混交对红松人工林细根生物量和空间分布的影响1)
2016-08-18钱文丽卢元王韶仲谷加存
钱文丽 卢元 王韶仲 谷加存
(东北林业大学,哈尔滨,150040) (辽宁省林业调查规划院) (东北林业大学)
混交对红松人工林细根生物量和空间分布的影响1)
钱文丽卢元王韶仲谷加存
(东北林业大学,哈尔滨,150040)(辽宁省林业调查规划院)(东北林业大学)
以黑龙江省尚志市帽儿山实验林场的红松(Pinuskoraiensis)人工纯林、红松×胡桃楸(Juglansmandshurica)和红松×水曲柳(Fraxinusmandschurica)混交林为研究对象,采用土钻法估计并比较了不同混交处理下红松、伴生树种和其他木本植物细根(直径≤2.0 mm)生物量及其垂直分布特征。结果表明:细根总生物量(0~30 cm)以纯林最高(212.6 g·m-2),红松×胡桃楸混交林最低(164.7 g·m-2),而红松×水曲柳混交林居中(200.8 g·m-2);纯林中红松细根生物量显著高于红松×胡桃楸混交林,但与红松×水曲柳混交林没有显著差异;混交没有引起林分细根总生物量垂直分布的改变,均为随着土壤深度增加而减少;混交后红松细根生物量在土壤表层(0~10 cm)分配的比例增加,但是吸收根(直径≤0.5 mm)生物量在土壤底层(20~30 cm)分配的比例在红松×胡桃楸混交林(64.5%)中比纯林高(40.1%),而在红松×水曲柳混交林中所占比例较低(22.0%)。方差分析显示,混交树种、土壤深度和根直径等级均是影响红松细根生物量的重要因子。这些结果表明,混交对红松人工林细根生物量及其空间分布有明显影响,这为深入认识红松及其混交树种间的相互作用提供了必要的理论依据。
混交林;红松;水曲柳;胡桃楸;细根
We used soil core method to determine how species mixing betweenPinuskoraiensis,JuglansmandshuricaandFraxinusmandschuricaaffects the fine root (diameter ≤2.0 mm) biomass and spatial distribution inP.koraiensisplantation. Roots were categorized into three groups: root ofP.koraiensis, root of component tree species, and root of other woody plants. Total fine root biomass per unit area was the highest (212.6 g·m-2) in pureP.koraiensisplantation, the lowest (164.7 g·m-2) inP.koraiensis×J.mandshuricaplantation, and medium (200.8 g·m-2) inP.koraiensis×F.mandschuricaplantation. Root biomass ofP.koraiensiswas significantly higher in pure stand than that ofP.koraiensis×J.mandshuricaplantation, while was similar to that ofP.koraiensis×F.mandschuricaplantation. Species mixing did not change the vertical distribution of total fine root biomass, with a tendency of decline associated with soil depth increasing. The proportion of fine roots ofP.koraiensisat the surface layer (0 to 10 cm) was higher in mixed-species forest plantations than in pure plantation, however, the proportion of absorptive roots at the deepest soil layer (20-30 cm) was the highest (64.5%) inP.koraiensis×J.mandshuricaplantation, following by pure (40.1%) andP.koraiensis×F.mandschurica(22.0%) plantations. Factorial ANOVA indicated that species mixing, soil depth and root diameter classes all significantly contributed to the variation of root biomass. Therefore, the species mixing had significant effect on fine root biomass and vertical distribution ofP.koraiensis, and provide some insights into the belowground ecological interaction betweenP.koraiensisand its component species.
细根(直径≤2.0 mm)具有巨大的吸收表面积和较强的生理活性,是林木水分和养分吸收的主要器官[1],并在森林生态系统的物质循环与能量流动上发挥了重要作用[2]。随着森林生态系统及全球碳循环研究的深入,细根及其相关研究已成为国内外生态学研究的热点问题[3]。树种混交作为重要的森林经营措施之一,不仅影响林分干材的生长过程,而且对细根动态和分布特征均有影响。一些研究表明,混交会促进主要树种细根生长,如BRASSARD et al.[3]对北美寒带森林的研究显示,美洲颤杨(Populustremuloides)和胶冷杉(Abiesbalsamea)、北美云杉(Piceaspp.)混交林的细根生物量和生产力均高于美洲颤杨纯林。WANG et al.[4]对异叶铁杉(Tsugaheterophylla)、摺扇崖柏(Thujaplicata)混交林的研究中也得到相同变化规律。然而也有研究发现,混交对主要树种细根生物量没有影响,甚至使之降低。如LEUSCHNER et al.[5]的研究表明欧洲山毛榉(Fagussylvatica)和岩生栎(Quercuspetraea)混交林细根生物量与纯林相比没有显著变化,而BAUHUS et al.[6]研究显示蓝桉(Eucalyptusglobulus)和黑荆(Acaciamearnsii)混交林比黑荆纯林活细根生物量降低了12.25%。以往研究结果的差异可能与林分组成、林龄、立地质量等的不同有关。因此,针对不同树种或在不同立地条件下开展混交对细根生长与动态的研究,对于全面认识混交林细根变异规律和作用机制具有重要意义。
红松(Pinuskoraiensis)是我国东北地区主要的造林树种之一,也是我国重要的用材树种。以往研究关注了混交对红松阔叶林林分空间结构的影响[7-9]、林分抚育对红松生产力和生物量的影响[10-11]等方面,但多是集中于林木地上部分,针对混交林地下细根生物量及其动态的研究开展较少[12-15]。本文以黑龙江省尚志市帽儿山地区红松人工纯林、红松×胡桃楸(Juglansmandshurica)和红松×水曲柳(Fraxinusmandschurica)混交林为研究对象。通过土钻法估计并比较不同混交处理下不同直径等级(≤0.5 mm,>0.5~1.0 mm,>1.0~2.0 mm)细根生物量及其垂直分布特征。主要目的是比较树种混交对红松人工林细根生物量和垂直分布的影响,明确影响混交林细根(特别是红松细根)生物量的主要因子。研究结果将为全面认识红松及其混交树种间的相互作用提供必要的理论依据。
1 研究区概况
研究地点位于黑龙江省尚志市帽儿山实验林场尖砬沟森林培育实验站(127°30′~127°34′E,45°21′~45°25′N),该地区属温带大陆性季风气候,平均海拔300 m,坡度10°~15°,属于长白山系张广才岭西北坡。年平均气温2.8 ℃,年平均降水量723 mm,年平均蒸发量1 094 mm,无霜期120~140 d。研究样地设于山坡中部,土壤为暗棕壤,土层厚度在30~40 cm。人工林营造于1986年,红松纯林为10行,红松混交林配置方式均为5行红松、3行阔叶树,各林分株行距均为1.5 m×2.0 m。在林分踏查的基础上,于2013年9月末,在红松纯林、红松×胡桃楸和红松×水曲柳混交林(简称为处理A、B和C)内随机设置样地3块重复样地(长度20 m,宽度为纯林或混交林带总宽度16~20 m),进行每木检尺并调查土壤状况,林分和土壤特征见表1。
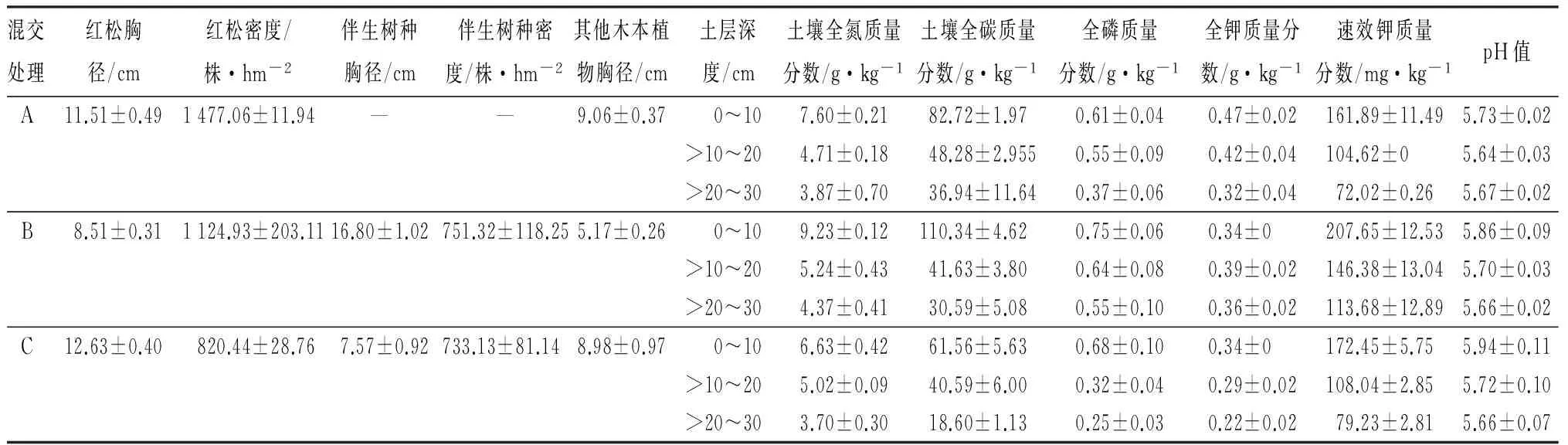
表1 红松人工纯林和混交林林分与土壤特征
注:A为红松纯林,B为红松×胡桃楸,C为红松×水曲柳;表中数值为平均值±标准误差;—表示数据不符合分析要求或没有数据。
2 研究方法
2.1样地设置
2013年9月末,在每块重复样地(共9块)内随机选取5个样点(混交林中在红松和伴生树种林带交界均取样),清除地表凋落物,通过土钻法(土钻内径58 mm)钻取土芯,取样深度设为30 cm,分为3层(0~10 cm为表层,>10~20 cm为亚表层,>20~30 cm为底层),合计取样数为135个。样品装入已编号的封口袋,4 h内带回实验站处理。用2 mm筛筛出各约100 g土芯部分鲜土,并手工将土壤中的根系个体挑出,风干后依据相应国家标准[16],进行理化性质分析。其余含有根系的土芯放入小桶内充分浸泡(约2 h),以便根系与土壤分离,在流水中过40目筛网,去除杂质及土壤颗粒,洗净后将根系低温冷冻保存。根据根系形态特征和分支结构,剔除草本植物根系,进一步根据外形、颜色、弹性等将细根分为红松、伴生树种(水曲柳和胡桃楸)、其他木本植物根系(主要树种以外的乔木和灌木)3类,并区分根系死活状态。区分类别的基础上,将所有活根按直径≤0.5 mm、0.5~1.0 mm和1.0~2.0 mm分级。将所有细根样品置于65 ℃下烘干至恒质量并称质量(精确度=0.000 1 g)。
2.2数据处理
数据计算和处理以单个土芯内单位面积的细根生物量为基础。在区分根系类别(红松、伴生树种和其他木本植物根系)的基础上,分别计算各样点内对应土壤深度(0~10 cm,>10~20 cm,>20~30 cm)和直径等级(≤0.5 mm、>0.5~1.0 mm和>1.0~2.0 mm)的根系生物量,整个土壤剖面(0~30 cm)则以相同直径等级根系指标加和计算。在单因素方差分析基础上,进行多重比较(LSD法),比较混交处理对不同土壤深度及整个土壤剖面(0~30 cm)红松、伴生树种和其他木本植物各直径等级细根生物量的影响;采用三因素方差分析方法,检验混交、土壤深度和直径等级对红松、伴生树种和其他木本植物细根生物量的影响,显著性水平设为P<0.05。所有数据统计分析均采用SPSS for Windows 19.0(2011年,SPSS Inc.,USA)完成。
3 结果与分析
3.1不同混交处理各径级细根生物量比较
不同混交处理细根(直径≤2.0 mm)生物量(0~30 cm土层之和)之间存在差异,处理A(红松纯林)、B(红松×胡桃楸混交林)和C(红松×水曲柳混交林)细根总生物量分别为212.6、164.7和200.8 g·m-2,其中红松细根生物量分别为73.4、10.0和67.2 g·m-2,其他木本植物细根生物量分别为139.1、65.5和98.2 g·m-2,混交林处理B、C伴生树种细根生物量分别为89.1(胡桃楸)和35.4 g·m-2(水曲柳)。处理A中红松各直径等级细根生物量与处理C没有显著差异,但二者均显著高于处理B(表2)。在所有处理中,红松细根生物量均低于相同直径等级伴生树种和其他木本植物根系之和,特别是在处理B中(表2)。处理B中胡桃楸各直径等级细根生物量高于其他木本植物,而处理C中水曲柳细根生物量则低于对应的其他木本植物根系(表2)。
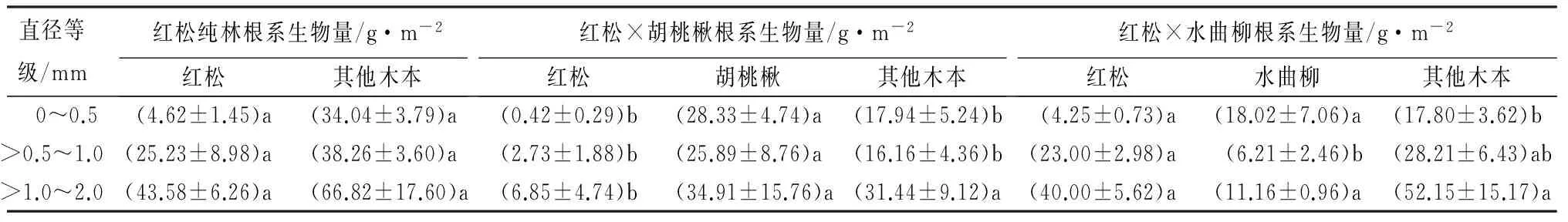
表2 不同混交处理下红松、伴生树种和其他木本植物根系生物量(0~30 cm土壤深度)
注:表中括号内数值为平均值±标准误差,同一行相同类群根系不同小写字母表示不同处理下根系生物量差异显著(P<0.05)。
3.2不同混交处理细根生物量垂直分布差异
混交并没有改变细根(直径≤2.0 mm)总生物量的垂直分布格局,在各处理中均表现为随着土壤深度增加而减少,处理A、B和C中土壤表层(0~10 cm)细根生物量占整个取样深度的比例分别为48.4%、49.6%和48.1%,红松、伴生树种和其他木本植物细根也主要集中在土壤表层0~10 cm(表3)。但是,混交后红松细根生物量在土壤表层分配的比例增加:纯林处理A中根系生物量在各层中占整个土壤剖面的比例分别为37.9%、33.6%、28.4%,处理B中比例分别为52.8%、22.9%、24.3%,处理C为42.5%、32.9%、24.6%。然而,红松不同直径等级细根生物量的垂直分布对混交处理的响应不同,直径0~0.5 mm的根群更敏感:处理A中该根群生物量在土壤表层至底层的分布比例分别为36.1%、23.8%和40.1%,处理B为21.7%、13.8%和64.5%,而处理C中为48.3%、29.7%和22.0%(表3)。但是,混交树种(胡桃楸和水曲柳)与其他木本植物直径0~0.5 mm的根群生物量均表现出随土层加深而减少的特征(表3)。
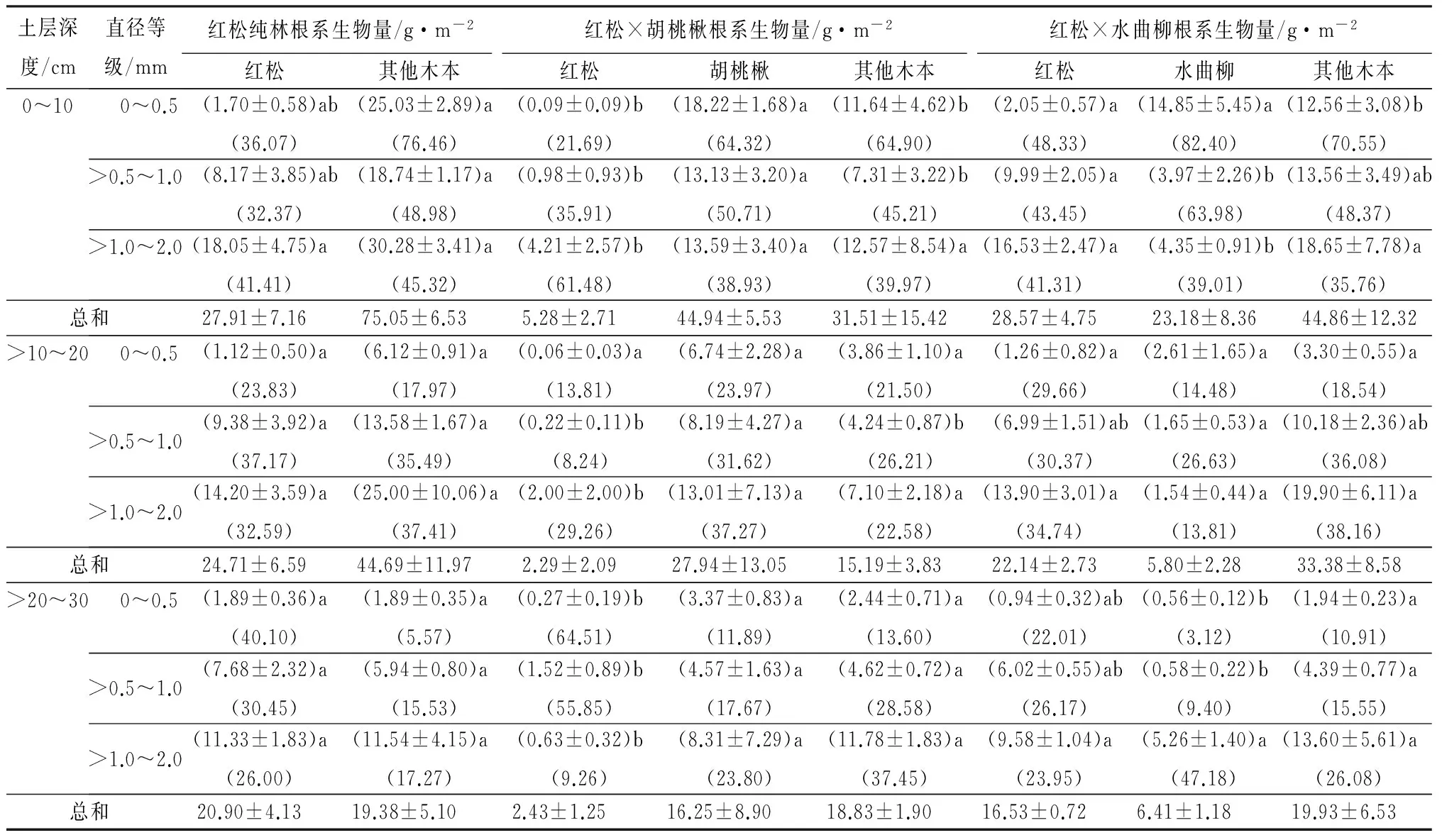
表3 不同混交处理下红松、伴生树种和其他木本植物根系生物量空间分布特征
注:表中第一个括号内数值为平均值±标准误差,第二个括号内数值为该类群根系生物量在某个土层占整个土壤剖面细根总生物量的百分比;同一行相同类群根系不同小写字母表示不同处理下生物量差异显著(P<0.05)。
3.3影响红松细根生物量及其垂直分布的因子分析
红松细根生物量显著受混交处理、土壤深度和直径等级的影响,但是不同混交处理下相同直径等级生物量的关系有所不同(表现为混交处理和直径等级的交互作用,表4)。混交林中,胡桃楸和水曲柳细根生物量存在显著的差异,但是直径等级的影响不显著;其他木本植物细根生物量与红松类似,受到混交处理、土壤深度和直径等级的显著影响,同时表现出土壤深度与混交处理的显著交互作用(表4)。
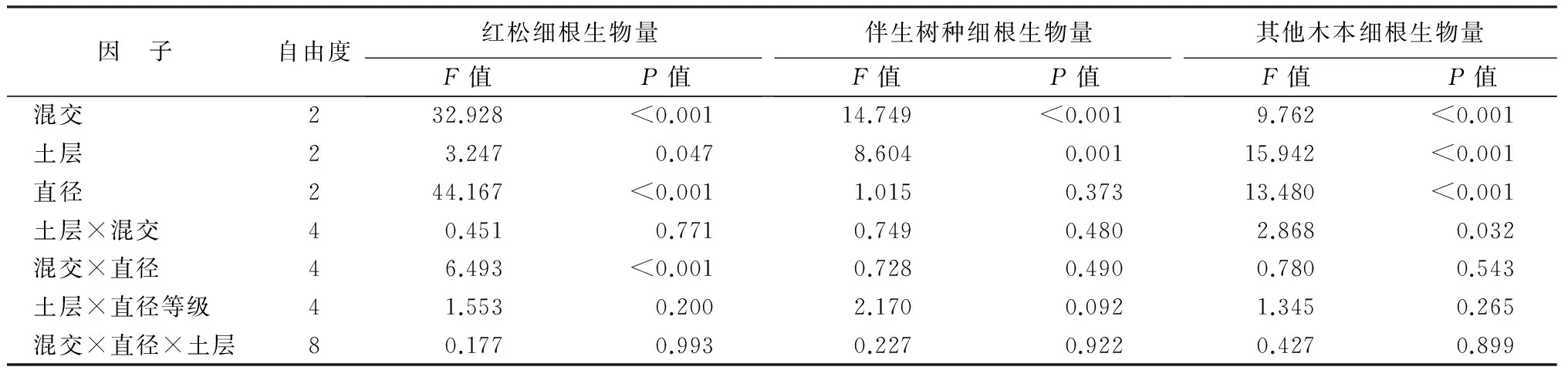
表4 混交、土壤深度(土层)和直径等级(直径)对细根生物量的影响
注:粗体数值表示统计上显著(P<0.05)。
4 讨论
4.1混交对红松细根生物量的影响
混交对林分细根总生物量和红松细根生物量均有影响(表2和表4)。当不考虑其他木本植物细根生物量时,混交林中红松及其伴生树种细根生物量之和均高于纯林中红松细根生物量(约1.3倍)。与此类似,SCHMID et al.[17]的研究表明挪威云杉(Piceaabies)与欧洲山毛榉(Fagussylvatica)混交后细根总生物量是云杉纯林的1.9倍;BRASSARD et al.[3]的研究也发现混交促进细根总生物量增加。这可能是因为根系生态位分离使混交林中目的树种和伴生树种根系对资源的获取和利用较纯林更充分[18],林分林木生产力更高,细根生物量增加。但是,需要指出的是,红松×胡桃楸混交林中细根总生物量的增加主要受胡桃楸根系的影响,而红松细根生物量显著低于纯林,这可能是胡桃楸抑制了红松的生长,表现为红松平均胸径和细根生物量均低于纯林(表1和表2)。当考虑其他木本植物细根生物量时,红松纯林中其根系生物量远高于混交林,因此导致红松纯林细根总生物量较高,但是与红松×水曲柳混交林差异不显著。红松×胡桃楸混交林具有较低的细根总生物量,一方面可能是因为胡桃楸林木个体生长上占优势(平均胸径高于红松及其他木本植物),因此在资源竞争上处于优势;另一方面,也可能与胡桃楸林木根系分泌物和凋落物分解过程中存在化感效应有关[19],但是化感作用的程度还需要进一步的研究来证实。
4.2混交对红松细根垂直分布的影响
由于森林土壤资源(特别是养分)有效性从表层至底层逐渐降低,导致树木根系垂直分布具明显的成层性。本研究中各林分细根总生物量均随土壤深度增加而减少,表层细根生物量占总生物量的近一半(表3),以往研究也报道了类似的结果。例如,陈硕芃等[20]对小兴安岭大青山红松人工林的研究显示,表层(0~10 cm)细根总生物量占0~30 cm土层的42.9%~53.5%;王树堂等[15]研究报道长白山阔叶红松林木本植物总细根生物量占整个土壤剖面的52.5%~60.0%。土壤表层丰富的养分、较高的温度,可能是导致细根在表层大量分布的重要原因[20],而混交并没有改变林分细根总生物量的垂直分布规律。
然而,混交后红松细根生物量的垂直分布规律有所改变。混交导致红松细根总生物量在表层分布的比例有所提高,由37.9%(纯林)增加至42.5%(红松×水曲柳)和52.8%(红松×胡桃楸)。类似地,其他研究也发现了混交后根系垂直分布改变的现象。与挪威云杉(Piceaabies)纯林相比,混交林中云杉根长密度在表层腐殖质层分配比例增加,且在此层具有较大的土壤N和P的吸收[21]。进一步分析发现,红松×胡桃楸混交林中表层根系生物量增加主要是由于非吸收根生物量(直径0.5~2.0 mm)增加引起,因为吸收根(直径≤0.5 mm)生物量主要分配至底层(64.5%);而红松×水曲柳混交林中表层红松细根生物量的增加则主要由吸收根与非吸收根的共同增加引起的(表3)。理论上,目的树种与混交树种在资源吸收上存在竞争关系,一方根系生物量的改变,将会引起另一方细根垂直分布的改变。我们发现,红松以外所有木本植物细根生物量(即伴生树种和其他木本之和)在表层分布的比例在红松×胡桃楸混交林(49.4%)和红松×水曲柳混交林(50.9%)中相差不大,但是均低于红松纯林(54.0%)。因此,综合来看,红松×胡桃楸混交林中红松细根生物量在表层分配比例较高,并非简单地因为其他树种(包括胡桃楸)根系的竞争在表层降低导致,但是具体原因还有待进一步研究。此外,红松×胡桃楸混交林中红松吸收根生物量在底层分配比例较大,表明红松根系的资源吸收在表层受到更强的抑制,从而通过在底层增加吸收根的分配来避免与其他树种在养分丰富的土壤表层展开竞争。总之,混交改变了红松细根生物量的垂直分布格局,但影响程度与混交树种有关,这可能受到混交树种根系形态结构、空间分布和生长动态等因素的影响[22]。
5 结论
通过比较红松人工纯林、红松×胡桃楸和红松×水曲柳混交林内红松、伴生树种和其他木本植物细根生物量及其垂直分布特征,结果表明林分细根总生物量(0~30 cm)和红松细根生物量在纯林与其他混交林分中存在差异。混交并没有引起林分细根总生物量垂直分布特征的改变,但导致红松细根生物量在土壤表层(0~10 cm)分配的比例增加。红松吸收根(直径≤0.5 mm)生物量在土壤底层(20~30 cm)分配的比例在不同林分中具有最大的变异性,显示出吸收根对资源竞争具有更高的敏感性。混交树种、土壤深度和根直径等级均是影响红松细根生物量的重要因子。这些结果增进了我们对红松及其混交树种间的相互作用及其潜在机制的认识。
[1]张小全,吴可红.森林细根生产和周转研究[J].林业科学,2001,37(3):126-138.
[2]NORBY R J, JACKSON R B. Root dynamics and global change: seeking an ecosystem perspective[J]. New Phytologist,2000,147(1):3-12.
[3]BRASSARD B W, CHEN H Y H, BERGERON Y, et al. Differences in fine root productivity between mixed-and single-species stands[J]. Functional Ecology,2011,25(1):238-246.
[4]WANG X L, KLINKA K, CHEN H Y, et al. Root structure of western hemlock and western red cedar in single-and mixed-species stands[J]. Canadian Journal of Forest Research,2002,32(6):997-1004.
[5]LEUSCHNER C, HERTEL D, CONERS H, et al. Root competition between beech and oak: a hypothesis[J]. Oecologia,2001,126(2):276-284.
[6]BAUHUS J, KHANNA P K, MENDEN N. Aboveground and belowground interactions in mixed plantations ofEucalyptusglobulusandAcaciamearnsii[J]. Canadian Journal of Forest Research,2000,30(12):1886-1894.
[7]张健,郝占庆,宋波,等.长白山阔叶红松林中红松与紫椴的空间分布格局及其关联性[J].应用生态学报,2007,18(8):1681-1687.
[8]胡艳波,惠刚盈,戚继忠,等.吉林蛟河天然红松阔叶林的空间结构分析[J].林业科学研究,2003,16(5):523-530.
[9]安慧君.阔叶红松林空间结构研究[D].北京:北京林业大学,2003.
[10]李俊清,王业遽.红松人工幼林透光伐效果分析[J].应用生态学报,1991,2(1):8-13.
[11]张春锋,殷鸣放,孔祥文,等.不同间伐强度对人工阔叶红松林生长的影响[J].辽宁林业科技,2007,33(1):12-15.
[12]杨丽韫,李文华.长白山原始阔叶红松林细根分布及其周转的研究[J].北京林业大学学报,2005,27(2):1-5.
[13]朱胜英,周彪,毛子军,等.帽儿山林区6种林分细根生物量的时空动态[J].林业科学,2006,42(6):13-19.
[14]杨丽韫,罗天祥,吴松涛.长白山原始阔叶红松(Pinuskoraiensis)林及其次生林细根生物量与垂直分布特征[J].生态学报,2007,27(9):3609-3617.
[15]王树堂,韩士杰,张军辉,等.长白山阔叶红松林表层土壤木本植物细根生物量及其空间分布[J].应用生态学报,2010,21(3):583-589.
[16]张万儒,杨光滢,屠星南.森林土壤分析方法[M].北京:中国标准出版社,1999.
[17]SCHMID I, KAZDA M. Root distribution of Norway spruce in monospecific and mixed stands on different soils[J]. Forest Ecology and Management,2002,159(1):37-47.
[18]LOREAU M. Biodiversity and ecosystem functioning: a mechanistic model[J]. Proceedings of the National Academy of Sciences,1998,95(10):5632-5636.
[19]JOSE S. Black walnut allelopathy: current state of the science[M]//MALIK A U, INDERJI T. Chemical Ecology of Plants: Allelopathy in Aqutic and Terrestrial Ecosy Stems. Birkhauser Basel: Plant Signal Transduction,2002:159-167.
[20]陈硕芃,王韶仲,谷加存,等.密度结构对大青川红松人工林细根生物量与根长密度的影响[J].森林工程,2013,29(4):1-7.
[21]RICHARDS A E, FORRESTER D I, BAUHUS J, et al. The influence of mixed tree plantations on the nutrition of individual species: a review[J]. Tree Physiology,2010,30(9):1192-1208.
[22]JOSE S, WILLIAMS R, ZAMORA D. Belowground ecological interactions in mixed-species forest plantations[J]. Forest Ecology and Management,2006,233(2):231-239.
Influence of Species Mixing on Fine Root Biomass and Spatial Distribution inPinuskoraiensisPlantation//
Qian Wenli
(Northeast Forestry University, Harbin 150040, P. R. China); Lu Yuan(Liaoning Institute of Forest Inventory and Planning); Wang Shaozhong, Gu Jiacun(Northeast Forestry University)//Journal of Northeast Forestry University,2016,44(2):1-5.
Mixed-species forest plantation;Pinuskoraiensis;Fraxinusmandschurica;Juglansmandshurica; Fine root
钱文丽,女,1991年1月生,东北林业大学林学院,硕士研究生。E-mail:554931491@qq.com。
谷加存,东北林业大学林学院,副教授。E-mail:gjcnefu@163.com。
2015年8月20日。
S725.2
1)国家林业科技支撑计划课题(2012BAD21B0202)。
责任编辑:潘华。
