成年心肌梗死小鼠心肌细胞去分化现象观察
2016-09-05沈文艳陈瑶李军
沈文艳,陈瑶,李军
( 1上海交通大学医学院附属仁济医院,上海200217;2上海交通大学Med-X 临床干细胞研究中心)
成年心肌梗死小鼠心肌细胞去分化现象观察
沈文艳1,2,陈瑶2,李军2
( 1上海交通大学医学院附属仁济医院,上海200217;2上海交通大学Med-X 临床干细胞研究中心)
目的观察成年心肌梗死小鼠心肌细胞是否存在去分化现象。方法20只成年小鼠随机分为观察组和对照组各10只,观察组制备心肌梗死模型,对照组仅行假手术处理。采用RT-PCR法检测两组心肌组织 α-横纹肌肌动蛋白(α-SA)、横纹肌蛋白(α-actinin)、转录因子nkx2.5、肌细胞增强因子2(mef2)、细胞增殖核抗原(Ki67)、干细胞标记物血管内皮生长因子受体(flk-1) mRNA,采用免疫荧光染色法检测两组心肌组织磷酸化组蛋白(PH3)、连接蛋白43(cx43)。结果观察组心肌组织α-SA、α-actinin、nkx2.5、mef2 mRNA表达量均低于对照组,Ki67、flk-1、mRNA表达量及PH3、cx43均高于对照组,P均<0.05, 结论成年心肌梗死小鼠心肌细胞存在去分化现象。
心肌梗死;心肌细胞;细胞去分化;α-横纹肌肌动蛋白;横纹肌蛋白;转录因子;细胞增殖核抗原;磷酸化组蛋白
急性心肌梗死(AMI)是冠状动脉急性、持续性缺血缺氧所引起的心肌坏死,是引起充血性心力衰竭的重要原因[1]。心肌梗死发生后,梗死区域的心肌细胞因为缺血损伤而失去了功能,心脏的泵血功能严重受损[2, 3]。目前常用治疗心肌梗死的方法一般是药物和介入治疗,这些治疗方法对已梗死的心肌无法修复。传统上认为,哺乳动物的心脏是一个终末分化器官[4],而心肌细胞是一种终末分化的细胞,心肌梗死后心肌细胞没有再生能力而导致心力衰竭的发生[5]。然而,最近越来越多的研究显示新生小鼠及新生儿心肌均具有再生能力[6~8]。也有研究显示,成体动物的心肌细胞可能也具有类似的能力[9]。连接蛋白43(cx-43)、 α-横纹肌肌动蛋白(α-SA)、横纹肌蛋白(α-actinin)、转录因子nkx2.5、肌细胞增强因子2(mef2)是常用的心肌细胞标记物,细胞增殖核抗原(Ki67)和磷酸化组蛋白(PH3)是常用的细胞增殖标记物,血管内皮生长因子受体(flk-1)是常用的干细胞标记物。我们在前期研究中发现,心肌梗死可以诱导小鼠心肌细胞在体外发生类似去分化的现象,但是体内的研究目前尚未见报道。2013年5月~2014年3月,我们观察了心肌梗死是否可诱导小鼠成体心肌细胞发生去分化现象。现报告如下。
[10] Bayat N, Ebrahimi-Barough S, Ardakan MM, et al. Differentiation of human endometrial stem cells into schwann cells in fibrin hydrogel as 3D culture[J]. Mol Neuro, 2015,6(4):1-7.
[11] Fan L, Yu Z, Li J, et al. Schwann-like cells seeded in acellular nerve grafts improve nerve regeneration[J]. BMC Mus Dis, 2014,15(6):165.
[12] Miura K, Okada Y, Aoi T, et al. Variation in the safety of induced pluripotent stem cell lines[J]. Nat Biotech, 2009,27(8):743-745.
[13] Xu H, Wang Y, He Z, et al. Direct conversion of mouse fibroblasts to GABAergic neurons with combined medium without the introduction of transcription factors or miRNAs[J]. Cell Cycle, 2015,14(15):2451-2460.
[14] Hoveizi E, Ebrahimi-Barough S, Tavakol S, et al. In Vitro differentiation of human iPS cells into neural iike cells on a biomimetic polyurea[J]. Mol Neuro, 2016,7(12):1-7.
[15] Liu Z, Tang Y, Lü S, et al. The tumourigenicity of iPS cells and their differentiated derivates[J]. J Cell Mol Med, 2013,17(6):782-791.
[16] Hu W, Qiu B, Guan W, et al. Direct conversion of normal and alzheimer′s disease human fibroblasts into neuronal cells by small molecules[J]. Cell stem cell, 2015,17(2):204-212.
[17] Li M, Zhang P, Li H, et al. TGF-β1 is critical for wallerian degeneration after rat sciatic nerve injury[J]. Neur Sci, 2015(284):759-767.
[18] Einheber S, Hannocks MJ, Metz CN,et al.Transforming growth factor-beta 1 regulates axon/Schwann cell interactions[J]. Jour Cell Bio, 1995,129(2):443-458.
[19] Leung AW, Kent Morest D, Li JY. Differential BMP signaling controls formation and differentiation of multipotent preplacodal ectoderm progenitors from human embryonic stem cells[J]. Develop Bio, 2013,379(2):208-220.
[20] Bonaguidi MA, Peng CY, McGuire T, et al. Noggin expands neural stem cells in the adult hippocampus[J]. J Neuro Sci, 2008,28(37):9194-9204.
[21] Monje PV. To myelinate or not to myelinate: fine tuning cAMP signaling in schwann cells to balance cell proliferation and differentiation[J]. Neu Regen Res, 2015,10(12):1936-1937.
[22] Latasa MJ, Cosgaya JM. Regulation of retinoid receptors by retinoic acid and axonal contact in schwann cells[J]. PLoS One,2011,6(2):17023.
[23] Zhang L, Ma Z, Smith GM, et al. GDNF-enhanced axonal regeneration and myelination following spinal cord injury is mediated by primary effects on neurons[J]. Glia, 2009,57(11):1178-1191.
1 材料与方法
1.1材料6~8周龄C57BL/6 SPF级雄性小鼠20只(体质量22~25 g),购自上海斯莱克模型动物有限公司;包埋剂OCT购自Sakura公司,1%SDS购自Sigma公司,Hanks平衡盐溶液购自法国Biowest公司,脱氧核糖核酸酶Ⅰ(DNase Ⅰ)购自Roche公司;鼠源抗小鼠细胞Ki67抗体购自美国BD公司,兔源抗小鼠PH3抗体购自美国Cell Signaling 公司,鼠源抗小鼠α-SA抗体购自英国Abcam公司,兔抗小鼠cx43抗体购自美国CST公司;细胞核荧光染料 4,6-二脒基-2-苯基吲哚(DAPI)购自美国Sigma公司,驴源抗兔Alexa Fluor488荧光染料,驴源抗兔Alexa Fluor555荧光染料均购自美国Invitrogen公司。引物由上海生工生物合成有限公司合成。
1.2小鼠心肌梗死模型制备将20只小鼠随机分为观察组和对照组各10只。观察组用异氟烷预麻箱麻醉后打开胸腔,于左心耳下缘约2 mm处结扎冠状动脉左前降支,制备心肌梗死模型。以小鼠冠状动脉左前降支供血区域心肌颜色变白[10]视为造模成功,最终造模成功10只。对照组仅麻醉后打开胸腔,不结扎血管行假手术处理。
1.3心肌组织α-SA、α-actinin、nkx2.5、mef2、Ki67、flk-1 mRNA表达检测采用RT-PCR法。造模7 d取两组小鼠,断头处死后取心肌组织;按照试剂盒说明书提取出总RNA,逆转录合成cDNA后进行扩增,以β-actin为内参。引物序列:β-actin上游引物5′-GGCTGTATTCCCCTCCATCG-3′,下游引物5′-CCAGTTGGTAACAATGCCATGT-3′,引物长度154 bp;Ki67上游引物5′-CCTGCCCGACCCTACAAAAT-3′,下游引物5′-TCCGCCGTCTTAAGGTAGGA-3′,引物长度313 bp;flk-1上游引物5′-AGAGTCAGACAACAACAATTGGCGAGAC-3',下游引物5′-GCATCATAAGGCAAGCGTTCACAGC-3′,引物长度451 bp;Nkx2.5上游引物5′-CAATGCCTATGGCTACAACGC-3′,下游引物5′-GAGTCATCGCCCTTCTCCTAAA-3′,引物长度297 bp;α-SA上游引物5′-CAGCCCTCTTTCATTGGT-3′,下游引物5′-CTGCCTCATCATACTCTT-3′,引物长度310 bp;mef上游引物5′-AAGTACACCGAGTACAACG-3′,下游引物5′-TAACTGGCATCTCAAAGC-3′,引物长度235 bp;α-actinin上游引物5′-GACCCAGATCATGTTTGAGACCTTC-3′,下游引物5′-CATTATGAGTTACACCATCGCCAGA-3′,引物长度130 bp。 反应体系 30 μL, 扩增条件:72 ℃、45 s延伸, 95 ℃ 、30 s,55 ℃、30 s,30个循环,72 ℃、10 min,4 ℃冷却,进行琼脂糖凝胶电泳。用Image J软件分析目的条带灰度,以其与β-actin的灰度比值计算目的基因相对表达量。实验重复3次。
1.4心肌组织PH3、cx43表达检测采用免疫荧光染色法。取造模7 d时两组小鼠心肌组织, OCT包埋切片,驴血清封闭;用1%BSA稀释抗体(1∶100)一抗孵育,4 ℃ 过夜。次日37 ℃复温45 min,二抗孵育后洗涤。DAPI复染,封片,采用荧光显微镜观察心肌组织PH3、cx43染色结果。用Image J软件分别测免疫荧光结果中阳性荧光面积比上该视野的面积,计算相对表达量。实验重复3次。
2 结果
2.1两组心肌组织α-SA、α-actinin、nkx2.5、mef2、Ki67、flk-1 mRNA表达比较见表1。
2.2两组心肌组织PH3、cx43表达比较见表2。
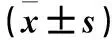
表1 造模7 d两组心肌组织α-SA、α-actinin、nkx2.5、mef2、Ki67、flk-1 mRNA相对表达量比较
注:与对照组比较,#P<0.05。
3 讨论
长期以来,人们认为心肌细胞是终末分化细胞,自身没有再生能力。有研究显示,心肌细胞可以被诱导再分裂,使心肌细胞去分化,重新进入细胞周期[11]。新生的心肌细胞也可以通过去分化的方式参与心脏自稳态的维护和病变组织的修复[12]。研究发现,成年小鼠心肌细胞可在急性损伤后发生去分化现象,以保证在存活基础上在特定条件下进一步再生为心肌细胞。
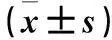
表2 造模7 d两组心肌组织PH3、cx43相对表达量比较
注:与对照组比较,#P<0.05。
本研究发现,造模7 d观察组心肌组织cx-43、α-SA及α-actinin mRNA相对表达量均低于对照组,提示心肌肌节解聚现象明显,心肌细胞横纹肌结构破坏。 Nkx2.5和mef2是两个与心脏发育密切相关的转录因子。Nkx2.5参与了心脏形成,包括右侧环化、心腔分化、心脏间隔功能成熟以及工作心肌和传导系统的维持等过程[13]。心肌梗死后nkx2.5和mef2的表达减少,提示心肌梗死后成熟心肌细胞结构丧失,退回到未成熟心肌细胞或前体细胞状态。我们前期研究显示,心肌梗死后小鼠的心肌细胞有Ki67表达的增加,并与乳鼠心肌组织表达接近,进一步说明心肌梗死后部分心肌细胞的增殖能力可能退回到接近乳鼠未成熟心肌细胞或前体细胞的阶段。在正常成体组织中,绝大部分细胞处于非增殖状态,细胞学上称为G0期。要构建良好的组织结构并执行特定的功能,必须依靠这些增殖周期外的G0期细胞。Ki67免疫组化染色可将大部分G0期以外的增殖细胞标记,因而也被称为细胞的增殖指数。Ki67阳性率越高,说明处于增殖周期的细胞比例越高,组织分化越好。PH3只出现于有丝分裂期细胞染色体,静止期细胞不表达,PH3表达的增高可以更准确地提示心肌梗死诱导成体心肌细胞增殖[14]。本研究发现,观察组心肌组织存在Ki67和PH3高表达,与以往研究结果一致。flk-1是启动胚胎造血发育的必需基因。Shalaby等[15]报道,flk-1基因能调节早期中胚层来源的间质细胞迁移至造血微环境;Miyagi等[16]发现小鼠胚胎干细胞体外发育至表达flk-1阶段已具备重建造血功能,flk-1基因敲除的胚胎干细胞不能发育为造血细胞。本研究中,观察组心肌组织中flk-1相对表达量明显高于对照组,进一步提示心肌梗死后心肌细胞退回至未成熟心肌细胞或前体细胞阶段。
综上所述,成年心肌梗死小鼠心肌细胞存在去分化现象,但其具体作用机制尚有待于进一步研究。
[1] Guo J. Influence of dietary patterns on the risk of acute myocardial infarction in China population: the INTERHEART China study[J]. Chin Med J, 2013,126(3):464-470.
[2] Bearzi C. Human cardiac stem cells[J]. Proc Natl Acad Sci USA, 2007,104(35):14068-14073.
[3] Altarche W,Xifro F. Cardiac c-kit+AT2+cell population is increased in response to ischemic injury and supports cardiomyocyte performance[J]. Stem Cells, 2009,27(10): 2488-2497.
[4] Hosoda T. C-kit-positive cardiac stem cells and myocardial regeneration[J]. Am J Card Dis, 2012, 2(1): 58-67.
[5] Zaruba MM. Cardiomyogenic potential of C-kit(+)-expressing cells derived from neonatal and adult mouse hearts[J]. Circulation, 2010,121(18): 1992-2000.
[6] Porrello ER, Mahmoun Al, Simpson E, et al. Transient regenerative potential of the neonatal mouse heart[J]. Science, 2011,331(6020):1078-1080.
[7] Rücker MC, Pecker F, Godreau D, et al. Dedifferentiation of atrial myocytes during atrial fibrillation:role of fibroblast proliferation in vitro[J]. Cardiovasc Res, 2002,55(1):38-52.
[8] Polizzotti BD, Ganapathy B, Walsh S, et al. Neuregulin stimulation of cardiomyocyte regeneration in mice and human myocardium reveals a therapeutic window[J]. Sci Transl Med, 2015,7(281):231-245.
[9] Zhang Y, Li TS, Lee ST, et al. Dedifferentiation and proliferation of mammalian cardiomyocytes[J]. PLoS One, 2010,5(9):12559.
[10] Breitbach M, Bostani T, Roell W, et al. Potential risks of bone marrow cell transplantation into infarcted hearts [J]. Blood,2007,110(4):1362-1369.
[11] Engel FB, Schebest M, Duong MT, et al. P38 MAP kinase inhibition enables proliferation of adult mammalian cardiomyocytes[J]. Genes Dev, 2005,19(10):1175-1187.
[12] Beltrami AP,Urdanek K,Kajstura J,et al.Evidence that human cardiac myocytes divide after myocarction[J]. N Engl J Med,2001(344):1750-1757.
[13] Matsuura K. Cardiomyocytes fuse with surrounding noncardiomyocytes and reenter the cell cycle[J]. J Cell Biol, 2004,167(2):351-363.
[14] Engel FB,Schebest M,Duong MT,et al. P38 MAP kinase inhibition enables proliferation of adult mammalian cardiomyocytes[J]. Genes Dev, 2005,19(10):1175-1187.
[15] Shalaby F, Ho J, Stanford WL, et al. A requirement for Flk-1 in primitive and definitive hematopoiesis and vasculogenesis[J]. Cell, 1997,89(6);981-990.
[16] Miyagi T, Takeno M, Nagafuchi H, et al. FLK1+cells derived from mouse emhryonic stem cells recruiting VEGFR1+stem cells from bone-marrow microenvironment[J]. Nature Med, 2002,18(8):841-849.
(收稿日期:2016-01-15)
国家自然科学基金资助项目(81300088)。
李军(E-mail: jun.li@sjtu.edu.cn)
10.3969/j.issn.1002-266X.2016.18.011
R544.22
A
1002-266X(2016)18-0034-03
2016-01-20)
