人造林窗下香椿幼林细根的养分内循环
2018-09-04周义贵刘运科李德会李贤伟
周义贵,刘运科,苏 宇,李德会,李贤伟*
(1.四川省林业调查规划院,成都 610081;2.四川农业大学林学院,成都 611130;3.成都市农林科学院,成都 611130)
养分内循环指植物体内器官或组织之间的养分转移与交换。大量研究表明,叶片在衰老过程中会将氮、磷、钾等养分元素转移到其他组织[1]。细根(直径<2 mm)是植物根系的重要组成部分,是植物从土壤中捕获养分和水分的主要器官。虽然细根生物量仅占林分总生物量的0.5%~10%[2],但细根寿命短、周转快,在频繁的死亡过程中可将大量养分归还给土壤,同时也通过养分内循环将养分转移给其他器官或组织,以减少自身养分损失,降低对环境养分供应的依赖。因此,细根的养分内循环是植物保持营养的重要策略之一[1,3-4]。尽管如此,目前关于细根衰老死亡过程中是否存在养分内循环仍然存在很大争议[5]。有研究认为,细根存在养分内循环,衰老的细根在脱落前可以转移23%的氮、30%的磷和39%的钾[6]。而E.S.Nambiar等[7]却提出质疑,认为直接比较死根和活根之间的养分差异来确定细根衰老死亡过程中是否存在养分内循环的方法有明显缺陷。在无法准确获取细根死亡时间的基础上,直接从林地中采集的死亡细根不能确定是否发生分解,更不能精确分解量及养分浓度变化率,这使得研究结论缺乏可信度[7]。随着研究方法的不断完善,将细根剥离土壤后置于独立空间,则能够避免细根的分解对养分内循环的影响。吴勇等[8]利用黑色塑料瓶将剥离土壤的细根套住、密封,模拟细根衰老的方法获得了柳杉(Cryptomeria fortunei)1~3级细根养分内循环数据,陈珊等[5]也验证了该方法的可行性。相对于地上部分而言,植物根系的外部环境更为复杂,不断改善方法,对研究养分元素的迁移规律,探讨细根养分内循环将具有重要意义。
香椿(Toona sinensis)是亚热带地区珍贵的乡土阔叶树种,在退化人工针叶纯林的结构调整和生态功能提升中具有先天的优势,可作为改善林分结构、提升森林质量的重要备选树种[9]。本试验选取川中丘陵区柏木低效林近自然经营改造模式,通过“开窗补阔”,以补植的香椿为对象,采用吴勇等[8]的方法,将细根剥离土壤后置于独立空间,在确保其不分解的前提下,定期获取不同根序级的衰老细根,并分析其氮、磷、钾、钙、镁等主要养分元素动态,确定细根在衰老死亡过程中养分元素是否发生内循环及其速率大小。从养分利用效率角度,为退化森林生态系统恢复过程中的树种选择提供数据支撑。以期为实现林分“混交-异龄-复层”的结构,形成优质、健康、高效、稳定的森林生态系统提供一定的理论依据。
1 材料和方法
1.1 研究区概况
试验地位于四川省德阳市旌阳区和新镇(104°25′E,31°04′N),海拔 510~540 m,属于低山丘陵地貌。该区属亚热带湿润和半湿润气候区,年均气温16~17℃,年总降水量880~940 mm,主要集中在5—10月,冬干春旱。年均无霜期 270~290 d[10],土壤多为紫色土。该区在20世纪80年代营造了大面积的柏木纯林,因树种单一、初植密度过大及经营管理滞后,导致林分郁闭度大、层次结构简单、物种多样性低、地力衰退、林分长势普遍衰弱。现平均树高6.5 m,平均胸径8.0 cm,郁闭度大于0.8,单位蓄积量为44.7 m3/hm2,远低于83.6 m3/hm2的全国平均水平,是典型的低产低效林[11]。林下灌木以黄荆(Vitex negundo)、马桑(Coriaria nepalensis)、火棘(Pyracantha fortuneana)、铁仔(Myrsine africana)为主;草本植物以凤尾蕨(Pteris cretica)、栗褐苔草(Carex brunnea)、荩草(Arthraxon hispidus)为主。
1.2 样地设置
2012年3月,选择在西南坡向、中坡位、坡度20°~30°、生长状况及经营管理水平相似的地段进行林窗式采伐[10]。人造林窗为以南北向长、东西向短的近椭圆形,面积为200 m2,3次重复,各林窗之间预留5~10 m的过渡带。将树枝树干等残体移除,但未掘根。处理完成后,于2012年3月下旬在每个林窗中引种香椿,采用“品”字形的方式,按50 cm×50 cm×50 cm的规格窝穴栽种,株行距为1 m×1.5 m,每窝穴施基肥2.5~3.0 kg[9]。初植香椿为2 a生实生苗,平均树高30 cm,平均地径0.43 cm。期间,只对香椿周边进行折灌、除草处理,没有施肥等其他经营措施。2015年4月对样地基本情况进行调查,3个样地的香椿长势良好,平均树高6.0 m,胸径4.53 cm,平均冠面积3.87 m2。
1.3 试验方法
试验开始于2015年4月下旬。在3个样地分别选取5株香椿平均木,在林下距树基1m处移除枯落物。参照Guo D.L.等[12]的根序分级方法,以根轴末端未分枝的根分为1级细根,1级细根着生于2级细根上,依次分到5级细根,对于生于高级细根(3~5级细根)上的没有分支的细根也分为1级细根,用木刀轻轻去除表层土壤,露出1级细根,沿1级细根仔细向2级细根搜寻,直至找到5级细根分枝处,将该根束分蘖的所有1~5级细根小心从土壤中剥离出来,用刷子轻轻刷去附着在细根上的泥土,将根束全部放置于内径6 cm,深25 cm的黑色塑料瓶中,瓶口用防水胶密封,防止土壤水、土壤动物和林地凋落物等杂物进入,然后将塑料瓶垂直于地面埋回原地土壤10~20 cm土层中[8],既保证细根衰老过程中能正常呼吸,也避免了细根分解对养分内循环的影响。每株样树重复5次,共获得75个根束,每根束细根鲜重约5 g。从实验当天开始,每间隔7 d分别从每株样树上截取1个根束,每次采集15个根束,共采集5次(采集时间分别为:2015-4-23,2015-4-30,2015-5-7,2015-5-14,2015-5-21)。根束采集后放入自制便携式冰箱立即带回实验室。用去离子水清洗干净,测定各级根序形态参数(表1),在65℃烘箱中烘烤48 h后粉碎保存。分别采用凯氏定氮法测氮,钼锑抗比色法测磷,原子吸收分光光度法测定钾、钙、镁的浓度[13]。采用陈珊等[5]的方法计算各级细根养分内循环率。
N级细根养分内循环率/%=N级细根期初与期末养分浓度变化量/N级细根期初养分浓度×100%
式中,N=1,2,…,5,期初浓度为 2015-4-23取样养分浓度;期末浓度为2015-5-21取样养分浓度。

表1 香椿1~5级细根根长、直径、比根长Table1 1~5 fine root length,diameter,and specific root length of Toona sinensis
1.4 数据处理与统计分析
所有数据均采用Microsoft Excel 2010、SPSS 20.0进行处理与分析。采用单因素方差分析(one-way ANOVA)分析香椿1~5级细根养分浓度和内循环率。
2 结果与分析
2.1 香椿1~5级细根初始养分浓度
从表2可知,香椿1~5级细根中氮的初始浓度最大,其次是钾、钙、镁、磷,分别为 4.85~18.75 g/kg、4.01~5.19 g/kg、2.93~3.25 g/kg、1.46~1.59 g/kg、0.76~1.13 g/kg,其中氮、磷、钾浓度随着根序的增加而降低。方差分析显示,氮浓度在1、2级细根之间差异不显著(P>0.05),但均显著高于 3~5级细根(P<0.05);磷浓度在 1级细根中显著最高(P<0.05),在 2、3 级细根及4、5级细根差异不显著(P>0.05),但二者之间差异显著(P<0.05);钾浓度在相邻根序间差异不显著(P>0.05),但在低级细根(1、2级)中显著高于高级细根;钙、镁浓度在各根序间差异不显著(P>0.05)。
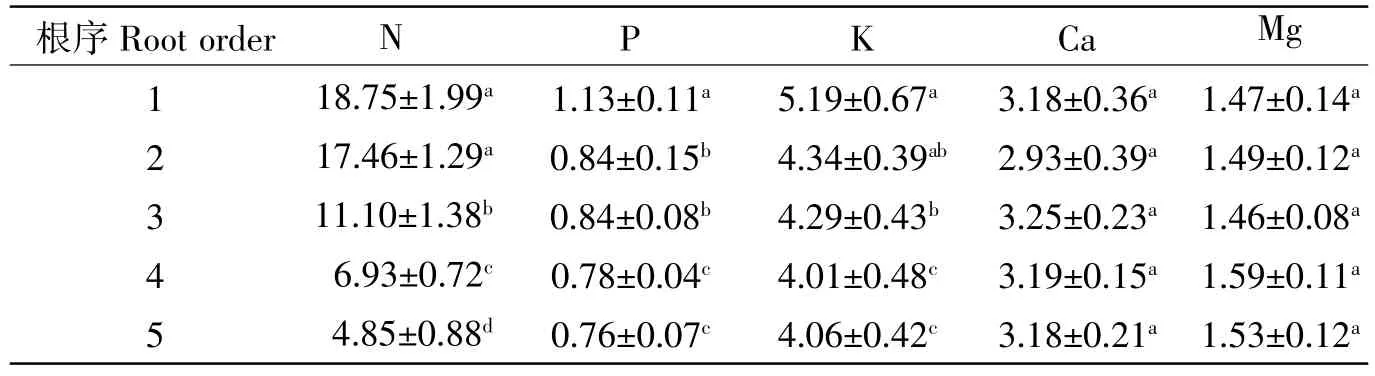
表2 香椿1~5级细根初始养分浓度Table2 1~5 fine root initial nutrient concentration of Toona sinensis g·kg-1
2.2 香椿细根衰老过程中养分浓度变化
2.2.1 香椿细根衰老过程中氮浓度变化
由图1可知,不同等级根序中的氮浓度变化规律均不相同。1级细根的氮浓度呈现下降趋势,累计降低了30.56%。2、3、4级细根的氮浓度均表现为先升后降,说明氮在细根衰老过程中发生迁移,存在内循环。由于还未衰老的2级细根接受了从1级细根中转移出来的氮,其浓度在处理3周后达到峰值,为20.93 g/kg。处理3周后开始表现出衰老特征,氮元素迁出速率明显加快,2级细根氮累计迁移率为46.49%。3级细根的氮浓度变化规律与2级细根的氮浓度变化相似,3级细根氮累计迁移率为25.68%。4级细根中氮在处理4周后出现峰值,为8.71 g/kg,表明氮元素的迁移伴随着不同根序细根的衰老而发生。5级细根的氮浓度变化基本稳定,说明在5周的试验期间内,5级细根主要起养分运输作用。
2.2.2 香椿细根衰老过程中磷浓度变化
如图1所示,细根在衰老过程中磷浓度变化与氮元素相似,也存在内循环。1级细根的磷浓度均处于下降过程,且在处理1周后有一个剧烈的下降过程,但在处理2周后下降趋势明显减缓,说明1级细根的磷迁移较为迅速,累计降低了0.46 g/kg。由于2级细根接受了从1级细根迁移出来的磷,其浓度在第2周达到峰值,为1.23 g/kg,说明处理2周后开始表现出衰老特征,磷元素迁出速率明显加快,2级细根磷累计迁移率为56.86%。3级细根磷浓度变化与2级细根变化相似,同样在处理2周后即达到峰值。相对于2~3级同根序细根的氮浓度变化,磷的迁移发生则更为迅速。4、5级细根中的磷浓度则没有明显的变化趋势。
2.2.3 香椿细根衰老过程中钾浓度变化
钾元素是极易移动的养分元素,同样在细根衰老过程中存在内循环,且在处理的第1周就迅速做出反应。如图1所示,1级细根的钾浓度在处理第1周就迅速下降,之后呈现较平缓的变化,1级细根钾累计迁移率为21.77%。2级细根的钾浓度在第2周达到峰值,说明2级细根接受了从1级细根迁移出来的钾,2周后开始表现出衰老和养分转移的特性。3、4级细根的钾浓度先升后降,均在3周后开始表现出养分转移的特性。5级细根的钾浓度无明显变化。
2.2.4 香椿细根衰老过程中钙和镁浓度变化
由图1可知,1~5级细根在衰老过程中钙浓度变化均不显著,基本在 2.93~3.25 g/kg之间,1、2、4级细根的钙随时间变化呈“W”型的趋势,3、5级细根变化规律不明显。镁浓度在各级根序中的变化均不同,且变化幅度均较小。1、4级细根的镁浓度在处理前期下降,在1周后上升,但1级细根的镁浓度变化幅度比4级细根大。2、3级细根的镁浓度在1周后下降,随后开始上升,在处理第3周达到峰值。第4周2级细根镁浓度开始下降,而3级细根的镁浓度趋于稳定的水平。5级细根的镁浓度处于波动状态,无明显规律变化。表明香椿细根衰老过程中,钙、镁可能存在内循环,但迁移率极小。
2.3 香椿细根衰老过程中的养分内循环率
由表3可知,香椿1~5级细根在衰老过程中,氮、磷、钾养分迁移明显,存在内循环。氮、磷和钾的内循环率平均为16.41%、21.95%和17.09%,主要发生在1级和2级细根中,其中磷的内循环率最大,其次是氮、钾。氮、磷的养分内循环在2级细根中最显著,其内循环率分别高达29.58%、33.97%,钾的养分内循环则主要集中在1级细根,其内循环率为22.11%,均远高于3~5级细根。钙和镁元素在细根衰老过程中浓度变化不大,说明这2种元素移动性较差(浓度变化率较小或为负值,暂不分析内循环率)。方差分析显示,同一元素在1~2级细根以及4~5级细根中的内循环率无显著差异(P>0.05),但低级细根(1、2级)养分内循环显著高于高级细根(3~5级)(P<0.05),3级细根与 4、5级细根之间的养分内循环差异显著(P<0.05)(3级细根作为过渡性根),说明在分析香椿的细根养分内循环时可以将1~2级细根和3~5级细根分别归为同一结构模块进行研究。

图1 香椿细根衰老过程中养分浓度变化Figure1 The change of Nutrient concentration in Toona sinensis fine root with senescence

表3 1~5级香椿细根元素的内循环率Table3 Nutrient internal cycling rate of different root order of Toona sinensis %
3 讨论
自20世纪80年代,一些学者开始关注细根养分内循环研究,但因树种、研究方法等的差异,细根衰老过程养分内循环的机理至今仍无定论[1,14]。多数研究者认为树木的细根存在养分内循环[8,15],也有研究认为这只不过是大多数植物的一个特征,部分树木的细根基本不存在养分的转移。有学者通过比较研究发现,同一树种的细根中有的元素存在内循环,有的元素则不发生迁移。已开展的细根养分内循环的研究大多数为直接比较活根和死根之间的养分差异来确定是否存在养分内循环及内循环率的大小,其结果会受到细根分解的影响,所得结论的可靠性值得商榷[5,8]。由此可见,细根养分内循环研究的关键是获取没有分解的衰老或死亡细根[8]。如果将待研究植物细根剥离土壤,放置在无法接触土壤微生物、养分和水分的独立空间,人为促进细根的衰老死亡进程,避免了细根分解对养分内循环的影响,对于细根衰老过程中养分内循环的正确评估将有积极的意义[8]。本研究将香椿细根置于独立空间,避免了衰老过程中的分解对养分内循环的影响[5]。结果表明,香椿细根在衰老过程中氮、磷、钾元素有明显的迁移现象,存在养分内循环,其速率分别为4.14%~29.58%、10.06%~33.97%、13.23%~22.11%,养分循环利用效率较高。且3种元素的迁移均先发生在寿命短、周转快的1级细根中,随后向2级及更高根序级细根移动,但2~3级细根的磷浓度峰值比氮、钾出现的早,说明在细根开始衰老时,磷更容易转移,且转移更迅速;钙和镁元素则基本不发生转移,这与养分元素自身的移动性较差有关,二者在植物体内可能是以稳定化合物的形式存在[5]。香椿1~5级细根中氮、磷和钾的内循环率平均为16.41%、21.95%和17.09%,且养分内循环主要集中在1~2级细根,其内循环率远大于3~5级细根,这说明低级细根(1~2级)对养分内循环的贡献率高于高级细根(3~5级)。主要是因为,一方面低级细根具有较高的氮、磷、钾浓度(表2),而养分浓度是影响养分内循环率的重要因素[5,17]。另一方面,根据植物叶子寿命与内循环强度成反比可推测细根养分内循环可能与细根寿命有密切关系,低级细根寿命显著短于高级细根[17-20],细根根序等级可能是衡量细根养分内循环的重要依据[5]。
根系的研究者一直试图在根系中找到一些类似于功能上某个模块的功能单元。如在北美的9个树种的根系上发现了根痕,进而推测细根像叶子一样,每年或者不定期的成簇脱落[18]。随后,Guo D.L.等[12,18]通过解剖特征分析初生和次生结构、菌根浸染等进一步论证了1级和2级细根成为一个统一功能模块的可能。本研究对香椿1~5级细根养分内循环进行了测定,结果表明同一养分元素在1~2级细根以及3/4~5级细根中的内循环率内无显著差异,但低级细根养分内循环显著高于高级细根,3级细根与4、5级细根之间的养分内循环差异显著,说明在分析香椿的细根养分内循环时可以将1~2级细根、3级细根和4~5级细根分别归为不同结构模块进行研究。但不同结构模块在吸收养分中是否有不同功能,内循环的机理是否一致,仍待进一步研究。
目前多数的研究者认为,养分内循环是植株为适应贫瘠环境而提高养分循环利用率的一种自我保护机制[1,5]。Chen X.D.等[21]研究了混交刺槐后,侧柏林长势明显变好,认为刺槐进入后,能够有效地促进养分循环。本研究证明香椿细根存在养分内循环,对于引入树种适应柏木低效林林地贫瘠的土壤环境,提高低效林改造成功率有重要意义。因此,川中丘陵区柏木低效林“开窗补阔”生态改造模式引进的乡土阔叶树种香椿,与柏木林构成针阔混交的块状镶嵌格局,不仅改善了低效柏木林的景观格局,增加了系统稳定性,而且香椿具有较高的光合效率和生产力[9],林分固碳能力强[11,22]。因此,在与本研究选取的条件相似的地区,可将香椿作为柏木纯林等退化人工林改造中的优良备选树种推广使用。
