁迫过程的一条主要途径,在植物耐受干旱和盐胁迫过程中发挥重要的保护功能[20]。
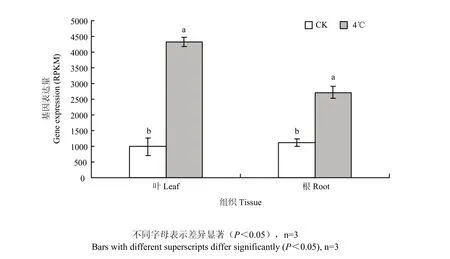
图8 GhDHN1在豫2067三叶期叶片和根系经低温胁迫后的转录丰度Fig. 8 Transcript abundance analysis of GhDHN1 gene in leaves and roots of Yu2067 after treatment at 4℃ low temperature for 24 h
脱水素是一类重要LEA蛋白,通常包含有Y片段、S片段和K片段保守域,其中K片段是所有脱水素均具备的特征结构域[21]。研究表明,富含赖氨酸K片段一般位于蛋白序列的C端,可以形成双亲α螺旋,α螺旋能够和其他蛋白的脱水表面或生物膜表面发生互作,这种互作能保护其他蛋白水分的丢失,在细胞脱水保护过程中发挥重要功能[22]。在本研究中发现,GhDHN1蛋白在C端具有2个K片段保守域,二级结构分析显示该蛋白的α螺旋(alpha helix)占到整个蛋白54.98%,无规则卷曲(random coil)占到整个蛋白的41.23%,FAN等[23]研究表明这种无规则卷曲结构对于增加细胞中束缚水含量,保护细胞脱水功能非常有利。S片段是由一系列丝氨酸残基组成,ALSHEIKH等[24]研究表明S片段的磷酸化可以使脱水素在信号肽的引导下进入细胞核,GhDHN1在N端具有一个S片段,共含有16个丝氨酸(Serine)磷酸化位点,5个苏氨酸(Threonine)磷酸化位点,1个酪氨酸(Tyrosine)磷酸化位点,推测这些磷酸化位点在棉花脱水素的功能行使过程中发挥重要作用,这一功能还有待于进一步的验证。研究表明,GhDHN1蛋白呈酸性且带负带荷,亚细胞定位于细胞质膜附近,DANYLUK等[25]报道,小麦冷诱导脱水素基因也呈酸性,在细胞质膜上被检测到,因此,2个物种的脱水素蛋白有相似的性质和亚细胞定位,推测可能具有相似的功能。
大量研究表明在非生物逆境胁迫下,植物脱水素的表达和积累与植物抗逆性之间存在着正相关关系[26-28]。本研究转录组测序结果表明,GhDHN1受低温胁迫棉花叶片和根中均上调表达;实时荧光定量分析表明,GhDHN1在受低温胁迫的根、茎、叶中均上调表达,特别是在叶中上调表达的更为明显,这些结果充分说明GhDHN1受低温的诱导,这跟GhDHN1蛋白的性质有一定相关性,据报道,酸性或中性的SKn、Kn、Y2Kn型脱水素蛋白优先可被低温诱导,如小麦WCO410属于酸性的K3S型脱水素,可在维管组织中受低温诱导[25];另据报道,脱水素在抵御低温等非生物胁迫过程发挥着重要作用[29],有研究表明过量表达脱水素基因可增强拟南芥的低温抗性,提高其抗寒力[30];GhDHN1在低温胁4 h和24 h在棉花的叶片中上调倍数2次达到高峰,这与徐丽等[31]在核桃中的研究结果相似,徐丽等研究表明,JrDHN在4℃胁迫下核桃叶片中表达增加,4℃处理4 h后达到最大值;这些结果与花生中K2S型脱水素基因AhDHN1的功能不完全一样,AhDHN1对高盐和干旱胁迫则有明显响应,对低温胁迫无响应[32],推测可能是因为花生中该脱水素基因序列与棉花中该基因序列差异较大造成的;GhDHN1在抗冷材料和冷敏感材料中表达差异显著,其表达量与棉花的抗冷性呈正相关,这与小麦脱水素有相似的功能,研究表明,在适合生长的温度条件下,小麦脱水素在转录水平和蛋白水平在抗冻性强的小麦品种中的积累远多于不抗冻品种,所以在温度降低过程中具有更高的适应性,具有更强的抗冷性,因此,脱水素可以作为不同材料间的抗冷性筛选鉴定标记[33]。以上结果表明,脱水素蛋白在棉花响应低温胁迫过程中可能具有复杂的响应模式。
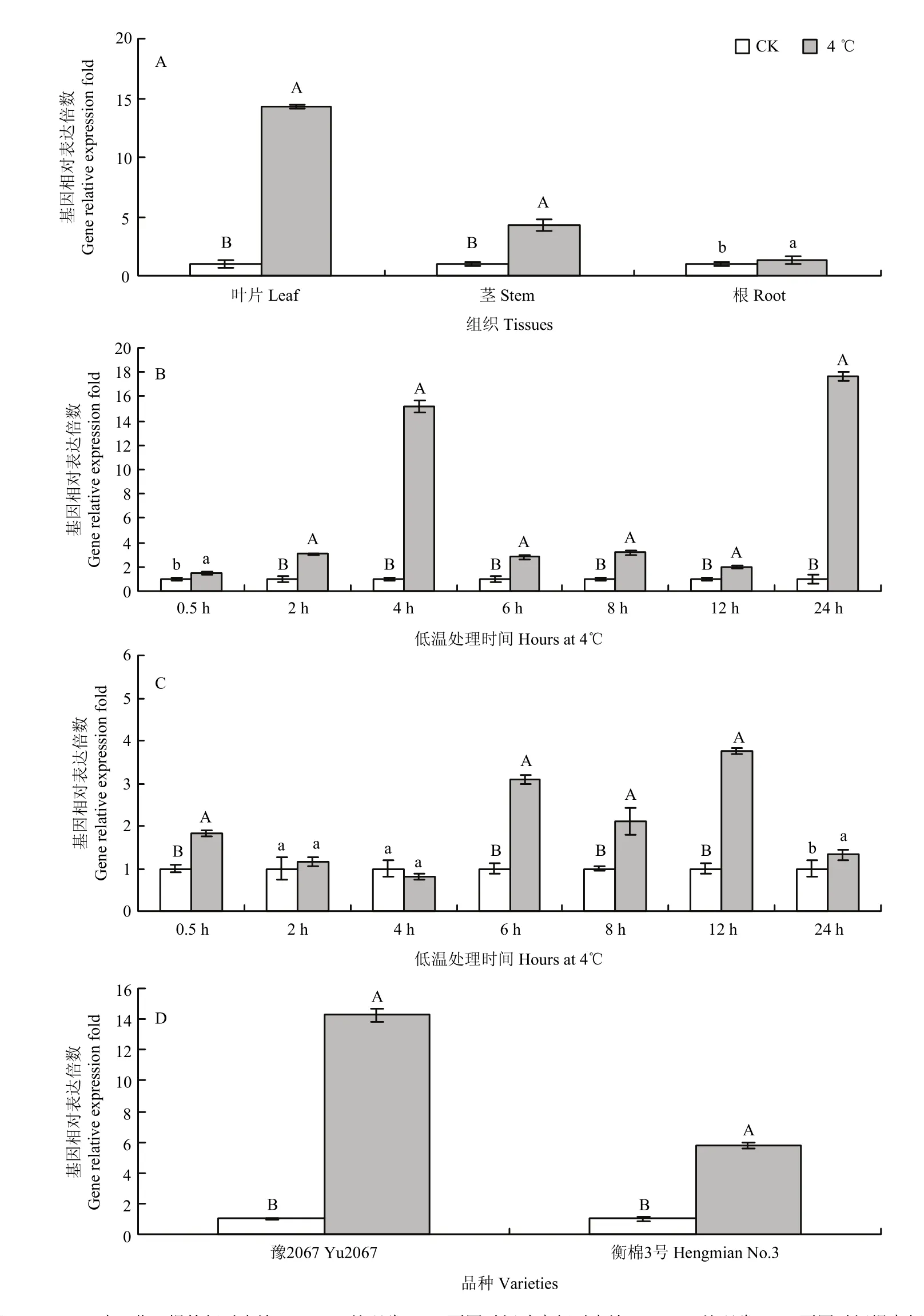
图9 低温胁迫条件下GhDHN1实时荧光定量分析Fig.9 Expression of GhDHN1 gene in cotton under low temperature stress treatment with qRT-PCR method
4 结论
陆地棉GhDHN1属于典型的K2S型脱水素,与可可亲缘关系最近,在响应低温胁迫中起正调控作用,在抗冷材料和冷敏感材料中表达差异极显著,其表达量与棉花的抗冷性呈正相关,可以作为筛选不同抗冷材料的分子标记,同时可以作为培育棉花抗冷材料的候选基因。
References
[1] 李新国, 毕玉平, 赵世杰, 孟庆伟, 何启伟, 邹琦. 短时低温胁迫对甜椒叶绿体超微结构和光系统的影响. 中国农业科学, 2005,38(6): 1226-1231. LI X G, BI Y P, ZHAO S J, MENG Q W, HE Q W, ZOU Q. Effects of short-term chilling stress on the photosystems and chloroplast ultrastructure in sweet pepper. Scientia Agricultura Sinica, 2005,38(6): 1226-1231. (in Chinese)
[2] SAWAN Z M, HANNA L I, GAD E, KARIM G A, MCCUISTION W L. Relationships between climatic factors and flower and boll production in Egyptian cotton (Gossypium barbadense). Journal of Arid Environments, 2002, 52(4): 499-516.
[3] POSAS F, CHAMBERS J R, HEYMAN J A, HOEFFLER J P, DE NADAK E, ARIÑO J. The transcriptional response of yeast to saline stress. The Journal of Biological Chemistry, 2000, 275(23):17249-17255.
[4] SHINOZAKI K, YAMAGUCHI-SHINOZAKI K. Gene networks involved in drought stress response and tolerance. Journal of Experimental Botany, 2007, 58(2): 221-227.
[5] BIES ETHÈVE N, GAUBIER-COMELLA P, DEBURES A,LASSERRE E, JOBET E, RAYNAL M, COOKE R, DELSENY M. Inventory, evolution and expression profiling diversity of the LEA (late embryogenesis abundant) protein gene family in Arabidopsis thaliana. Plant Molecular Biology, 2008, 67(1): 107-124.
[6] DURE L, CROUCH M, HARADA J, DAVID HO T H, MUNDY J,QUATRANO R, THOMAS T, SUNG Z R. Common amino acid sequence domains among the LEA proteins of higher plants. Plant Molecular Biology, 1989, 12(12): 475-486.
[7] HANIN M, BRINI F, EBEL C, TODA Y, TAKEDA S, MASMOUDI K. Plant dehydrins and stress tolerance: Versatile proteins for complex mechanisms. Plant Signaling & Behavior, 2011, 6(10): 1503-1509.
[8] HARA M, FUJINAGA M, KUBOI T. Radical scavenging activity and oxidative modification of citrus dehydrin. Plant Physiology Biochemistry, 2004, 42(7/8): 657-662.
[9] CHAKRABORTEE S, BOSCHETTI C, WALTON L J, SARKAR S,RUBINSZTEIN D C, TUNNACLIFFE A. Hydrophilic protein associated with desiccation tolerance exhibits broad protein stabilization function. Proceedings of the National Academy of Sciences of the USA, 2007, 104(46): 18073-18078.
[10] KOSOVÁ K, HOLKOVÁ L, PRÁŠIL I T, PRÁŠILOVÁ P,BRADÁČOVÁ M, VÍTÁMVÁS P, ČAPKOVÁ V. Expression of dehydrin 5 during the development of frost tolerance in barley (Hordeum vulgare). Journal of Plant Physiology, 2008, 165(11):1142-1151. (in Chinese)
[11] 郭鹏, 张士刚, 金华, 邹吉祥, 董燕, 姜国斌. 番茄脱水素基因SlDHN2b的克隆与表达分析. 园艺学报, 2012, 39(10): 2015-2022. GUO P, ZHANG S G, JIN H, ZOU J X, DONG Y, JIANG G B. Cloning and characterization of dehydrins gene SlDHN2b in tomato. Acta Horticulturae Sinica, 2012, 39(10): 2015-2022. (in Chinese)
[12] 张宁, 孙敏善, 刘露露, 孟凡荣, 任江萍, 尹钧, 李永春. 小麦脱水素基因 TaDHN-1的特征及其对非生物胁迫响应. 中国农业科学,2013, 46(4): 849-858. ZHANG N, SUN M S, LIU L L, MENG F R, REN J P, YIN J, LI Y C. Characterization of a dehydrin gene TaDHN-1 and its response to abiotic stresses in wheat. Scientia Agricultura Sinica, 2013, 46(4):849-858. (in Chinese)
[13] PUHAKAINEN T, HESS M W, MAKELA P, SVENSSON J, HEINO P, PALVA E T. Overexpression of multiple dehydrin genes enhances tolerance to freezing stress in Arabidopsis. Plant Molecular Biology,2004, 54(5): 743-753.
[14] HALDER T, AGARWAL T, RAY S. Isolation, cloning, and characterization of a novel dehydrin (SbDhn2) protein. Protoplasma,2015, 11(4): 1-14.
[15] ZHANG L, OHTA A, TAKAGI M, IMAI R. Expression of plant group 2 and group 3 lea genes in Saccharomyces cerevisiae revealed functional divergence among LEA proteins. Journal Biochemistry,2000, 127(4): 611-616.
[16] SWIRE-CLARK G A, MARCOTTE W R. The wheat LEA protein Em functions as an osmoprotective molecule in Saccharomyces cerevisiae. Plant Molecular Biology, 1999, 39(1): 117-128.
[17] MORTAZAVI A, WILLIAMS B A, MCCUE K, SCHAEFFER L,WOLD B. Mapping and quantifying mammalian transcriptomes byRNA-Seq. Nature Methods, 2008, 5(7): 621-628.
[18] AFRIN S, ZHU J, CAO H Z, HUANG J G, XIU H, LUO T, LUO Z Y. Molecular cloning and expression profile of an abiotic stress and hormone responsive MYB transcription factor gene from Panax ginseng. Acta Biochimica et Biophysica Sinica, 2015, 47(4): 267-277.
[19] CLOSE T J, KORTT A A, CHANDLER P M. A cDNA-based comparison of dehydration-induced proteins (dehydrins) in barley and corn. Plant Molecular Biology, 1989, 13(1): 95-108.
[20] HUNDERTMARK M, HINCHA D K. LEA (late embryogenesis abundant) proteins and their encoding genes in Arabidopsis thaliana. BMC Genomics, 2008, 9: 118.
[21] ALLAGULOVA C R, GIMALOV F R, SHAKIROVA F M,VAKHITOV V A. The plant dehydrins: Structure and putative functions. Biochemistry, 2003, 68(9): 945-951.
[22] HANIN M, BRINI F, EBEL C, TODA Y, TAKEDA S, MASMOUDI K. Plant dehydrins and stress tolerance: Versatile proteins for complex mechanisms. Plant Signaling and Behavior, 2011, 6(10): 1503-1509.
[23] FAN Z, WANG X. Isolation and characterization of a novel dehydrin gene from Capsella bursa-pastoris. Molecular Biology, 2006, 40(1):43-50.
[24] ALSHEIKH M K, HEYEN B J, RANDALL S K. Ion binding properties of the dehydrin ERD14 are dependent upon phosphorylation. Journal of Biology Chemistry, 2003, 278(42) : 40882- 40889.
[25] DANYLUK J, PERRON A, HOUDE M, LIMIN A, FOWLER B,BENHAMOU N, SARHAN F. Accumulation of an acidic dehydrin in the vicinity of the plasma membrane during cold acclimation of wheat. The Plant Cell, 1998, 10(4): 623-638.
[26] WAHID A, CLOSE T J. Expression of dehydrins under heat stress and their relationship with water relations of sugarcane leaves.Biologia Plantarum, 2007, 51(1): 104-109.
[27] XU J, ZHANG Y X, GUAN Z Q, WEI W, HAN L, CHAI T Y. Expression and function of two dehydrins under environmental stresses in Brassica juncea L.. Molecular Breeding, 2008, 21(4):431-438.
[28] MINGEOT D, DAUCHOT N, VAN CUTSEM P, WATILLON B. Characterisation of two cold induced dehydrin genes from Cichorium intybus L.. Molecular Biology Reports, 2009, 36(7): 1995-2001.
[29] CLOSE T J. Dehydrins: A commonality in the response of plants to dehydration and low temperatures. Physiologia Plantarum, 1997,100(2): 291-296.
[30] PUHAKAINEN T, HESS M W, MÄKELÄ P, SVENSSON J, HEINO P, PALVA E T. Overexpression of multiple dehydrin genes enhances tolerance to freezing stress in Arabidopsis. Plant Molecular Biology,2004, 54(5): 743-753.
[31] 徐丽, 陈新, 魏海蓉, 张力思, 宗晓娟, 王甲威, 朱东姿, 刘庆忠.核桃Y2SK2型脱水素基因JrDHN的克隆、表达和单核苷酸多态性分析. 园艺学报, 2014, 41(8): 1573-1582. XU L, CHEN X, WEI H R, ZHANG L S, ZONG X J, WANG J W,ZHU D Z, LIU Q Z. Molecular cloning, expression and single nucleotide polymorphisms analysis of typical Y2SK2dehydrin in Juglans. Acta Horticulturae Sinica, 2014, 41(8): 1573-1582. (in Chinese)
[32] 陈娜, 胡冬青, 潘丽娟, 迟晓元, 陈明娜, 王通, 王冕, 杨珍, 禹山林. 花生中胁迫相关基因AhDHN1的克隆及非生物胁迫下表达分析. 核农学报, 2014, 28(12): 2159-2166. CHEN N, HU D Q, PAN L J, CHI X Y, CHEN M N, WANG T,WANG M, YANG Z, YU S L. Cloning of a Dehydrin Gene AhDHN1 and its expression analysis during abiotic stresses in peanut. Journal of Nuclear Agricultural Sciences, 2014, 28(12): 2159-2166. (in Chinese)
[33] GALIBA G, VÁGÚJFALVI A, LI C, SOLTÉSZ A, DUBCOVSKY J. Regulatory genes involved in the determination of frost tolerance in temperate cereals. Plant Science, 2009, 176(1): 12-19.
(责任编辑 李莉)
Molecular Clone and Expression of GhDHN1 Gene in Cotton (Gossypium hirsutum L.)
WANG Jun-juan, MU Min, WANG Shuai, LU Xu-ke, CHEN Xiu-gui, WANG De-long, FAN Wei-li,YIN Zu-jun, GUO Li-xue, YE Wu-wei, YU Shu-xun
(Institute of Cotton Research, Chinese Academy of Agricultural Sciences/State Key Laboratory of Cotton Biology/Key Laboratory for Cotton Genetic Improvement, Ministry of Agriculture, Anyang 455000,Henan)
【Objective】In order to explore functional genes related to low temperature stress tolerance of cotton, the characteristics of cotton dehydrin gene and its expression patterns responsed to low temperature stress in cotton were analyzed, thus providing a theoretical basis for the application of dehydrin gene in cotton chilling tolerant breeding. 【Method】 In this study, based on the upland cotton genome sequence, specific primers were designed by Primer 5 software and the dehydrin gene was cloned from the upland cotton variety Yu2067, named GhDHN1. Bioinformatics analysis was conducted to analyze the properties, amino acids content, functional domains and evolutionary relationships of the gene. Plant expression vector pBI121:GFP at Xbaand Ⅰ SmaⅠrestriction site s was constructed with double enzyme digestions and the transient expression vector pBI121-GhDHN1::GFP was constructed by In-Fusion connection technology. And the subcellular localization of GhDHN1 was studied by transient expression analysis of onion epidermal cells. Combined with the transcriptome sequencing data of chilling-resistant cotton variety Yu2067, real-time fluorescent quantitative PCR expression of leaves, stems, and roots in chilling-resistant Yu2067 and chilling-sensitive variety Hengmian No.3 under low temperature stress treatments (4℃, 24 h) at trefoil stage was performed to study the function of GhDHN1. The expression of the gene in the leaves of two different cold resistant varieties was compared. The dynamic expression of GhDHN1 gene in leaves and roots of Yu2067 was detected under 4℃ low temperature treatment.【Result】 A cotton dehydrin gene was cloned and the sequencing analysis showed that the cDNA of the dehydrin gene was 726 bp, and the gene encoded 211 amino acids with a predicted molecular weight of about 23.79 kD and the isoelectric point was 5.04. Amino acid sequence analysis indicated that the GhDHN1 was rich in glutamic acid content (26.10%) and lysine amino acid content (19.40%)with a half-life of 30 hours. GhDHN1 was acidic and negatively charged, of which 60 percent were negatively charged residues. The second structure analysis showed that the alpha helix of GhDHN1 contained 116 amino acid residues, accounting for 54.98% of the protein composition and random coil contained 87 amino acid residues. GhDHN1 gene was located on Dt_chr9 chromosome of AD genome. GhDHN1 gene contained a 90 bp intron at 259-348 position of cDNA and two exons in lengths of 258 bp and 378 bp,respectively. SMART and CDD analysis showed that GhDHD1 includes two conserved lysine-rich K fragments, a conserved serine-rich S fragment and a dehydrin functional domain pfam00257 and therefore was sorted into K2S subfamily of dehydrins. Phylogenetic analysis showed that GhDHN1 of cotton had the closest relationship with DHN1 of cacao. The analysis of the transient expression in onion epidermal cells showed that GhDHN1 was mainly localized near the cell plasma membrane. Transcriptome analysis showed that GhDHN1 gene was up-regulated in leaves and roots of cotton after low temperature treatments at the trefoil stage. qRT-PCR analysis showed that GhDHN1 was up-regulated in leaves, stems and roots after low temperature. The expression fold in leaves was higher than that in stems and roots. There were two expression peaks in leaves under low temperature treatment for 4 h and 24 h, and there were also two peaks in roots of the low temperature treatment for 6 h and 12 h. The leaf expression of the gene in cold resistant varieties was 2.47 times the expression in the cold sensitive varieties. The results showed that GhDHN1 gene may be involved in the adaptability regulation of low temperature. 【Conclusion】 GhDHN1 belonged to the member of K2S subfamily of dehydrins. Phylogenetic analysis showed that GhDHN1 had the closest relationship with DHN1 of cacao. GhDHN1 gene was induced by low temperature stress. Significant expression difference of GhDHN1 between the cold-resistance variety and cold-sensitive variety was found, and the expression of GhDHN1 was positively correlated with cotton cold-resistance. The expression of GhDHN1 can be thus employed as a marker of cotton chilling resistance. At the same time GhDHN1 gene can also serve as an important candidate gene in cotton cultivation of cold-resistant materials.
cotton; dehydrin; sub-cellular localization; low temperature stress; transcriptional expression; real-time fluorescent quantitative PCR
2016-02-24;接受日期:2016-05-24
河南省基础与前沿技术研究计划项目(142300413232)
联系方式:王俊娟,Tel:15093920872;E-mail:wjj2004liyuan@sina.com。通信作者喻树迅,Tel:0372-2562201;E-mail:ysx195311@163.com
