GGTA1-/-五指山小型猪SLAI类基因分子特征及与HLA相似性分析
2016-09-13蒋应弟曾国敏石宁宁李西睿纪慧丽潘登科滚双宝
蒋应弟,曾国敏,石宁宁,李西睿,纪慧丽,潘登科*,滚双宝
(1. 甘肃农业大学动物科学技术学院,兰州 730070;2.中国农业科学院北京畜牧兽医研究所农业部畜禽遗传资源与种质创新重点实验室,北京 100193;3. 甘肃农业大学生命科学技术学院,兰州 730070)
GGTA1-/-五指山小型猪SLAI类基因分子特征及与HLA相似性分析
蒋应弟1,2,曾国敏2,3,石宁宁2,李西睿2,纪慧丽2,潘登科2*,滚双宝1*
(1. 甘肃农业大学动物科学技术学院,兰州730070;2.中国农业科学院北京畜牧兽医研究所农业部畜禽遗传资源与种质创新重点实验室,北京100193;3. 甘肃农业大学生命科学技术学院,兰州730070)
目的明晰α-1,3半乳糖基转移酶基因敲除(GGTA1-/-)的五指山小型猪SLA经典I类基因分子结构特征及其与人HLA的相似性,对研究异种器官移植细胞性排斥反应具有重要意义。 方法 采集6头祖代GGTA1-/-五指山小型猪耳组织,利用RT-PCR对SLAI类基因(SLA-1、SLA-3、SLA-2)扩增、克隆及测序,对序列进行BLAST分析,并采用生物信息学方法对SLAI类基因分子结构特征及其与人HLA的同源性进行分析。 结果测序分析表明,共获得6条等位基因序列,其中4条为已公布的等位基因(SLA-1*0703、SLA-2*1102、SLA-3*0401、SLA-3*0403),另2条为新等位基因(SLA-1*0401wz01、SLA-2*11wz01)。五指山小型猪与人HLA同源性为70.5%~72.1%。SLA-1*0401wz01、SLA-1*0703、SLA-2*11wz01、SLA-2*1102和SLA-3*0401的CD8+分子识别的关键结合域与人HLA氨基酸序列比对,均仅在位点225、228处发生了突变(T→S、T→M),其他位点高度保守。SLA-2*11wz01和SLA-2*1102与人HLAI类基因NK细胞抑制性受体(killerinhibitoryreceptor,KIR)结合区氨基酸同源性较高,在NKTA-1亚型结合域内仅有1个氨基酸差异,而在NKTA-2 和NKTA-3亚型结合域内有2个氨基酸差异。 结论从免疫细胞介导的异种排斥反应角度出发,GGTA1-/-五指山小型猪SLAI类基因氨基酸序列与人HLA高度相似,可作为未来猪-人异种器官移植的良好供体之一。
GGTA1-/-;五指山小型猪;SLA;异种移植;免疫排斥
α-1,3半乳糖基转移酶基因敲除(GGTA1-/-)猪的诞生有效克服了异种移植时超急性排斥反应的发生,随后细胞性排斥成为目前亟待解决的问题[1-4]。免疫细胞介导的异种排斥反应可分为适应性细胞反应和固有细胞免疫反应。其中适应性免疫细胞主要指T淋巴细胞,而固有免疫细胞包括自然杀伤(naturalkiller,NK)细胞、单核吞噬细胞、中性粒细胞和树突状细胞(DC)等。研究报道猪白细胞抗原(swineleukocyteantigen,SLA)是引起异种移植细胞性免疫排斥的遗传因素之一[5-7]。异种移植引起的CD8+T细胞免疫排斥反应的大小主要取决于供体细胞表面SLAⅠ类分子与被CD8+分子识别的HLA分子的同源性[8-9]。人NK细胞的活性受表面抑制性受体的调控。当抑制受体识别自身MHCI分子后,对NK细胞毒作用产生抑制性信号,避免NK细胞对自体细胞的攻击[10]。但异种移植时,人NK细胞抑制性受体不能识别SLAI分子,因而不能向NK细胞受体提供抑制性信号,从而产生抗猪的NK细胞毒作用[11]。
五指山小型猪是目前我国异种器官移植研究常用实验动物之一。国内关于五指山小型猪SLAI类基因的研究大多集中在多态性较高的外显子或单个基因的克隆分析[12-14],对SLAI类基因全长cDNA序列结构特征分析及与人HLA相似性报道较少。利用体细胞核移植技术获得的GGTA1-/-五指山小型猪已被多次应用于异种器官移植临床前研究[15-16],鉴于此,本研究以GGTA1-/-五指山小型猪为研究对象,通过对其SLA经典I类基因(SLA-1、SLA-2、SLA-3)分子结构特征研究及与人HLA相似性分析,为筛选更适合异种移植研究的GGTA1-/-五指山小型猪提供参考依据。
1 材料与方法
1.1 材料
采集6头祖代7~8月龄、体重在10~40kg普通级的GGTA1-/-五指山小型猪耳组织样本,液氮中保存。
动物组织总RNA提取试剂盒购自北京天根生化科技有限公司;反转录试剂盒、pEASY-T1SimpleCloningKit购自北京全式金生物技术有限公司;TaKaRaLATaq 聚合酶、2×GCBufferII购自宝生物工程(大连)有限公司;DNA纯化回收试剂盒、DH10b感受态细胞购自中美泰和生物技术(北京)有限公司。
1.2方法
1.2.1RNA的提取及反转录
提取样本的总RNA;用1.5%琼脂糖凝胶电泳检测总RNA质量;采用反转录试剂盒将RNA反转录为cDNA,-20℃保存备用。
1.2.2SLA-1、SLA-3和SLA-2基因的扩增
参考GenBank中SLAI类基因序列,用Lasergene7.1软件包中的PrimerSelect软件分别设计SLA-1和SLA-3扩增引物,SLA-2扩增引物引用Gao等[19]的研究,引物由中美泰和生物技术(北京)有限公司合成。引物序列及退火温度见表1。PCR反应体系:10.0μL2×GCbufferII,上下游引物(10μmol/L)各1.0μL,0.4μLdNTPs(10mmol/L), 0.1μLLaTaq酶,1.0μLcDNA模板,6.5μLddH2O,总反应体系为20μL。PCR扩增程序: 95℃预变性 5min;95℃变性 30s,表1中各退火温度 30s,72℃延伸80s,35个循环;72℃延伸10min,16℃保存。

表1 SLAⅠ类基因的扩增引物
1.2.3基因的克隆及序列分析
PCR产物用纯化试剂盒切胶回收,克隆并经阳性鉴定后送中美泰和生物技术(北京)有限公司进行双向测序,每个基因座筛选至少8个以上阳性单克隆菌液进行测序,利用Lasergene7.1软件包中的SeqMan程序对获得的序列进行剪切拼接,拼接序列通过BLAST(http://www.ncbi.nlm.nih.gov/BLAST/)比对分析,新等位基因的序列提交至GenBank并获取相应序列号。
1.2.4同源性分析
使用MegAlign软件在氨基酸水平上对五指山小型猪与NIH小型猪、人和鼠SLAI类等位基因同源性分析,且与被CD8+分子识别的HLA关键结合域氨基酸相似性、与HLAⅠ类基因KIR结合区氨基酸相似性进行比较。
2 结果与分析
2.1SLAⅠ类基因的扩增

图1 总RNA(A)及SLA-1(B.1)、SLA-2(B.2)、SLA-3(B.3)基因Fig.1 A: Agarose gel assay of the total RNA; B: Agarose gel assay of RT-PCR amplification of SLA-1, SLA-2 and SLA-3 genes
GGTA1-/-五指山小型猪耳组织总RNA电泳结果(图1A),显示:RNA质量较好;SLA-1、SLA-2和SLA-3均有特异性扩增条带,大小分别为1200bp、1231bp和1300bp,与预期扩增结果一致(图1B)。2.2SLAI类基因序列分析
在GGTA1-/-五指山小型猪SLA-1、SLA-2、SLA-3基因座上均获得2条序列,经BLAST分析,SLA-1基因中的1条为新等位基因,将序列提交于GenBank,获得其序列号:SLA-1*0401wz01 (AccessionNo:KT715501),另1条SLA-1*0703已报道;SLA-3基因中2条等位基因序列均已被报道,分别为SLA-3*0401 (AccessionNo:AF464011 )、SLA-3*0403 (AccessionNo:KT715502);SLA-2基因中的SLA-2*1102 (AccessionNo:JQ361654 )为已发现的等位基因,另外1条为新的等位基因,将其提交于GenBank,获得序列号:SLA-2*11wz01 (AccessionNo:KT194213)。将SLA-1和SLA-2基因座上的新等位基因分别与NCBI中相应等位基因核苷酸比对发现新等位基因的高变异位点主要集中在第2、3外显子上,其他位点相对保守。
2.3SLAⅠ类基因在个体中的分布
6头GGTA1-/-五指山小型猪个体中SLA经典Ⅰ类基因座上共获得6条等位基因序列,6个等位基因在个体中的分布情况见表2。
2.4SLAI类等位基因的同源性分析
将本研究获得等位基因序列与美国NIH小型猪、人及鼠相应等位基因在氨基酸水平进行同源性比较,发现五指山小型猪SLAI类等位基因序列与NIH小型猪在氨基酸水平上同源性在83.2% ~100%之间,与人HLA同源性为70.5% ~72.1%。SLA-1*0401wz01与NIH猪SLA-1*0401相比,核苷酸序列仅在位点564处发生同义突变(G→C),与人HLA同源性高达70.8%;SLA-2基因座上等位基因SLA-2*11wz01、SLA-2*1102与NIH小型猪相应等位基因同源性高达92.1%,与人HLA同源性高达71.3%,与鼠的同源性仅为62.2%;SLA-3基因座上等位基因SLA-3*0401、SLA-3*0403与NIH小型猪相应等位基因同源性高达94.5%,与人的同源性高达72.1%。各等位基因的同源性比对情况见表3。
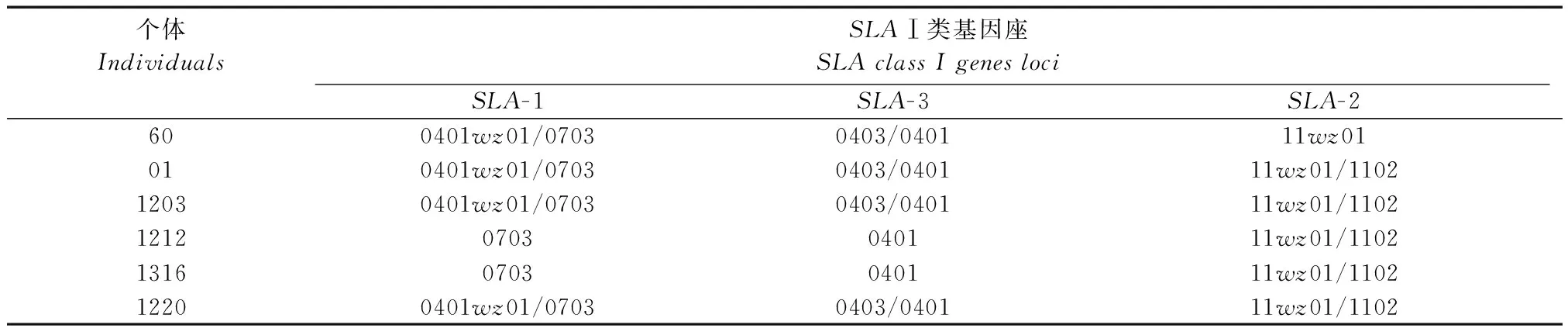
表2 SLA I类基因座上等位基因的分布

表3 SLA-1、SLA-2、SLA-3等位基因氨基酸同源性比较
注:A*0201表示人群中出现频率较高的HLA-A*0201基因;斜体表示NIH小型猪SLA-1、SLA-2、SLA-3基因座上出现频率较高的等位基因,依次为:SLA-1*0701(AF464036)、SLA-1*0401(AF464002)、SLA-2*0401(AF014006)、SLA-2*0301(AF014004)、SLA-3*0301(AF014003)、SLA-3*0602(DQ303227)。H-2Db表示鼠的相应等位基因。
Note.A*0201standsforHLA-A*0201,aclassIallelewiththehighestfrequencyinhumanpopulations.Italicstandsforalleles,aclassIallelewiththehighestfrequencyintheNIHpopulationsminipigs,asfollows:SLA-1*0701 (AF464036),SLA-1*0401 (AF464002),SLA-2*0401 (AF014006),SLA-2*0301 (AF014004),SLA-3*0301 (AF014003),andSLA-3*0602 (DQ303227).H-2Dbstandsforcorrespondingmousealleles.
2.5SLAI类分子与被CD8+分子识别的HLA分子序列的同源性比较
将五指山小型猪SLAI类SLA-1、SLA-2、SLA-3座位上的等位基因推测得到氨基酸序列与被CD8+分子识别结合的HLA-A*0201关键氨基酸序列区域进行比对(表4)。结果显示,SLA-1*0401wz01、SLA-1*0703、SLA-3*0401、SLA-2*1102和SLA-2*11wz01等位基因都仅在位点225和228处发生突变,由苏氨酸(Thr)→丝氨酸(Ser)、苏氨酸(Thr)→蛋氨酸(Met),说明SLA-1*0401wz01、SLA-1*0703、SLA-3*0401SLA-2*1102和SLA-2*11wz01与人CD8+受体结合所必需的关键氨基酸残基高度相似。但SLA-3*0403等位基因在位点225处由苏氨酸(Thr)→甘氨酸(Gly),在位点228处由苏氨酸(Thr)→蛋氨酸(Met),并在位点227~228之间插入4个氨基酸,依次为:谷氨酰胺(GIn)、丝氨酸(Ser)、谷氨酰胺(GIn)、酪氨酸(Tyr)。鼠的H-2Db等位基因氨基酸序列分别与被CD8+分子识别并结合的HLA-A*0201关键氨基酸序列区域比对发现,H-2Db有4个位点发生了突变。
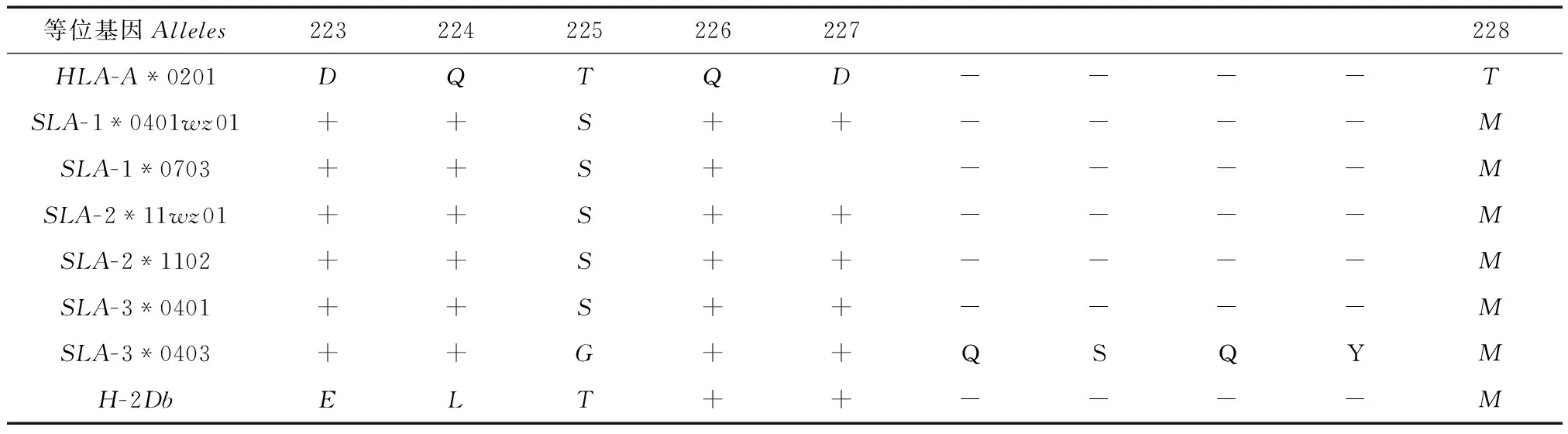
表4 五指山小型猪SLA I类分子与被CD8+分子识别的HLA氨基酸序列的同源性比较
注:斜体字母表示插入的氨基酸;“+”表示与人的HLA-A*0201氨基酸相同;“-”表示缺失的氨基酸;HLA-A*0201人群中出现频率较高的等位基因。
Note.Theitalicstandsforinsertedaminoacids. “+”indicatesthattheaminoacidsareidenticaltohumanHLA-A*0201. “-”indicatesthatthedeletedaminoacid.A*0201standsforHLA-A*0201,aclassIallelewiththehighestfrequencyinhumanpopulations.
2.6SLAI类基因与HLAI类基因KIR结合区重要氨基酸的比较分析
表5为SLAI类基因与HLAI类基因KIR结合区关键氨基酸的比较分析。从表中可以看出,SLA-2*11wz01和SLA-2*1102与人HLAI类基因KIR结合区氨基酸同源性较高,在NKTA-1亚型结合域内仅有1个氨基酸差异,而在NKTA-2 和NKTA-3亚型结合域内存在2个氨基酸差异。SLA-1*0401wz01基因与人HLAI类基因KIR结合区氨基酸差异较大,在3种亚型结合域内均只有一个氨基酸相同。SLA-1*0703、SLA-3*0401和SLA-3*0403基因与人HLAI类基因KIR结合区氨基酸同源性介于SLA-2*11wz01和SLA-1*0401wz01之间。
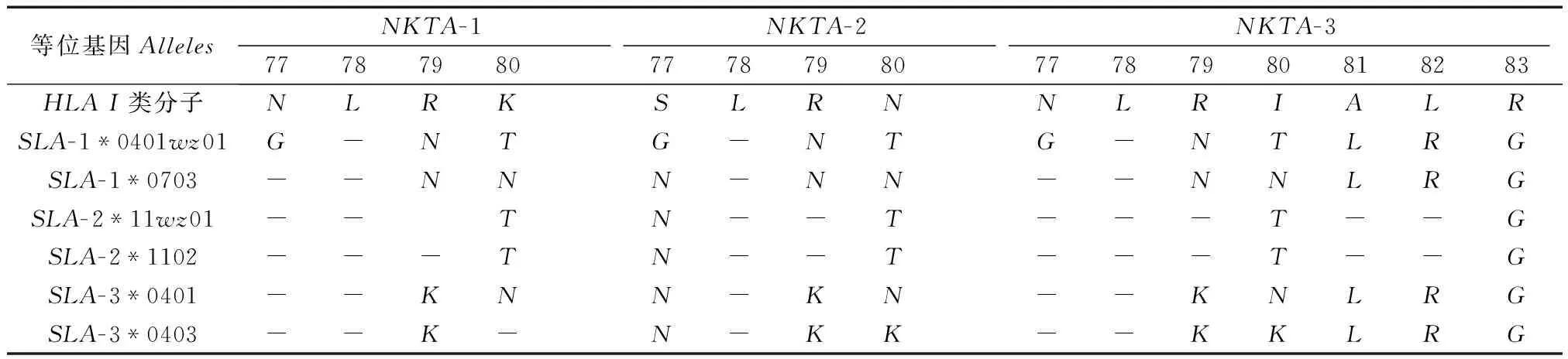
表5 SLA I类分子与HLA I类分子中KIR的配体氨基酸序列比较
注:NKTA-1/2/3表示3种与KIR结合的HLA氨基酸残基序列;“-”表示与人的HLA氨基酸序列相同。
Note.NKTA-1,NKTA-2andNKTA-3standforthreekindsofHLAaminoacidresidueswhichbindingtoKIRs. “-”indicatesthattheaminoacidresiduesareidenticaltothatinhumanHLAclassI.
3 讨论
SLAI类分子是引起细胞性排斥反应的核心因素和器官移植配型的重要位点[17],因此,明确猪SLA基因分子结构特征及与HLA同源性对异种器官移植配型具有重要的意义。SLA基因已知的NIH小型猪是目前国际异种器官移植研究常用的实验小型猪模型,且已被多次应用于猪-人血管性器官移植。本研究以NIH小型猪SLAI类基因分子序列为参考,比较GGTA1-/-五指山小型猪SLAI类基因与人HLA同源性高低。GGTA1-/-五指山小型猪SLA-1、SLA-3、SLA-2基因座上各等位基因与人HLA-A*0201同源性在70.5%~72.1%之间,与NIH小型猪相似性为83.2%~100%。五指山小型猪SLAI类分子与人HLA同源性范围和Chen等[18]对巴马、云南版纳猪和贵州香猪SLA-1、SLA-2基因分子与人HLA同源性相近。可见猪-人异种移植时,供受体之间SLA编码的膜蛋白不可能完全相同,只能选择SLA同源性较高的供受体进行异种移植。
已有研究表明HLA第223~228个氨基酸是与CD8+分子受体相互作用的关键结合域[18]。本研究SLA-1*0401wz01、SLA-1*0703、SLA-2*11wz01、SLA-2*1102和SLA-3*0401与人HLA-A*0201氨基酸序列比较发现,由苏氨酸(Thr)→丝氨酸(Ser)、苏氨酸(Thr)→蛋氨酸(Met),其他位点高度保守,这与Gao等[19]在巴马猪上研究报道一致。除此之外SLA-3*0403等位基因在位点225处由苏氨酸(Thr)→甘氨酸(Gly),在位点228处由苏氨酸(Thr)→蛋氨酸(Met),并在位点227~ 228之间插入4个氨基酸,这种多个氨基酸插入关键结合域有可能引起强烈的免疫排斥反应,即SLAI类分子本身作为外来抗原被人T细胞间接识别,这有待于对五指山猪SLA经典I类基因功能及与人的CD8+T细胞免疫位点进一步研究。
为解决因猪血管内皮细胞表达的SLAI类分子与HLAI类分子中KIR的配体序列不匹配,而引起的猪血管损伤问题,等Sasaki[20]将多态性较小且能和3种人KIR相匹配的HLA-G基因转入猪内皮细胞,体外功能实验表明,人NK细胞对猪内皮细胞的攻击明显减弱。本研究中SLA-2*11wz01和SLA-2*1102与人HLAI类基因KIR结合区仅有1(NKTA-1)或2(NKTA-2、NKTA-3)个氨基酸差异,SLA-2*11wz01和SLA-2*1102这种高度的同源性是否说明人NK细胞能明显降低对GGTA1-/-五指山小型猪血管内皮细胞的攻击,有待于进一步研究。
综上所述,从免疫细胞介导的异种排斥反应角度出发,GGTA1-/-五指山小型猪SLAI类基因氨基酸序列与人HLA高度相似,与人免疫细胞受体的配体关键结合域相似性较高,可作为未来猪-人异种器官移植的良好供体之一。
[1]CooperDK,EkserB,TectorAJ.Immunobiologicalbarrierstoxenotransplantation[J].IntJSurg, 2015, 23: 211-216.
[2]SchmockelM,NollertG,ShahmohammadiM,etal.Transgenichumandecayacceleratingfactormakesnormalpigfunctionasaconcordantspecies[J].JHeartLungTranspl, 1997, 169(7): 758-764.
[3]PhelpsCJ,KoikeC,VauqhtTD,etal.ProductionofAlpha1,3galactosyltransferase-deficientpigs[J].Science2003, 299(5605): 411- 414.
[4]郑道山, 冯冲, 朱彦宾, 等.利用启动子缺陷型打靶载体敲除五指山小型猪GGTA1基因 [J].生物技术通讯, 2011, 22: 458-462.
[5]DonnellyCE,YatkoC,JohnsonEW,etal.HumannaturalkillercellsaccountforMHCclassIrestrictedcytolysisofporcinecells[J].CellImmunol, 1997, 175:171-178.
[6]YamadaK,SachsDH,DerSimonianH.Humananti-porcinexenogeneicTcellresponse.Evidenceforallelicspecificityofmixedleukocytereactionandforbothdirectandindirectpathwayofrecognition[J].JImmunol, 1995,155: 5249-5256.
[7]BraveryCA,BattenP,YacoubMH,etal.DirectrecognitionofSLA-andHLA-likeclassIIantigensonporcineendotheliumbyhumanTcellsresultsinTcellactivationandreleaseofinterleukin-2 [J].Transplatation,1995, 60: 1024-1032.
[8]XieJ,ChenFX,LiNL,etal.NovelSLAclassIallelesofChinesepigstrainsandtheirsignificanceinxenotransplantation[J].JImmunol,2000,20: 136-139.
[9]VaimanM,RenardC,LafageP,etal.Evidenceforahistocompatibilitysysteminswine(SLA-A) [J].Transplatation,1970,10: 155-164.
[10]LeBas-BernardetS,BlanchoG.Currentcellularimmunologicalhurdlesinpig-to-primatexenotransplantation[J].TransplImmunol, 2009, 21(2): 60-66.
[11]KitchensWH,UeharaS,ChaseCM,etal.Thechangingroleofnaturalkillercellsinsolidorganrejectionandtolerance[J].Transplantation, 2006,81(6): 811-817.
[12]孙俊丽,牟玉莲,刘小林,等. 五指山小型猪近郊系白细胞抗原I类3基因的研究 [J].中国农业科学, 2007,40(9): 2053-2061.
[13]顾茂松,赵书红.五指山小型猪SLA-DQA基因的SNPs及猪人MHCI类区自然杀伤细胞KIRs结合区氨基酸变异的比较分析 [D].硕士学位论,华中农业大学,pp. 24-31.
[14]吴群,熊平,陈实,等.近交系海南五指山猪SLA经典I类和II类分子序列分析 [J].现代医学免疫, 2004,24: 23-26.
[15]窦科峰,李霄,陶开山,等.异种肝移植现状与未来 [J].中国实用外科杂志, 2014, 34: 14-18.
[16]JiHC,LiX,YueSQ,etal.PigBMSCstransfectedwithhumanTFPIcombatspeciesincompatibilityandregulatethehumanTFpathwayinvitroandinarodentmodel[J].CellPhysiolBiochem,2015,36: 233-249.
[17]ChadonP,RenardC,VaimanM.Themajorhistocompatibilitycomplexinswine[J].ImmunolRev, 1999, 167: 179-192.
[18]ChenFX,TangJ,LiNL,etal.NovelSLAclassIallelesofChinesepigstrainsandtheirsignificanceinxenotransplantation[J].CellRes,2003,13: 285-294.
[19]GaoCX,JiangQ,GuoD,etal.Characterizationofswineleukocyteantigen(SLA)polymorphismbysequence-basedandPCR-SSPmethodsinChineseBamaminiaturepigs[J].DevCompImmunol, 2014,45: 87-96.
[20]SasakiH,XuXC,SmithDM,etal.HLA-Gexpressionprotectsporcineendothelialcellsagainstnaturalkillercellmediatedxenogeneiccytotoxicity[J].Transplantation,1999, 67:31-37.
Characterization of swine leukocyte antigen class I genes and homologyanalysisofthesimilaritytoHLAinGGTA1-/-Wuzhishanminipigs
JIANGYing-di1,2,ZENGGuo-min2,3,SHINing-ning2,LIXi-rui2,JIHui-li2,PANDeng-ke2*,GUNShuang-bao1*
(1.CollegeofAnimalScienceandTechnology,GansuAgriculturalUniversity,Lanzhou730070,China;2.KeyLaboratoryofFarmAnimalGeneticResourceandGermplasmInnovationofMinistryofAgriculture,InstituteofAnimalScience,ChineseAcademyofAgriculturalSciences,Beijing100193;3.CollegeoflifeScienceandTechnology,GansuAgriculturalUniversity,Lanzhou730070)
ObjectiveThisstudywasaimedtocharacterizetheswineleukocyteantigen(SLA)classIgenesofGGTA1-/-Wuzhishanminipigsandcomparetheirsimilaritytohumanleukocyteantigen(HLA).Ithasimportantimplicationsforunderstandingthecellularrejectioninxenotransplantation.MethodsSpecimensofeartissuefromsixfoundingGGTA1-/-Wuzhishanminipigswerecollected,andtheSLAclassIgenes(SLA-1,SLA-3,SLA-2)wereamplifiedbyRT-PCR.PurifiedproductswereclonedintopEASY-T1vectorsandsequenced,followedbyBLASTalignmentandusingbioinformatcanalysistocharacterizetheSLAclassIgenesandcomparewiththesimilaritytoHLA.ResultsAtotalofsixallelesweredetected,amongthemalleleswerepreviouslyreported(SLA-1*0703,SLA-2*1102,SLA-3*0401,SLA-3*0403),andtheotherwerenovel(SLA-1*0401wz01,SLA-2*11wz01).ThehomologybetweenallelesofSLAclassIgenesinWuzhishanminipigsandHLAwasfrom70.5%to72.1%.ThehomologyanalysisofcriticalaminoacidresiduesonHLAbindingwithhumanCD8+moleculesshowedthatSLA-1*0401wz01,SLA-1*0703,SLA-2*11wz01,SLA-2*1102andSLA-3*0401occurredmutantataminoacidpositions225and228 (T→S,T→M),whereastheotherlociwerehighlyconserved.TherewasahighhomologyataminoacidlevelbetweenSLA-2*11wz01,SLA-2*1102andHLAclassIgeneswhichareNKcellKIRsbindingsites.ConclusionsTheaminoacidsequencesofSLAclassIgenesofGGTA1-/-WuzhishanminipigshaveahighhomologytoHLA.Fromthepointofviewofcell-mediatedxenograftrejection,theaminoacidsequencesofSLAclassIgenesofGGTA1-/-WuzhishanminipigshaveahighhomologytoHLA,therefore,Wzhishanminipigsmaybecomeagoodpotentialdonorforpig-humanxenotransplantation.
GGTA1-/-;Wuzhishanminipigs;Swineleukocyteantigen,SLA;Xenotransplantation;Immunerejection
GUNShuang-bao,E-mail:gunsb@gsau.edu.cn;PANDeng-ke,Email:pandengke2002@163.com
国家重点基础研究发展计划(编号:2015CB554103)。
蒋应弟(1986-),女,硕士研究生,研究方向:动物遗传育种与繁殖。E-mail: 529149896@qq.com
滚双宝(1967-),男,博士,教授,主要从事动物遗传育种。E-mail:gunsb@gsau.edu.cn
潘登科(1972-),男,博士,副研究员,主要从事转基因猪模型。E-mail:pandengke2002@163.com
研究报告
Q95-33
A
1005-4847(2016)04-0375-06
10.3969/j.issn.1005-4847.2016.04.008
2016-01-18
