4个观赏竹种的光合特性及其影响因子分析
2016-08-12黄程前黄文韬
黄 滔, 刘 玮, 唐 红, 黄程前, 黄文韬, 唐 军
(1. 湖南省森林植物园, 湖南 长沙 410116; 2. 中南林业科技大学, 湖南 长沙 410004)
4个观赏竹种的光合特性及其影响因子分析
黄滔1,2, 刘玮1, 唐红1, 黄程前1, 黄文韬1, 唐军1
(1. 湖南省森林植物园, 湖南 长沙 410116; 2. 中南林业科技大学, 湖南 长沙 410004)
本研究以白纹阴阳竹(Hibanobambusatranquillansf.shiroshimaH. Okamura)、鼓节竹(Bambusatuldoides‘Swolleninternode’)、花秆早竹(Phyllostachysviolascensf.viridisulcataP. X. Zhang et W. X. Huang)和美丽箬竹(IndocalamusdecorusQ. H. Dai)4个观赏竹种为供试材料,分析了各竹种的光响应和CO2响应曲线及参数、光合和气体交换参数以及相关环境因子的日变化规律,并通过相关性分析、逐步多元回归分析和通径分析探讨了影响4个竹种叶片净光合速率(Pn)的主要因子。测定结果表明:随光合有效辐射强度(PAR)或胞间CO2浓度(Ci)的提高,各竹种叶片的Pn值均逐渐增大,但增幅有一定差异;各竹种的光响应和CO2响应参数均有较大差异,其中,白纹阴阳竹叶片的表观量子产量(AQY)最低,CO2饱和点(CSP)和CO2补偿点(CCP)也较低,其他参数均最高;而花秆早竹叶片的AQY和CCP值均最高,而其他参数总体上均最低。4个竹种的光合及气体交换参数日变化曲线均有一定差异;美丽箬竹叶片Pn值日变化曲线呈“双峰型”;白纹阴阳竹、鼓节竹和花秆早竹叶片的Pn值日变化曲线均呈“单峰型”,峰值分别出现在14:00、10:00和12:00;总体上看,4个竹种叶片的气孔导度(Gs)、Ci、蒸腾速率(Tr)和水分利用效率(WUE)总体上具有相似的日变化趋势。分析结果表明:按照总影响效应由高至低进行排序,对白纹阴阳竹叶片Pn值的主要影响因子为PAR、Tr、大气CO2浓度(Ca)、Ci,对鼓节竹叶片Pn值的主要影响因子为PAR、Tr、Ca、Gs,对花秆早竹叶片Pn值的主要影响因子为PAR、Tr、Gs、气温(Ta)、Ci,对美丽箬竹叶片Pn值的主要影响因子为PAR、空气相对湿度(RH)、Ta、Ca、Tr、Gs。综合分析结果显示:4个竹种中,白纹阴阳竹光合能力最强,但对弱光的适应和利用能力较弱,CO2同化能力强且适应范围广;鼓节竹和美丽箬竹对光和CO2的利用能力和适应范围均居中;花秆早竹光合能力和CO2同化能力最差,但对弱光的适应和利用能力较强,CO2适应范围较窄。
观赏竹种; 光合特性; 气体交换参数; 环境因子; 相关性分析; 通径分析
园林植物的光合生理特性与其自身的遗传特性和周围的环境因素紧密相关,是植物栽培过程中重要的依据指标之一[1-2]。竹类植物是重要的园林绿化植物,种类繁多、用途广泛。近年来,观赏竹的生态和景观功能日益受到重视,市场前景十分广阔。竹类的研究主要集中在生长规律[3-4]和光合特性[5-8]方面,对其无性繁殖[9-10]和分子生物学[11-12]方面也有相关研究报道。研究结果表明,不同竹种的光合生理特性差异较大,且影响光合作用的主要因素也不尽相同[5-8],因此,应该按照不同竹种的光合特性进行合理配置、栽培和管理。
白纹阴阳竹(Hibanobambusatranquillansf.shiroshimaH. Okamura)隶属于阴阳竹属(HibanobambusaMaruy. et H. Okamura),属混生竹种;新叶黄白色,渐绿间有较宽的黄白色条纹,叶色亮丽,是极具观赏价值的观叶竹种。鼓节竹(Bambusatuldoides‘Swolleninternode’)隶属于簕竹属(BambusaSchreber);秆高6~8 m,竹秆节间下部缩短膨大,秆形奇特,为优良的园林观秆竹种。花秆早竹(Phyllostachysviolascensf.viridisulcataP. X. Zhang et W. X. Huang)隶属于刚竹属(PhyllostachysSieb. et Zucc.),属散生竹种;秆金黄色,沟槽白色,为珍稀的观秆竹种。美丽箬竹(IndocalamusdecorusQ. H. Dai)隶属于箬竹属(IndocalamusNakai);叶片呈带状披针形,叶片较大,在园林中应用广泛,是优良的观叶竹种。目前这4个观赏竹种尚未被广泛推广,因此,研究这4个观赏竹种的光合生理特性,并有针对性的制定各竹种的栽培措施,有助于对它们进行合理推广和应用。
为揭示上述4个观赏竹种的光合生理特性,作者对它们的光响应和CO2响应特征及光合参数日变化进行了研究,同时还分析了各竹种的净光合速率与部分生理生态因子的关系,以期掌握各竹种的光合作用规律,为筛选出适合不同生态环境的栽培竹种,以及不同竹种的推广种植、配置造景、植被恢复和生态建设等提供基础研究数据。
1 材料和方法
1.1材料
供试白纹阴阳竹、鼓节竹、花秆早竹和美丽箬竹均为3年生竹,种植于湖南省森林植物园观赏竹专类园,地理坐标为东经113°00′、北纬28°20′,海拔70~85 m。种植地年均气温17.2 ℃,极端高温40.6 ℃,极端低温-11.3 ℃;年平均降水量1 412.3 mm;全年无霜期281 d。主要土壤类型为第四纪网纹红壤,土壤pH 5.6左右。
选取同一地块不同栽植位置且生长旺盛、长势一致的健壮植株为样株,各竹种样株的平均秆高和平均秆直径见表1。


竹种Bamboospecies平均秆高/mAverageheightofculm平均秆直径/cmAveragediameterofculmHT0.82±0.080.85±0.04BT2.83±0.095.14±0.26PV4.92±0.313.58±0.21ID1.17±0.130.49±0.06
1)HT: 白纹阴阳竹Hibanobambusatranquillansf.shiroshimaH. Okamura; BT: 鼓节竹Bambusatuldoides‘Swolleninternode’; PV: 花秆早竹Phyllostachysviolascensf.viridisulcataP. X. Zhang et W. X. Huang; ID: 美丽箬竹IndocalamusdecorusQ. H. Dai.
1.2方法
1.2.1光响应曲线和CO2(ACI)响应曲线测定在2014年的9月2日、9月16日和9月28日的9:00至11:00,每个竹种选取样株3株,每样株各选取中部向阳的叶片3枚,用LI-6400便携式光合仪(美国LI-COR公司)测定叶片的净光合速率,结果取平均值。
测定光合响应曲线时,设定光合测定系统的叶室温度(25±1) ℃、胞间CO2浓度(Ci)400 μmol·mol-1、气体流速0.5 L·min-1、空气相对湿度(RH)70%,在光合有效辐射强度(PAR)为0、25、50、100、150、200、400、 600、 800、 1 000、 1 200、 1 400、 1 600和1 800 μmol·m-2·s-1条件下测定叶片净光合速率(Pn)。
测定CO2响应曲线时,安装CO2钢瓶后稳定20 min,按照上述条件设置测定系统的叶室温度、气体流速和空气相对湿度,PAR设为1 200 μmol·m-2·s-1,在胞间CO2浓度为50、 100、 200、 300、400、 600、800、1 000和1 200 μmol·m-2·s-1条件下测定叶片Pn值。
1.2.2光合和气体交换参数以及环境因子日进程测定于2014年9月选3个典型晴天,在每天的6:00至18:00测量光合和气体交换参数的日变化。各种竹均选择样株3株,每一样株各选择中部向阳的叶片3枚并挂牌标记,测定时保持叶片自然生长角度不变,每枚叶片重复测定3次。用LI-6400便携式光合仪每隔2 h测定1次。
光合和气体交换参数包括净光合速率、气孔导度(Gs)、胞间CO2浓度和蒸腾速率(Tr),并计算叶片水分利用效率(WUE)。同时对相关环境因子的日变化进行测定,包括光合有效辐射强度、气温(Ta)、空气相对湿度和大气CO2浓度(Ca)。
1.3数据处理和统计分析
采用EXCEL 2010软件和SPSS 17.0统计分析软件进行数据处理、方差分析、相关性分析和逐步多元回归分析等。另外,采用叶子飘等[13]的光合新模型对光响应曲线和CO2响应曲线进行拟合,并据此计算光响应参数和CO2响应参数。
2 结果和分析
2.1供试4个竹种叶片的光响应曲线及光响应参数分析
供试4个竹种叶片的光响应曲线见图1。结果显示:随光合有效辐射强度(PAR)的升高,4个竹种叶片的净光合速率(Pn)变化趋势相似,均呈先急剧升高后趋于平稳的变化趋势,但各竹种叶片的Pn值差异较大。
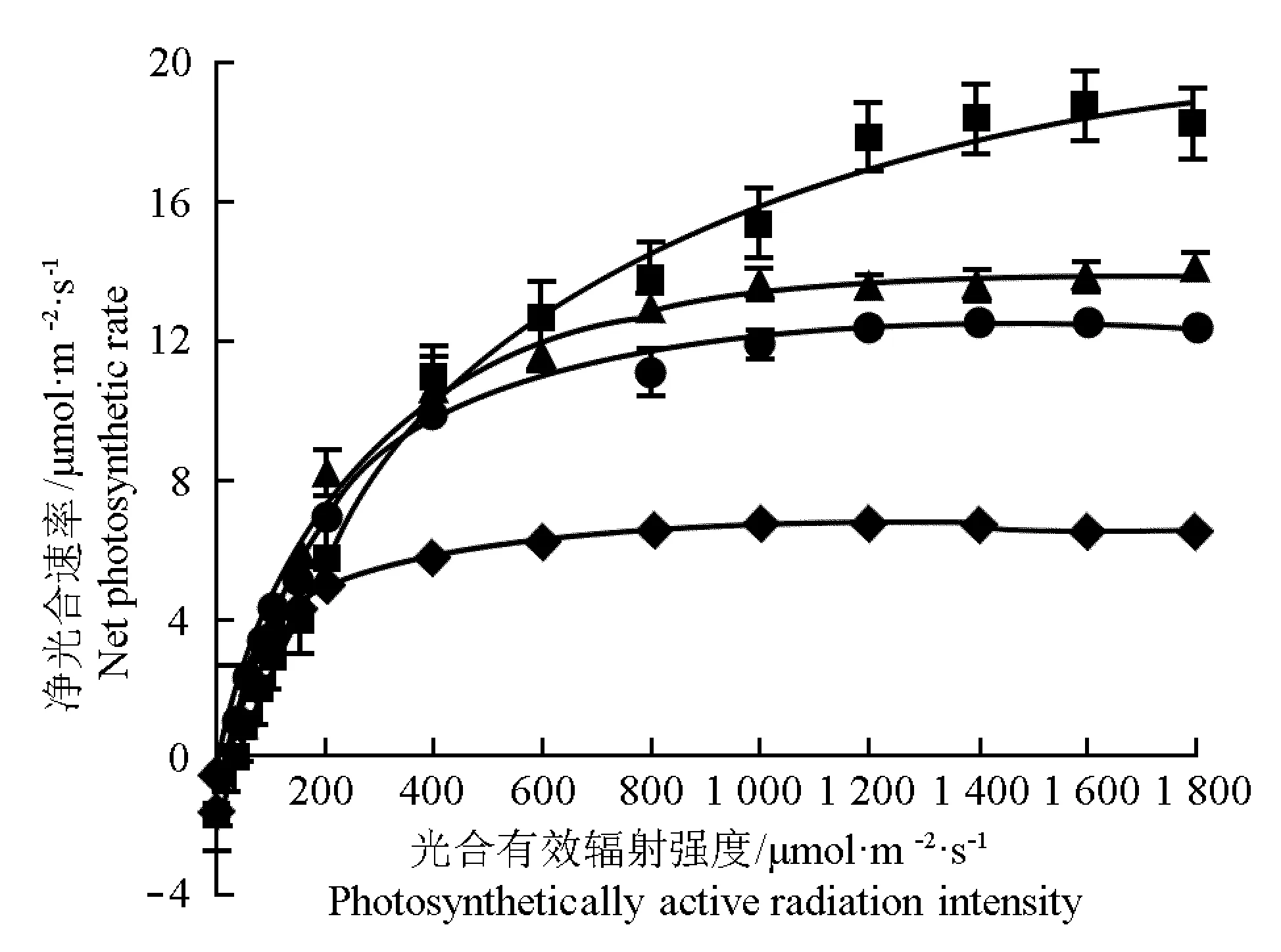
—■—: 白纹阴阳竹Hibanobambusatranquillansf.shiroshimaH. Okamura; —▲—: 鼓节竹Bambusatuldoides‘Swolleninternode’; —◆—: 花秆早竹Phyllostachysviolascensf.viridisulcataP. X. Zhang et W. X. Huang; —●—: 美丽箬竹IndocalamusdecorusQ. H. Dai.
图1供试4个竹种叶片的光响应曲线
Fig. 1Light response curve of leaf of four bamboo species tested
以PAR值为自变量X、Pn值为因变量Y,利用叶子飘等[13]的光合新模型进行拟合,获得各竹种叶片的光响应回归方程,并据此计算出各竹种的光响应参数,结果见表2。由表2可见:4个竹种叶片的表观量子产量(AQY)、最大净光合速率(Pmax)、光饱和点(LSP)、光补偿点(LCP)和暗呼吸速率(Rd)均存在显著性差异。其中,白纹阴阳竹叶片的Pmax、LSP、LCP和Rd值在供试4个竹种中均最高,分别为20.70、 2 644.06、29.47和1.33 μmol·m-2·s-1,与其他3个竹种有显著差异(P<0.05),而其AQY值显著低于其他3个竹种,为47.34 mmol·mol-1,说明供试的4个竹种中白纹阴阳竹的光合能力最强,但对弱光的适应和利用能力较弱,与光响应曲线的变化结果一致,因此,当PAR值低于25 μmol·m-2·s-1时,由于白纹阴阳竹叶片的光合同化速率低于其消耗速率,因而其叶片的Pn值均为负值。鼓节竹与美丽箬竹叶片间各光响应参数均无显著差异(P>0.05),且各参数的数值在4个竹种居中,表明供试的4个竹种中鼓节竹与美丽箬竹光合能力居中。在4个竹种中,花秆早竹叶片的AQY值最高,其Pmax、LSP、LCP和Rd值均最低,说明花秆早竹对弱光的适应和利用能力在供试的4个竹种中最强,但其光合能力较弱,因此,当PAR值低于50 μmol·m-2·s-1时,花秆早竹的Pn值高于其他3个竹种,但随PAR值的升高,其Pn值则明显低于其他3个竹种。

竹种 BamboospeciesAQYPmaxLSPLCPRd白纹阴阳竹Hibanobambusatranquillansf.shiroshima47.34±13.16c20.70±0.67a2644.06±45.63a29.47±2.83a1.33±0.08a鼓节竹Bambusatuldoides‘Swolleninternode’69.18±12.37b13.57±0.59b1565.81±12.32b11.09±1.74b0.74±0.05b花秆早竹Phyllostachysviolascensf.viridisulcata86.81±34.33a6.46±0.58c1146.28±11.78c5.84±1.62b0.48±0.05c美丽箬竹Indocalamusdecorus65.63±26.17b12.26±0.64b1371.87±8.16bc9.36±1.73b0.59±0.04bc
1)AQY: 表观量子产量Apparent quantum yield (mmol·mol-1); Pmax: 最大净光合速率The maximum net photosynthetic rate (μmol·m-2·s-1); LSP: 光饱和点Light saturation point (μmol·m-2·s-1); LCP: 光补偿点Light compensation point (μmol·m-2·s-1); Rd: 暗呼吸速率Dark respiration rate (μmol·m-2·s-1). 同列中不同的小写字母表示差异显著(P<0.05) Different small letters in the same column indicate the significant difference (P<0.05).
2.2供试4个竹种叶片的CO2响应曲线及CO2响应参数分析
供试4个竹种叶片的CO2响应曲线见图2。结果显示:供试4个竹种叶片CO2响应曲线的变化趋势基本一致,但随胞间CO2浓度(Ci)的升高,不同竹种间Pn值差异明显增大。
以Ci值为自变量X、Pn值为因变量Y,利用叶子飘等[13]的光合新模型进行拟合,获得各竹种叶片的CO2响应回归方程,并据此计算出各竹种的CO2响应参数,结果见表3。由表3可见:白纹阴阳竹叶片的羧化效率(CE)、Pmax和光呼吸速率(Rp)在供试的4个竹种中最高,CO2饱和点(CSP)和CO2补偿点(CCP)则相对较低,表明白纹阴阳竹的羧化酶活性较高,RUBP(1,5-二磷酸核酮糖)的再生速率也较高,且对低浓度CO2有较强的利用能力和同化能力。鼓节竹叶片的CE、Pmax、CCP和Rp值略低于白纹阴阳竹,但差异不显著,而其CSP值则在供试的4个竹种中最高,说明鼓节竹的同化能力略弱于白纹阴阳竹,但差异不大。花秆早竹叶片的CE、Pmax和Rp值在4个观赏竹种中均最低,CCP值则最高,表明花秆早竹的CO2同化能力弱,并且对低浓度CO2的利用能力也较差。美丽箬竹叶片CSP值在4个竹种中最低,但其CE、Pmax和Rp值均略高于花秆早竹,但明显低于白纹阴阳竹和鼓节竹,表明美丽箬竹利用低浓度CO2的能力略优于花秆早竹,但整体对CO2的同化能力较弱。

—■—: 白纹阴阳竹Hibanobambusatranquillansf.shiroshimaH. Okamura; —▲—: 鼓节竹Bambusatuldoides‘Swolleninternode’; —◆—: 花秆早竹Phyllostachysviolascensf.viridisulcataP. X. Zhang et W. X. Huang; —●—: 美丽箬竹IndocalamusdecorusQ. H. Dai.
图2供试4个竹种叶片的CO2响应曲线
Fig. 2CO2response curve of leaf of four bamboo species tested


竹种 BamboospeciesCEPmaxCSPCCPRp白纹阴阳竹Hibanobambusatranquillansf.shiroshima55.35±18.23a21.34±0.71a1266.48±21.62b59.41±8.83bc3.11±0.12a鼓节竹Bambusatuldoides‘Swolleninternode’51.11±11.62a18.76±1.47a1784.26±22.02a51.11±9.74c2.65±0.14ab花秆早竹Phyllostachysviolascensf.viridisulcata15.65±8.72b8.84±0.68c1300.03±15.71ab88.23±8.66a1.33±0.18c美丽箬竹Indocalamusdecorus28.34±12.64b13.05±1.04b1118.20±18.17b64.01±7.84b1.75±0.22bc
1)CE: 羧化效率Carboxylation efficiency (mmol·mol-1); Pmax: 最大净光合速率The maximum net photosynthetic rate (μmol·m-2·s-1); CSP: CO2饱和点CO2saturation point (μmol·m-2·s-1); CCP: CO2补偿点CO2compensation point (μmol·m-2·s-1); Rp: 光呼吸速率Photorespiration rate (μmol·m-2·s-1). 同列中不同的小写字母表示差异显著(P<0.05) Different small letters in the same column indicate the significant difference (P<0.05).
2.3供试4个竹种叶片的净光合速率及部分光合和气体交换参数的日变化规律分析
供试4个竹种叶片的净光合速率(Pn)日变化曲线见图3;各竹种叶片的部分光合和气体交换参数的日变化曲线见图4。
2.3.1净光合速率的日变化规律由图3可以看出:白纹阴阳竹、鼓节竹和花秆早竹叶片的Pn值日变化曲线均呈“单峰型”,仅峰值出现的时间有一定差异,而美丽箬竹叶片的Pn值日变化曲线呈“双峰型”。白纹阴阳竹叶片的Pn值在6:00至10:00迅速上升,10:00至14:00缓慢上升,14:00达到峰值,之后迅速下降;鼓节竹叶片的Pn值在6:00至10:00迅速上升,10:00达到峰值,之后逐渐下降;花秆早竹叶片的Pn值在6:00至10:00迅速上升,10:00至12:00缓慢上升,12:00达到峰值,之后逐渐下降。美丽箬竹叶片的Pn值在6:00至10:00迅速上升,10:00达到峰值,在10:00至14:00迅速下降,之后又缓慢上升,至16:00达到次峰值,之后又迅速下降。总体上看,白纹阴阳竹叶片的Pn值明显高于其他3个竹种,花秆早竹叶片的Pn值明显低于其他3个竹种。

—■—: 白纹阴阳竹Hibanobambusatranquillansf.shiroshimaH. Okamura; —▲—: 鼓节竹Bambusatuldoides‘Swolleninternode’; —◆—: 花秆早竹Phyllostachysviolascensf.viridisulcataP. X. Zhang et W. X. Huang; —●—: 美丽箬竹IndocalamusdecorusQ. H. Dai.
图3供试4个竹种叶片的净光合速率日变化曲线
Fig. 3Diurnal change curve of net photosynthetic rate of leaf of four bamboo species tested
2.3.2光合和气体交换参数的日变化规律由图4可见:白纹阴阳竹叶片蒸腾速率(Tr)的日变化趋势与Pn值日变化趋势基本一致,在6:00至10:00迅速上升,10:00至12:00缓慢上升,之后迅速下降;其水分利用效率(WUE)日变化趋势与Pn值日变化趋势相反,6:00至10:00迅速下降,10:00至16:00基本稳定,之后缓慢回升;其气孔导度(Gs)在6:00至8:00迅速上升,之后迅速下降,10:00至14:00基本稳定,14:00至16:00缓慢下降,16:00后迅速下降;其Ci值在12:00前呈下降趋势,之后逐渐上升。
鼓节竹叶片Tr值日变化趋势与其Pn值日变化趋势基本一致;其WUE值日变化趋势则与其Pn值日变化趋势相反;其Gs值在10:00前迅速上升,之后逐渐下降;其Ci值在12:00前呈逐渐下降趋势,12:00至14:00缓慢上升,随后又呈下降后上升的波动趋势。
花秆早竹叶片Tr值日变化趋势与其Pn值日变化趋势基本一致;其WUE值在12:00前逐渐下降,12:00至16:00上升,至16:00后又下降;其Gs值在12:00前逐渐上升,之后逐渐下降;其Ci值在14:00前逐渐下降之后迅速上升。
美丽箬竹叶片Tr值日变化趋势与其Pn值日变化趋势有一定差异,其Tr值在12:00前逐渐上升,之后逐渐下降;其WUE值在12:00前逐渐下降,12:00至16:00上升,16:00后又下降;其Gs值在8:00前上升,之后波动下降;其Ci值在16:00前总体上呈逐渐下降的趋势,16:00之后则上升。

—■—: 白纹阴阳竹Hibanobambusatranquillansf.shiroshimaH. Okamura; —▲—: 鼓节竹Bambusatuldoides‘Swolleninternode’; —◆—: 花秆早竹Phyllostachysviolascensf.viridisulcataP. X. Zhang et W. X. Huang; —●—: 美丽箬竹IndocalamusdecorusQ. H. Dai.
A: 气孔导度日变化曲线 Diurnal change curve of stomatal conductance; B: 胞间CO2浓度日变化曲线 Diurnal change curve of intercellular CO2concentration ; C: 蒸腾速率日变化曲线 Diurnal change curve of transpiration rate; D: 水分利用效率日变化曲线 Diurnal change curve of water use efficiency.
图4供试4个竹种叶片光合和气体交换参数的日变化曲线
Fig. 4Diurnal change curves of photosynthetic and gas exchange parameters of leaf of four bamboo species tested
总体上看,供试4个竹种叶片的Gs、Ci、Tr和WUE值总体上具有相似的日变化趋势,但其中峰值出现的时间、各指标测定值的波动变化等方面存在一定的差异。
2.4供试4个竹种叶片与光合参数相关的环境因子日变化规律分析
供试4个竹种叶片与光合参数相关的环境因子日变化曲线见图5。由图5可见:气温(Ta)在6:00至10:00上升,10:00至16:00几乎维持不变,16:00后下降;与Ta值相反,空气相对湿度(RH)在6:00至10:00下降,10:00至16:00维持不变,16:00后又逐渐上升;大气CO2浓度(Ca)在6:00至12:00急剧下降,12:00时达到最低值,维持一段时间后在16:00后迅速上升;PAR值在6:00至12:00迅速上升,12:00达到峰值,之后逐渐下降,但在18:00的PAR值仍高于8:00。
2.5供试4个竹种叶片净光合速率与光合和气体交换参数及相关环境因子的统计分析
2.5.1相关性分析和逐步多元回归分析供试4个竹种叶片净光合速率(Pn)与其他光合和气体交换参数及相关环境因子间的简单相关系数见表4。结果表明:4个竹种叶片的Pn值与叶片的气孔导度(Gs)、胞间CO2浓度(Ci)、蒸腾速率(Tr)和水分利用效率(WUE)以及光合有效辐射强度(PAR)、气温(Ta)、空气相对湿度(RH)和大气CO2浓度(Ca)总体上有显著(P<0.05)或极显著(P<0.01)相关性。其中,各竹种叶片的Pn值与Gs、Tr、Ta、PAR值呈极显著或显著正相关,与Ci、Ca和RH值总体上呈极显著负相关,仅美丽箬竹叶片的Pn值与Ci值呈不显著的负相关(P>0.05)。

A: 气温日变化曲线 Diurnal change curve of air temperature; B: 空气相对湿度日变化曲线 Diurnal change curve of air relative humidity; C: 大气CO2浓度日变化曲线 Diurnal change curve of air CO2concentration; D: 光合有效辐射强度日变化曲线 Diurnal change curve of photosynthetically active radiation intensity.
图5供试4个竹种叶片与光合参数相关的环境因子日变化曲线
Fig. 5Diurnal change curves of environmental factors related to photosynthetic parameters of leaf of four bamboo species
表4供试4个竹种叶片净光合速率(Pn)与不同因子间的简单相关系数
Table 4Simple correlation coefficient between net photosynthetic rate (Pn) of leaf of four bamboo species tested and different factors
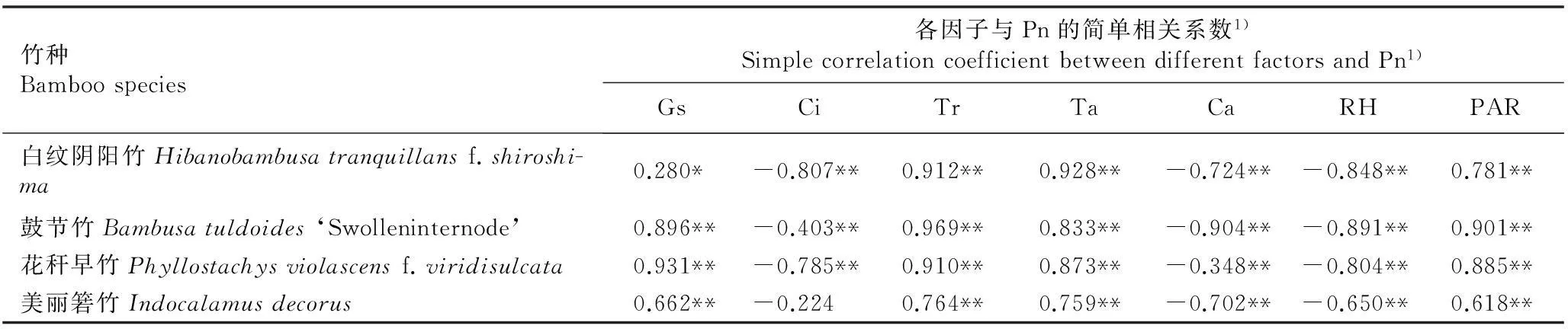
竹种Bamboospecies各因子与Pn的简单相关系数1)SimplecorrelationcoefficientbetweendifferentfactorsandPn1)GsCiTrTaCaRHPAR白纹阴阳竹Hibanobambusatranquillansf.shiroshi-ma0.280*-0.807**0.912**0.928**-0.724**-0.848**0.781**鼓节竹Bambusatuldoides‘Swolleninternode’0.896**-0.403**0.969**0.833**-0.904**-0.891**0.901**花秆早竹Phyllostachysviolascensf.viridisulcata0.931**-0.785**0.910**0.873**-0.348**-0.804**0.885**美丽箬竹Indocalamusdecorus0.662**-0.2240.764**0.759**-0.702**-0.650**0.618**
1)Gs: 气孔导度 Stomatal conductance; Ci: 胞间CO2浓度 Intercellular CO2concentration; Tr: 蒸腾速率 Transpiration rate; Ta: 气温 Air temperature; Ca: 大气CO2浓度 Air CO2concentration; RH: 空气相对湿度 Air relative humidity; PAR: 光合有效辐射强度Photosynthetically active radiation intensity. *:P<0.05; **:P<0.01.
对植物Pn值有影响作用的相关因子较多,简单相关性分析并不能准确反映它们之间的相互关系,为此,在前述简单相关性分析的基础上进一步分析了供试4个竹种叶片的Pn值与其他光合和气体交换参数及相关环境因子的偏相关性,结果见表5。结果表明:4个竹种叶片Pn值与各因子的偏相关性差异较大,与各竹种Pn值相关的主要因子并不一致。
利用逐步多元回归分析确定影响4个竹种叶片Pn值的主要因子,以Gs(X1)、Ci(X2)、Tr(X3)、Ta(X4)、Ca(X5)、RH(X6)和PAR(X7)为自变量,叶片Pn值为因变量Y,得到供试4个竹种叶片Pn值的最优回归方程分别为:Y白纹阴阳竹=1.572X3-0.861X2+1.346X5-0.222X7〔R2=0.981,P<0.01,标准估计误差(SEE) = 0.516〕;Y鼓节竹= 0.245X3- 0.286X5+ 0.316X7+ 0.233X1(R2= 0.989,P<0.01,SEE=0.499);Y花秆早竹= 0.288X1+ 0.199X7+ 0.741X4+0.595X2+ 0.324X3(R2= 0.981,P< 0.01,SEE=0.516);Y美丽箬竹= -0.402X3+ 0.663X4+ 0.659X1- 1.91X5+0.485X7+2.032X6(R2=0.951,P<0.01,SEE=0.820)。
由上述回归方程可知:白纹阴阳竹叶片Pn值的主要影响因子为Tr、Ci、Ca和PAR值;鼓节竹叶片Pn值的主要影响因子为Tr、Ca、PAR和Gs值;花秆早竹叶片Pn值的主要影响因子为Gs、PAR、Ta、Ci和Tr值;美丽箬竹叶片Pn值的主要影响因子为Tr、Ta、Gs、Ca、PAR和RH值,与偏相关分析结果基本一致。
表5供试4个竹种叶片净光合速率(Pn)与不同因子间的偏相关系数
Table 5Partial correlation coefficient between net photosynthetic rate (Pn) of leaf of four bamboo species tested and different factors

竹种Bamboospecies各因子与Pn的偏相关系数1)PartialcorrelationcoefficientbetweendifferentfactorsandPn1)GsCiTrTaCaRHPAR白纹阴阳竹Hibanobambusatranquillansf.shiroshi-ma0.014-0.791**0.821**0.257 0.637**0.254-0.520**鼓节竹Bambusatuldoides‘Swolleninternode’0.541**-0.3390.183**0.290* -0.378**-0.2250.688**花秆早竹Phyllostachysviolascensf.viridisulcata0.252*0.552*0.242*0.547** -0.066-0.1070.252*美丽箬竹Indocalamusdecorus0.379**0.225-0.358**0.321** -0.678**-0.504**0.401**
1)Gs: 气孔导度 Stomatal conductance; Ci: 胞间CO2浓度 Intercellular CO2concentration; Tr: 蒸腾速率 Transpiration rate; Ta: 气温 Air temperature; Ca: 大气CO2浓度 Air CO2concentration; RH: 空气相对湿度 Air relative humidity; PAR: 光合有效辐射强度 Photosynthetically active radiation intensity. *:P<0.05; **:P<0.01.
2.5.2通径分析对供试4个竹种叶片净光合速率(Pn)与其主要影响因子进行通径分析,结果见表6。由白纹阴阳竹的直接通径系数可知,Tr和Ca值对其Pn值有正效应,Ci和PAR值对其Pn值有负效应;各因子间的间接效应复杂,根据总通径系数,按照对白纹阴阳竹叶片Pn值的总影响效应由高至低排序,各主要影响因子依次为PAR、Tr、Ca、Ci。同理,按照对鼓节竹叶片Pn值的总影响效应由高至低排序,各主要影响因子依次为PAR、Tr、Ca、Gs;按照对花秆早竹叶片Pn值的总影响效应由高至低排序,各主要影响因子依次为PAR、Tr、Gs、Ta、Ci;按照对美丽箬竹叶片Pn值的总影响效应由高至低排序,各主要影响因子依次为PAR、RH、Ta、Ca、Tr、Gs。
表6供试4个竹种叶片净光合速率与其主要影响因子的通径分析结果1)
Table 6Result of path analysis on net photosynthetic rate of leaf of four bamboo species tested with main influence factors1)

主要影响因子Maininfluencefactor直接通径系数2)Directpathcoefficient2)间接通径系数3) Indirectpathcoefficient3)GsCiTrTaCaRHPAR总通径系数Totalpathcoefficient白纹阴阳竹Hibanobambusatranquillansf.shiroshima Ci-10.173—-0.75713.07223.95526.098 Tr0.8419.155—-12.659-24.870-27.532 Ca13.789-9.644-0.772—23.70527.079 PAR-27.7258.7890.754-11.790—-29.972鼓节竹Bambusatuldoides‘Swolleninternode’ Gs 0.003—0.0591.46818.59220.122 Tr0.0640.003—1.50724.39125.965 Ca-1.861-0.002-0.052—-20.701-22.615 PAR27.7060.0020.0561.391— 29.154
续表6Table 6 (Continued)

主要影响因子Maininfluencefactor直接通径系数2)Directpathcoefficient2)间接通径系数3) Indirectpathcoefficient3)GsCiTrTaCaRHPAR总通径系数Totalpathcoefficient花秆早竹Phyllostachysviolascensf.viridisulcata Gs0.004—-11.2190.1121.17935.26325.339 Ci12.916-0.004—-0.096-1.235-31.743-20.162 Tr0.1180.004-10.510—1.07236.26426.948 Ta1.3150.004-12.1310.096—31.48420.769 PAR38.7870.004-10.5700.1101.068—29.399美丽箬竹Indocalamusdecorus Gs0.009—-0.071 0.1560.8680.02911.31612.308 Tr-0.1070.006—0.66714.878-4.16646.71857.996 Ta1.0280.001-0.069—23.848-6.62746.67064.850 Ca-24.646-0.0000.065-0.994—6.723-45.339-64.192 RH6.8110.0000.065-1.000-24.326—-48.424-66.874 PAR58.5360.002-0.0850.81919.089-5.635—72.727
1)Gs: 气孔导度 Stomatal conductance; Ci: 胞间CO2浓度 Intercellular CO2concentration; Tr: 蒸腾速率 Transpiration rate; Ta: 气温 Air temperature; Ca: 大气CO2浓度 Air CO2concentration; RH: 空气相对湿度 Air relative humidity; PAR: 光合有效辐射强度 Photosynthetically active radiation intensity.
2)叶片净光合速率与主要影响因子的直接通径系数Direct path coefficient of net photosynthetic rate of leaf with main influence factors.
3)叶片净光合速率与主要影响因子的间接通径系数Indirect path coefficient of net photosynthetic rate of leaf with main influence factors.
3 讨论和结论
植物光合作用是一个十分复杂的生理过程,植物的光合特性一定程度上取决于其自身的遗传特性[1-2]。本研究中,供试4个竹种叶片的光合特性差异较大,除美丽箬竹外,白纹阴阳竹、鼓节竹和花秆早竹叶片净光合速率(Pn)日变化曲线均为“单峰型”。其中,白纹阴阳竹叶片Pn值明显高于其他竹种,其气孔在8:00后便可完全打开,导致Pn值迅速增加,随后在10:00至14:00之间Pn值有所提高但增幅不大,而其Gs值在8:00后迅速下降,Ci值在14:00也降到低谷,说明在正午阳光充足的条件下白纹阴阳竹的气孔会部分关闭。鼓节竹叶片Pn和Gs值在10:00时均达到峰值,12:00前Ci值呈下降趋势;花秆早竹叶片Pn值相对较低,在12:00时达到峰值,同时其Gs值最大,14:00前Ci值不断下降,鼓节竹和花秆早竹的Pn值达到峰值前其Gs和Ci值的变化趋势相似,说明在此期间二者的气孔均逐渐开放且光合作用较强,因此Ci值不断降低。美丽箬竹叶片的Pn值日变化曲线呈“双峰型”, Pn值在14:00降至最低,此时其Gs、Ci和Tr值均同时降低,说明美丽箬竹午间Pn值下降的原因与气孔限制因素有关[14]。
植物对光的响应可以基本阐释该植物的光合特性,其中,光饱和点(LSP)和光补偿点(LCP)可以反映植物对光能利用能力的强弱,一般认为,光饱和点高植物适应强光的能力强,光补偿点高即产物的消耗量大、表观量子产量(AQY)低则植物利用弱光的能力弱。CO2是光合同化的底物,其浓度直接影响植物的光合作用;CO2饱和点(CSP)高即植物对高浓度CO2适应性强,CO2补偿点(CCP)低则植物利用低浓度CO2能力强;羧化效率(CE)越高RuBP酶活性越高,从而使光合效率提高[15-17]。由供试4个竹种对光照和CO2的响应特征可见,4个竹种对光照和CO2的利用能力差异明显。其中,白纹阴阳竹叶片的LSP和LCP值均最高,同时其CE值也最高但AQY值最低,说明其光合能力最强,但利用弱光的能力差,产物消耗量也大;鼓节竹和美丽箬竹叶片的光响应参数在4个竹种中均居中,光合能力和CO2同化能力都低于白纹阴阳竹且高于花秆早竹,但鼓节竹的CE值显著高于美丽箬竹,这与其在10:00至14:00(强光条件下)Pn值高于美丽箬竹的结果相一致;花秆早竹叶片的LCP和LSP值均最低、AQY值最高,说明其光合能力最差且产物消耗量也低,但其对弱光的适应和利用能力较强,花秆早竹的CE值最低、CCP值最高,表明其CO2同化能力弱且适应范围窄。
简单相关性分析结果表明:供试4个竹种叶片Pn值与其他光合和气体交换参数及环境因子总体上显著相关,但其偏相关性分析结果则表明影响各竹种Pn值的主要因子各不相同,表明简单相关性分析不能全面客观反映各因子与Pn间的相关性,这与张文标等[18]对木荷(SchimasuperbaGardn. et Champ.)光合特性以及余红兵等[5]对花叶芦竹〔Arundodonaxvar.versicolor(Mill.) Stokes〕光合特性的研究结果一致。
本研究中,通过逐步多元回归分析得到供试4个竹种Pn值的主要影响因子,与偏相关分析结果一致。对4个竹种叶片Pn值与主要影响因子进行进一步通径分析,结果表明,各主要影响因子对4个竹种叶片Pn值的影响效应存在较复杂的相互作用,这与其他研究者对龟甲竹〔Phyllostachysheterocycla(Carr.) Mitford〕[8]、黄芩(ScutellariabaicalensisGeorgi)[19]和辣椒(CapsicumannuumLinn.)[20]等植物光合作用与环境因子相关性的研究结果类似。由通径分析结果还可以看出:供试4个竹种叶片Pn值的主要影响因子中均包含PAR和Tr值,且PAR值的影响效应较大,这与吴志庄等[7]对不同类型竹种光合特性的研究结果一致,显示PAR和Tr值均与不同竹种的Pn值具有显著或极显著相关性。
在实际的应用过程中,观赏竹的栽培利用应按照各自的光合特性和相关的影响因子,制定不同的栽培措施,因地制宜、因种制宜,保证各竹种旺盛生长,增强观赏性,使观赏竹能在园林建设中得到广泛应用。因此,应根据供试4个竹种各自的光合特性及其与相关环境因子的关系,规划合理的种植环境并制定适宜的栽培管理措施。其中,白纹阴阳竹和鼓节竹应栽培于阳光充足的区域,花秆早竹需种植在无强光直射且通风条件较好的区域,美丽箬竹则需种植在中等光照条件下。此外,根据不同竹种需要,合理灌溉和施肥,使其具有较好的生长状态和最佳的观赏性。
[1]于晓霞, 阮成江. 曼陀罗光合特性研究[J]. 植物资源与环境学报, 2011, 20(1): 40-45.
[2]李丽芳, 吴晓敏, 王立峰. 植物光合生理生态学研究进展[J]. 山西师范大学学报: 自然科学版, 2007, 21(3): 71-75.
[3]陈玉华, 宋丁全. 篌竹出笋成竹生长规律研究[J]. 南京林业大学学报: 自然科学版, 2005, 29(4): 109-112.
[4]楼扬, 孙祥, 江津凡. 黄纹竹出笋及幼竹高生长规律研究[J]. 世界竹藤通讯, 2010, 8(4): 7-10.
[5]余红兵, 曾磬, 肖润林, 等. 花叶芦竹光合特性日动态及其影响因素分析[J]. 湖南农业大学学报: 自然科学版, 2013, 39(2): 200-203.
[6]郑炳松, 金爱武, 程晓建, 等. 雷竹光合特性的研究[J]. 福建林学院学报, 2001, 21(4): 359-362.
[7]吴志庄, 杜旭华, 熊德礼, 等. 不同类型竹种光合特性的比较研究[J]. 生态环境学报, 2013, 22(9): 1523-1527.
[8]李迎春, 杨清平, 陈双林, 等. 龟甲竹光合生理特性及其与主要影响因子关系[J]. 林业科技开发, 2011, 25(2): 35-39.
[9]李在留, 辉朝茂. 珍稀竹种巨龙竹组织培养研究[J]. 林业科学, 2006, 42(2): 43-48.
[10]杨海芸, 王晓芹, 张宁, 等. 日本花叶矢竹组织培养与叶色变异研究[J]. 竹子研究汇刊, 2010, 29(4): 15-20.
[11]蒋瑶. 慈竹CBF1基因克隆及其耐寒性研究[D]. 成都: 四川农业大学林学院, 2012: 22-85.
[12]陈容, 张丽, 曹颖, 等. 低温胁迫下梁山慈竹再生植株叶绿素荧光特性和耐寒转录因子的表达[J]. 南京林业大学学报: 自然科学版, 2014, 38(4): 39-44.
[13]叶子飘, 高峻. 光响应和CO2响应新模型在丹参中的应用[J]. 西北农林科技大学学报: 自然科学版, 2009, 37(1): 129-134.
[14]许大全, 沈允钢. 植物光合作用效率的日变化[J]. 植物生理学报, 1997, 23(4): 410-416.
[15]靳甜甜, 刘国华, 胡婵娟, 等. 黄土高原常见造林树种光合蒸腾特征[J]. 生态学报, 2008, 28(11): 5758-5765.
[16]COSTE S, ROGGY J C, IMBERT P, et al. Leaf photosynthetic traits of 14 tropical rain forest species in relation to leaf nitrogen concentration and shade tolerance[J]. Tree Physiology, 2005, 25: 1127-1137.
[17]李永华, 张开明, 于红芳. 10个秋菊品种的光合特性及净光合速率与部分生理生态因子的相关性分析[J]. 植物资源与环境学报, 2012, 21(1): 70-76.
[18]张文标, 金则新, 柯世省, 等. 木荷光合特性日变化及其与环境因子相关性分析[J]. 广西植物, 2006, 26(5): 492-498.
[19]邵玺文, 韩梅, 韩忠明, 等. 不同生境条件下黄芩光合日变化与环境因子的关系[J]. 生态学报, 2009, 29(3): 1470-1477.
[20]邹学校, 马艳青, 张竹青, 等. 环境因子对辣椒光合与蒸腾特性的影响[J]. 植物资源与环境学报, 2005, 14(4): 15-20.
(责任编辑: 张明霞)
Analyses on photosynthetic characteristics of four ornamental bamboo species and its influence factors
HUANG Tao1,2, LIU Wei1, TANG Hong1, HUANG Chengqian1, HUANG Wentao1, TANG Jun1
(1. Hu’nan Forest Botanical Garden, Changsha 410116, China; 2. Central South University of Forestry and Technology, Changsha 410004, China),J.PlantResour. &Environ., 2016, 25(1): 24-33
Taking four ornamental bamboo species ofHibanobambusatranquillansf.shiroshimaH. Okamura,Bambusatuldoides‘Swolleninternode’,Phyllostachysviolascensf.viridisulcataP. X. Zhang et W. X. Huang andIndocalamusdecorusQ. H. Dai as materials tested, light response and CO2response curves and parameters, and diurnal change laws in photosynthetic and gas exchange parameters and related environmental factors related were analyzed, and the main factors affecting net photosynthetic rate (Pn) of leaf of four bamboo species were discussed by correlation analysis, stepwise multiple regression analysis and path analysis. The determination results show that with enhancing of photosynthetically active radiation intensity (PAR) or intercellular CO2concentration (Ci), Pn value of leaf of each bamboo species increases gradually, but there is a certain difference in their increasing ranges. There are great differences in light response and CO2response parameters of each bamboo species. In which, apparent quantum yield (AQY) of leaf ofH.tranquillansf.shiroshimais the lowest, CO2saturation point (CSP) and CO2compensation point (CCP) are also lower, and other parameters all are the highest, while both AQY and CCP values ofP.violascensf.viridisulcataare the highest, other parameters are the lowest generally. There are some differences in all diurnal change curves of photosynthetic and gas exchange parameters of four bamboo species. Diurnal change curve of Pn value of leaf ofI.decorusis “double-peak type”. That ofH.tranquillansf.shiroshima,B.tuldoides‘Swolleninternode’ andP.violascensf.viridisulcatais “single-peak type”, their peak values appear at 14:00, 10:00 and 12:00, respectively. Overall, diurnal change trends of stomatal conductance (Gs), Ci, Transpiration rate (Tr) and water use efficiency (WUE) of leaf of four bamboo species are similar. The analysis results show that according to total influence from high to low, main factors affecting Pn value of leaf ofH.tranquillansf.shiroshimaare PAR, Tr, air CO2concentration (Ca), Ci; those ofB.tuldoides‘Swolleninternode’ are PAR, Tr, Ca, Gs, those ofP.violascensf.viridisulcataare PAR, Tr, Gs, air temperature (Ta), Ci, and those ofI.decorusare PAR, air relative humidity (RH), Ta, Ca, Tr, Gs. The comprehensive analysis results indicate that among four bamboo species,H.tranquillansf.shiroshimahas the strongest photosynthetic ability, while it has a weak ability to adapt and use to low light, and its CO2assimilation ability is also strong with a wide adaptability;B.tuldoides‘Swolleninternode’ andI.decorusare at the middle both in utilization ability and in adaptation range of light and CO2;P.violascensf.viridisulcatais the worst both in photosynthetic ability and in CO2assimilation ability, while it has a strong ability to adapt and use to low light, and has a narrow CO2adaptation range.
ornamental bamboo species; photosynthetic characteristics; gas exchange parameters; environmental factors; correlation analysis; path analysis
10.3969/j.issn.1674-7895.2016.01.04
2015-04-23
湖南省科学技术厅项目(2011NK3019); 中央财政林业科技推广示范项目([2013]XT03)
黄滔(1977—),男,湖南长沙人,硕士研究生,高级工程师,主要从事森林生态和园林植物方面的研究。
Q945.11; S975
A
1674-7895(2016)01-0024-10
