不同产地南方红豆杉各家系幼苗生长、光合生理与株高生长节律的差异分析
2016-08-12楚秀丽尹增芳姜俊马周志春
肖 遥, 楚秀丽, 尹增芳, 姜俊马, 王 晖, 周志春
(1. 中国林业科学研究院亚热带林业研究所, 浙江 富阳 311400;2. 南京林业大学生物与环境学院, 江苏 南京 210037; 3. 浙江省淳安县富溪林场, 浙江 淳安 311700)
不同产地南方红豆杉各家系幼苗生长、光合生理与株高生长节律的差异分析
肖遥1,2, 楚秀丽1,①, 尹增芳2, 姜俊马3, 王晖3, 周志春1
(1. 中国林业科学研究院亚热带林业研究所, 浙江 富阳 311400;2. 南京林业大学生物与环境学院, 江苏 南京 210037; 3. 浙江省淳安县富溪林场, 浙江 淳安 311700)
为充分了解不同产地南方红豆杉〔Taxuswallichianavar.mairei(Lemée et H. Lév.) L. K. Fu et Nan Li〕各家系幼苗生长、光合生理及株高生长节律的差异,以产自江西分宜(FY)、浙江龙泉(LQ)、福建建瓯(JO)和福建明溪(MX)的20个家系的2年生容器苗为研究对象,对幼苗的株高和地径以及叶片的净光合速率和叶绿素含量进行比较,并对各家系的株高生长进行Logistic拟合回归分析以及拟合曲线参数和株高生长节律参数的计算和分析;在此基础上,分析了不同指标间的相关性并对供试的20个家系进行聚类分析。结果显示:不同产地间南方红豆杉的株高、地径、叶绿素含量和净光合速率均无显著差异,但这些指标在不同家系间均有极显著差异(P<0.01);各家系的株高、地径、净光合速率(Pn)以及叶绿素a(Chla)、叶绿素b(Chlb)和总叶绿素(Chl)含量的最大值较最小值分别高52.99%、29.63%、115.65%、39.81%、30.95%和37.24%。4个产地的株高生长均呈明显的“S型”曲线;各家系的最大生长速率(MGR)、线性生长速率(LGR)、线性生长总量(TLG)和线性生长持续时间(LGD)的最大值较最小值分别高60.00%、61.54%、63.73%和22.73%;从线性生长始期(t1)和线性生长终期(t2)来看,各家系的速生期为5月中旬或下旬至8月上旬。幼苗的株高与MGR、TLG、LGR、地径、Pn、Chlb和Chl等指标均极显著正相关,并与Chla显著正相关,而地径与MGR、TLG、LGR、Pn和Chlb等指标显著正相关,但株高和地径与LGD均无显著相关性。聚类分析结果表明:在欧氏距离10处,供试的20个家系被分成4个类型,其中,第Ⅳ类的6个家系的幼苗生长最佳且生长潜力最高。研究结果表明:不同产地南方红豆杉各家系的遗传变异丰富,且株高和地径生长主要受生长速率的影响;总体来看,家系LQ09、LQ11、FY21、FY26、MX12和MX18的幼苗生长潜力较高,可作为南方红豆杉的优良家系进行推广。
南方红豆杉; 株高生长节律; 光合生理; Logistic拟合回归分析; 相关性分析; 聚类分析
南方红豆杉〔Taxuswallichianavar.mairei(Lemée et H. Lév.) L. K. Fu et Nan Li〕是中国境内分布最广泛的红豆杉属(TaxusLinn.)植物,不仅能够用于提取抗癌成分紫杉醇,而且还是珍贵的用材树种[1-2]。南方红豆杉虽然是珍稀濒危植物,但其部分种群仍具有较高的遗传多样性[3-5];张蕊等[6]和王艺等[7]的研究结果均显示南方红豆杉的不同种源间存在显著的生长差异;焦月玲等[8]也认为南方红豆杉不同种源间的光合特性存在较大差异。可见,南方红豆杉具有丰富的遗传变异,充分利用南方红豆杉的这一特性对提高其栽培和育种效果有益。
生长节律对林木的存活以及种间和种内竞争具有重要作用[9],与林木对有机物的积累和消耗也有一定的联系[10]。对于经济树种而言,可以通过其生长节律准确判断副产物的最佳收获时期[11]。通常情况下,研究者多采用数学模型和数学方法来分析林木的生长节律[12-16],并且,多数树种的高生长节律均表现出“慢—快—慢”的“S型”生长趋势,但不同树种间或同一树种不同种源间林木的生长节律的差异却较大。随研究技术的不断发展,对林木生长节律的研究已深入至分子水平。如Avia等[17]通过基因表达技术探讨了欧洲赤松(PinussylvestrisLinn.)生长节律的发生机制,认为PsFTL2基因可调控欧洲赤松芽的生长,且该基因的表达量在不同产地的欧洲赤松种群间存在较大差异。
目前,南方红豆杉的野生资源相对匮乏,加之其在自然条件下生长缓慢,因此亟待选育出一批速生优良的南方红豆杉家系以提高其产量。为了实现这一目标,必须充分了解南方红豆杉的株高生长节律,尤其是不同种源和不同家系的株高生长节律的变化规律,但目前关于南方红豆杉家系水平的相关研究尚未见报道。
鉴于此,作者对来源于江西分宜、浙江龙泉、福建建瓯和福建明溪4个产地的南方红豆杉20个家系幼苗的株高、地径以及叶片净光合速率和叶绿素含量的差异进行了比较和巢氏方差分析,对各家系株高生长节律进行Logistic拟合回归分析及株高生长节律参数分析,并研究了株高生长节律参数与光合生理指标的相关性;在此基础上,依据欧氏距离对供试的20个家系进行聚类分析。以期明确南方红豆杉不同家系的株高生长节律,为南方红豆杉优良家系的筛选提供基础研究数据。
1 材料和方法
1.1材料
供试的南方红豆杉均种植于浙江省淳安县富溪林场龙门里苗圃内。该苗圃的具体地理坐标为北纬29°37′、东经119°01′;地处中北亚热带过渡季风气候区,四季分明,阳光充足,气候温和湿润,年均温17 ℃,年降水量1 430 mm,无霜期263 d;地形以山地为主,土壤为砂壤土,肥力中等。
从苗圃内的55个家系中选取来源于江西分宜(FY)、浙江龙泉(LQ)、福建建瓯(JO)和福建明溪(MX)4个产地的南方红豆杉2年生容器苗供试。每个产地选择5个代表性家系,编号分别为FY02、FY17、FY21、FY23、FY26、LQ02、LQ03、LQ05、LQ09、LQ11、JO12、JO13、JO14、JO17、JO22、MX07、MX12、MX18、MX20和MX21,共20个家系。
1.2方法
1.2.1幼苗移栽及管理采用完全随机区组设计,从各家系中选择株高11.00~23.00 cm、地径2.78~4.67 mm的容器苗,于2014年3月底移栽到直径15 cm、高20 cm的无纺布袋中, 栽培基质为V(泥炭)∶V(谷壳)∶V(黄泥)=4∶3∶3的混合基质,每盆1株。共5个小区,每个小区各家系均为10盆。苗期的除草、施肥、灌溉及病虫害防治等措施与常规容器苗管理相同,夏季高温时适当遮阳。
1.2.2指标测定于2014年4月10日随机选择3个小区,测定每个家系所有植株的株高,此后每月10日对这3个小区内所有植株的株高进行测定,直至11月10日为止。2014年11月10日,测定上述3个小区内所有植株的地径。结果取平均值。
在上述3个小区中随机选择1个小区,从每个家系选择3株代表性植株,于2014年9月27日至29日(晴朗天气)的上午9:30至10:30使用LI-6400便携式光合仪(美国LI-COR公司)测定植株冠层中部的成熟功能叶片中部的净光合速率,每株检测3枚叶片,结果取平均值。
于2014年9月29日,在上述3个小区中随机选择1个小区,从每个家系中选择3~5株代表性植株,采集冠层中部同方位的长势良好、叶色正常且无损伤的成熟功能叶片,混合后置于冰盒内带回实验室。将叶片剪碎并充分混匀,每个家系分别称取3份碎叶样品,采用分光光度法[18]测定叶片中的叶绿素含量,结果取平均值。
1.3数据处理和统计分析
采用Logistic拟合回归分析法拟合各家系的株高生长节律,拟合方程为y=k/(1+a·e-bt)。式中,y为株高生长量;t为生长时间;a和b为待定系数;k为既定条件下株高生长可能达到的极限值。参照朱仁海等[19]的方法计算南方红豆杉各家系幼苗的线性生长始期(t1)和线性生长终期(t2),再结合上述拟合方程计算各家系幼苗的最大生长速率(MGR)、线性生长速率(LGR)、线性生长总量(TLG)和线性生长持续时间(LGD)。
采用DPS 7.05软件对南方红豆杉产地间和产地内不同家系间的幼苗株高和地径、叶片的净光合速率和叶绿素含量进行巢式方差分析;采用SPSS 20.0软件进行Logistic拟合回归分析、相关性分析和聚类分析。文中各类图表制作和数据计算均采用EXCEL 2007软件完成。
2 结果和分析
2.1幼苗生长及叶片部分光合生理指标的差异分析
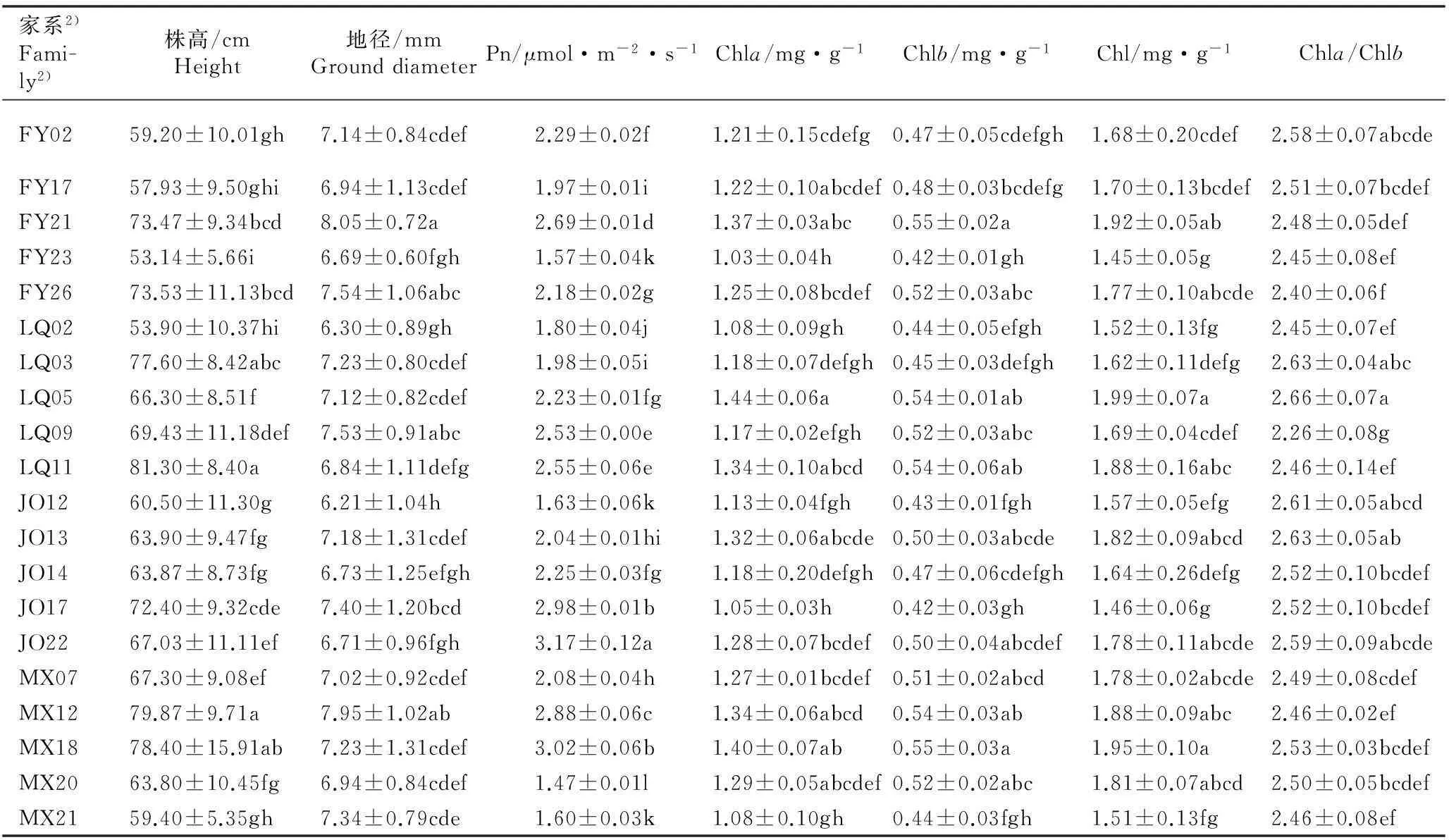
不同产地南方红豆杉各家系的幼苗株高和地径以及叶片的净光合速率和叶绿素含量的测定结果见表1。
2.1.1幼苗株高和地径的差异分析表1结果显示:不同产地南方红豆杉各家系的幼苗株高和地径分别为53.14~81.30 cm和6.21~8.05 mm,且差异显著。其中,产自浙江龙泉的家系LQ11的株高最高,产自江西分宜的家系FY23的株高最低,且前者较后者高52.99%;产自江西分宜的家系FY21的地径最大,产自福建建瓯的家系JO12的地径最小,且前者较后者高29.63%。
2.1.2叶片净光合速率的差异分析由表1可见:不同产地南方红豆杉各家系的叶片净光合速率差异明显。各家系叶片净光合速率的变幅为1.47~3.17 μmol·m-2·s-1,其中,产自福建建瓯的家系JO22的叶片净光合速率最大,产自福建明溪的家系MX20的叶片净光合速率最小,前者较后者高115.65%。
2.1.3叶片叶绿素含量的差异分析由表1还可以看出: 南方红豆杉各家系叶片的叶绿素含量也有明显差异。各家系叶片的叶绿素a和总叶绿素含量分别为1.03~1.44和1.45~1.99 mg·g-1,并且均以产自浙江龙泉的家系LQ05为最高、产自江西分宜的家系FY23为最低,前者分别较后者高39.81%和37.24%。各家系叶片的叶绿素b含量为0.42~0.55 mg·g-1,并且,以产自福建明溪的家系MX18为最高、产自福建建瓯的家系JO17和产自江西分宜的家系FY23为最低,前者较后两者高30.95%。各家系叶片的叶绿素a和叶绿素b的含量比值为2.26~2.66,最高值和最低值分别为产自浙江龙泉的家系LQ05和LQ09,且前者较后者高17.70%。

家系2)Fami-ly2)株高/cmHeight地径/mmGrounddiameterPn/μmol·m-2·s-1Chla/mg·g-1Chlb/mg·g-1Chl/mg·g-1Chla/ChlbFY0259.20±10.01gh7.14±0.84cdef2.29±0.02f1.21±0.15cdefg0.47±0.05cdefgh1.68±0.20cdef2.58±0.07abcdeFY1757.93±9.50ghi6.94±1.13cdef1.97±0.01i1.22±0.10abcdef0.48±0.03bcdefg1.70±0.13bcdef2.51±0.07bcdefFY2173.47±9.34bcd8.05±0.72a2.69±0.01d1.37±0.03abc0.55±0.02a1.92±0.05ab2.48±0.05defFY2353.14±5.66i6.69±0.60fgh1.57±0.04k1.03±0.04h0.42±0.01gh1.45±0.05g2.45±0.08efFY2673.53±11.13bcd7.54±1.06abc2.18±0.02g1.25±0.08bcdef0.52±0.03abc1.77±0.10abcde2.40±0.06fLQ0253.90±10.37hi6.30±0.89gh1.80±0.04j1.08±0.09gh0.44±0.05efgh1.52±0.13fg2.45±0.07efLQ0377.60±8.42abc7.23±0.80cdef1.98±0.05i1.18±0.07defgh0.45±0.03defgh1.62±0.11defg2.63±0.04abcLQ0566.30±8.51f7.12±0.82cdef2.23±0.01fg1.44±0.06a0.54±0.01ab1.99±0.07a2.66±0.07aLQ0969.43±11.18def7.53±0.91abc2.53±0.00e1.17±0.02efgh0.52±0.03abc1.69±0.04cdef2.26±0.08gLQ1181.30±8.40a6.84±1.11defg2.55±0.06e1.34±0.10abcd0.54±0.06ab1.88±0.16abc2.46±0.14efJO1260.50±11.30g6.21±1.04h1.63±0.06k1.13±0.04fgh0.43±0.01fgh1.57±0.05efg2.61±0.05abcdJO1363.90±9.47fg7.18±1.31cdef2.04±0.01hi1.32±0.06abcde0.50±0.03abcde1.82±0.09abcd2.63±0.05abJO1463.87±8.73fg6.73±1.25efgh2.25±0.03fg1.18±0.20defgh0.47±0.06cdefgh1.64±0.26defg2.52±0.10bcdefJO1772.40±9.32cde7.40±1.20bcd2.98±0.01b1.05±0.03h0.42±0.03gh1.46±0.06g2.52±0.10bcdefJO2267.03±11.11ef6.71±0.96fgh3.17±0.12a1.28±0.07bcdef0.50±0.04abcdef1.78±0.11abcde2.59±0.09abcdeMX0767.30±9.08ef7.02±0.92cdef2.08±0.04h1.27±0.01bcdef0.51±0.02abcd1.78±0.02abcde2.49±0.08cdefMX1279.87±9.71a7.95±1.02ab2.88±0.06c1.34±0.06abcd0.54±0.03ab1.88±0.09abc2.46±0.02efMX1878.40±15.91ab7.23±1.31cdef3.02±0.06b1.40±0.07ab0.55±0.03a1.95±0.10a2.53±0.03bcdefMX2063.80±10.45fg6.94±0.84cdef1.47±0.01l1.29±0.05abcdef0.52±0.02abc1.81±0.07abcd2.50±0.05bcdefMX2159.40±5.35gh7.34±0.79cde1.60±0.03k1.08±0.10gh0.44±0.03fgh1.51±0.13fg2.46±0.08ef
1)Pn: 净光合速率 Net photosynthetic rate; Chla: 叶绿素a含量 Content of chlorophylla; Chlb: 叶绿素b含量 Content of chlorophyllb; Chl: 总叶绿素含量 Content of total chlorophyll; Chla/Chlb: 叶绿素a与叶绿素b的含量比值Content ratio of chlorophyllato chlorophyllb. 同列中不同的小写字母表示差异显著(P<0.05) Different small letters in the same column indicate the significant difference (P<0.05).
2)FY: 江西分宜 Fenyi of Jiangxi; LQ: 浙江龙泉 Longquan of Zhejiang; JO: 福建建瓯 Jian’ou of Fujian; MX: 福建明溪 Mingxi of Fujian.
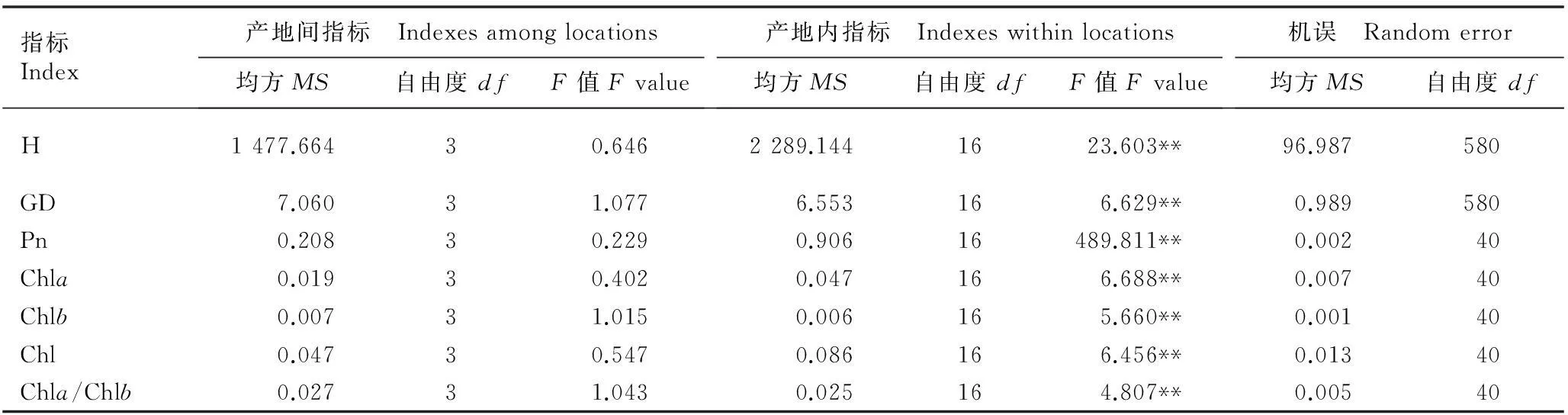
巢式方差分析结果(表2)显示:不同产地间南方红豆杉幼苗的株高和地径、叶片的净光合速率和叶绿素含量均无显著差异,但产地内不同家系间各指标均有极显著差异(P<0.01)。
2.2不同家系幼苗株高生长节律的差异分析
2.2.1Logistic拟合回归分析对不同产地南方红豆杉各家系幼苗的株高生长进行Logistic拟合回归分析,计算出的拟合曲线参数见表3。
分析结果显示:R2值为0.990~0.997,表明获得的各家系的拟合曲线的拟合度良好。4个产地中,产自浙江龙泉和福建建瓯的南方红豆杉各家系幼苗株高生长的平均k值均较高(分别为50.792和48.164)且二者间差异较小;产自福建明溪的南方红豆杉幼苗各家系株高生长的平均k值最大(51.037),而产自江西分宜的各家系幼苗株高生长的平均k值最小(45.220),且前者较后者高12.86%。各家系间的k值差异相对较大,其中产自浙江龙泉的家系LQ11的k值最大(60.828),产自江西分宜的家系FY17的k值最小(37.148),前者较后者高63.75%。产自浙江龙泉和福建明溪的南方红豆杉各家系幼苗株高生长的平均a值也较高,并以产自福建明溪的各家系为最高(18.576);产自江西分宜和福建建瓯的各家系幼苗株高生长的平均a值均较低,并以产自福建建瓯的各家系幼苗株高生长的平均a值为最低(15.186)。4个产地各家系幼苗株高生长的平均b值基本接近,差异不明显。
表2不同产地南方红豆杉各家系幼苗的株高和地径以及叶片的净光合速率和叶绿素含量的巢式方差分析结果1)
Table 2Result of nested variance analysis on height and ground diameter of seedlings, net photosynthetic rate and chlorophyll content in leaf of each family ofTaxuswallichianavar.mairei(Lemée et H. Lév.) L. K. Fu et Nan Li from different locations1)
指标Index产地间指标 Indexesamonglocations均方MS自由度dfF值Fvalue产地内指标 Indexeswithinlocations均方MS自由度dfF值Fvalue机误 Randomerror均方MS自由度dfH1477.66430.6462289.1441623.603**96.987580GD7.06031.0776.553166.629**0.989580Pn0.20830.2290.90616489.811**0.00240Chla0.01930.4020.047166.688**0.00740Chlb0.00731.0150.006165.660**0.00140Chl0.04730.5470.086166.456**0.01340Chla/Chlb0.02731.0430.025164.807**0.00540
1)H: 株高 Height; GD: 地径 Ground diameter; Pn: 净光合速率 Net photosynthetic rate; Chla: 叶绿素a含量 Content of chlorophylla; Chlb: 叶绿素b含量 Content of chlorophyllb; Chl: 总叶绿素含量 Content of total chlorophyll; Chla/Chlb: 叶绿素a与叶绿素b的含量比值Content ratio of chlorophyllato chlorophyllb. **:P<0.01.
表3不同产地南方红豆杉各家系幼苗株高生长的拟合曲线参数
Table 3Parameters of fitting curve of height growth of seedlings of each family ofTaxuswallichianavar.mairei(Lemée et H. Lév.) L. K. Fu et Nan Li from different locations

家系1)Fami-ly1)kabR2FY0240.78816.0400.0350.995FY2654.35319.7230.0320.994FY2154.42417.8830.0310.996FY2339.38516.1890.0330.992FY1737.14818.9690.0340.996均值Average45.22017.7610.0330.995LQ1160.82815.3730.0320.995LQ0952.70615.5350.0310.994LQ0354.59624.0100.0350.995LQ0237.97218.6490.0310.996LQ0547.86017.0540.0320.996均值Average50.79218.1240.0320.995JO1248.36911.3050.0290.990JO1748.52519.4480.0330.996JO2247.14719.7700.0330.997JO1349.74112.5900.0290.992JO1447.03812.8170.0290.993均值Average48.16415.1860.0310.994MX1855.34219.7970.0330.993MX0749.25719.3440.0330.994MX1258.78213.4350.0290.993MX2044.46026.8130.0320.997MX2147.34213.4890.0350.994均值Average51.03718.5760.0320.994
1)FY: 江西分宜 Fenyi of Jiangxi; LQ: 浙江龙泉 Longquan of Zhejiang; JO: 福建建瓯 Jian’ou of Fujian; MX: 福建明溪 Mingxi of Fujian.
由Logistic拟合曲线(图1)可以看出:4个产地南方红豆杉的株高生长均呈明显的“S型”曲线,其中,产自浙江龙泉和福建明溪的南方红豆杉幼苗的株高生长拟合曲线基本重合。

—○—: 福建明溪 Mingxi of Fujian; —▲—: 浙江龙泉 Longquan of Zhejiang; —■—: 福建建瓯 Jian’ou of Fujian; —◆—: 江西分宜 Fenyi of Jiangxi.
生长时间从4月10日开始计算 Growth time is accounted from April 10.
图1不同产地南方红豆杉幼苗株高生长的Logistic拟合曲线
Fig. 1Logistic fitting curves of height growth of seedlings ofTaxuswallichianavar.mairei(Lemée et H. Lév.) L. K. Fu et Nan Li from different locations
2.2.2株高生长节律参数分析不同产地南方红豆杉各家系幼苗株高生长节律参数见表4。
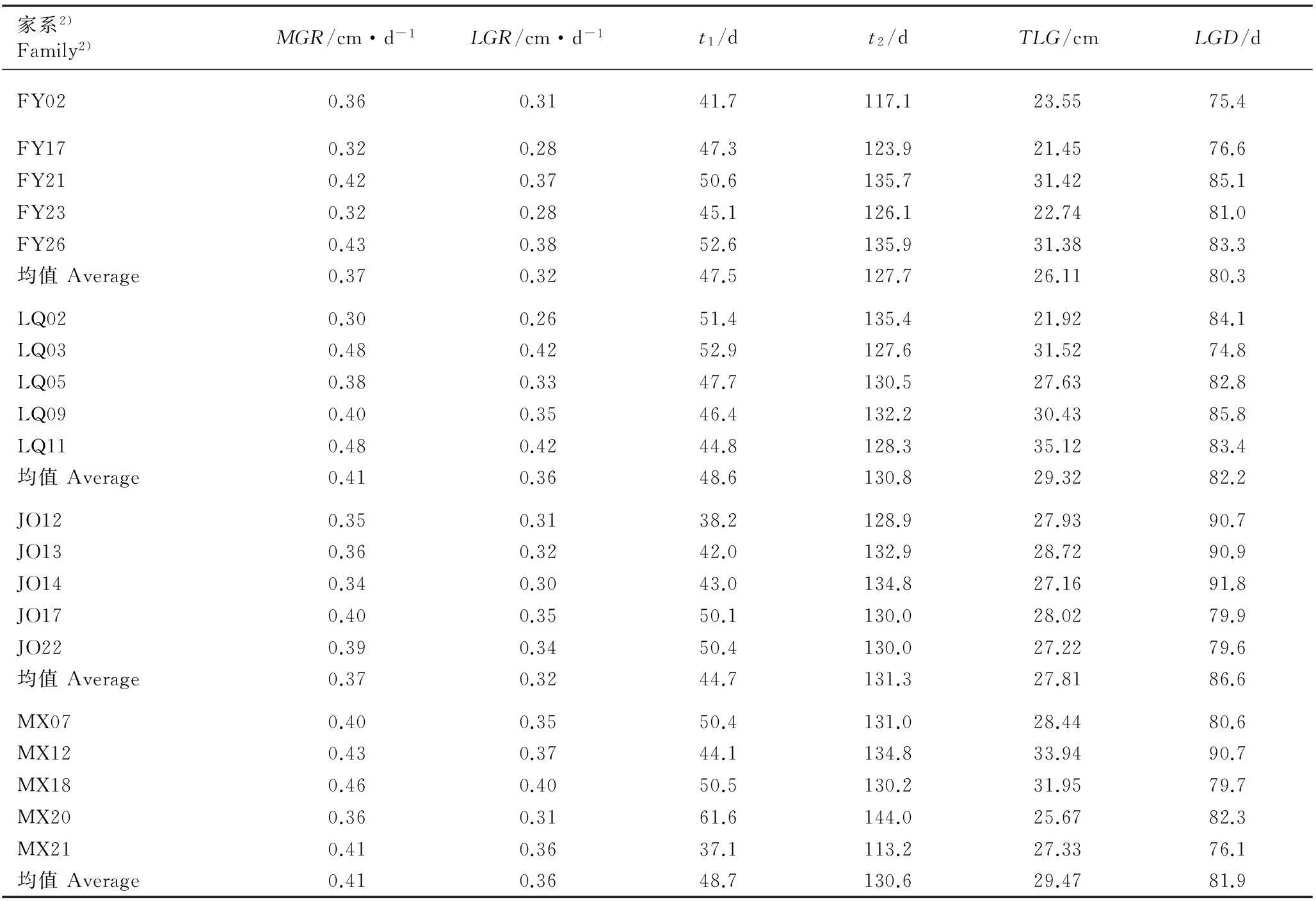
结果显示:供试4个产地南方红豆杉20个家系的幼苗株高生长节律参数总体上差异较大,最大生长速率(MGR)、线性生长速率(LGR)、线性生长始期(t1)、线性生长终期(t2)、线性生长总量(TLG)和线性生长持续时间(LGD)分别为0.30~0.48 cm·d-1、0.26~0.42 cm·d-1、37.1~52.9 d、113.2~144.0 d、21.45~35.12 cm和74.8~91.8 d,其最大值分别较最小值高60.00%、 61.54%、 42.59%、 27.21%、 63.73%和22.73%。
以4月10日为生长起始点,从各家系的t1和t2值可以看出,供试的南方红豆杉各家系幼苗均在5月中旬或下旬进入速生期,其速生期大约可持续2个半月至3个月,并在8月上旬结束速生状态,但产自福建建瓯的家系JO12、JO13和JO14的线性生长持续时间均超过3个月。
由表4还可见:产自江西分宜和福建建瓯的各家系幼苗的MGR、LGR、t1和TLG的平均值均小于产自浙江龙泉和福建明溪的各家系,产自浙江龙泉和福建建瓯的各家系幼苗的t2和LGD平均值则高于产自江西分宜和福建明溪的各家系,说明生长速率对南方红豆杉各家系幼苗生长的影响效应比线性生长持续时间更大。
表4不同产地南方红豆杉各家系幼苗的株高生长节律参数1)
Table 4Parameters of height growth rhythm of seedlings of each family ofTaxuswallichianavar.mairei(Lemée et H. Lév.) L. K. Fu et Nan Li from different locations1)
家系2)Family2)MGR/cm·d-1LGR/cm·d-1t1/dt2/dTLG/cmLGD/dFY020.360.3141.7117.123.5575.4FY170.320.2847.3123.921.4576.6FY210.420.3750.6135.731.4285.1FY230.320.2845.1126.122.7481.0FY260.430.3852.6135.931.3883.3均值Average0.370.3247.5127.726.1180.3LQ020.300.2651.4135.421.9284.1LQ030.480.4252.9127.631.5274.8LQ050.380.3347.7130.527.6382.8LQ090.400.3546.4132.230.4385.8LQ110.480.4244.8128.335.1283.4均值Average0.410.3648.6130.829.3282.2JO120.350.3138.2128.927.9390.7JO130.360.3242.0132.928.7290.9JO140.340.3043.0134.827.1691.8JO170.400.3550.1130.028.0279.9JO220.390.3450.4130.027.2279.6均值Average0.370.3244.7131.327.8186.6MX070.400.3550.4131.028.4480.6MX120.430.3744.1134.833.9490.7MX180.460.4050.5130.231.9579.7MX200.360.3161.6144.025.6782.3MX210.410.3637.1113.227.3376.1均值Average0.410.3648.7130.629.4781.9
1)MGR: 最大生长速率 The maximum growth rate;LGR: 线性生长速率 Linear growth rate;t1: 线性生长始期 The beginning day of linear growth;t2: 线性生长终期 The final day of linear growth;TLG: 线性生长总量 Total linear growth;LGD: 线性生长持续时间 Linear growth duration.
2)FY: 江西分宜 Fenyi of Jiangxi; LQ: 浙江龙泉 Longquan of Zhejiang; JO: 福建建瓯 Jian’ou of Fujian; MX: 福建明溪 Mingxi of Fujian.
2.3幼苗株高生长节律参数、生长指标和叶片部分光合生理指标间的相关性分析
南方红豆杉的幼苗株高生长节律参数、幼苗株高和地径、叶片净光合速率和叶绿素含量的相关性分析结果见表5。
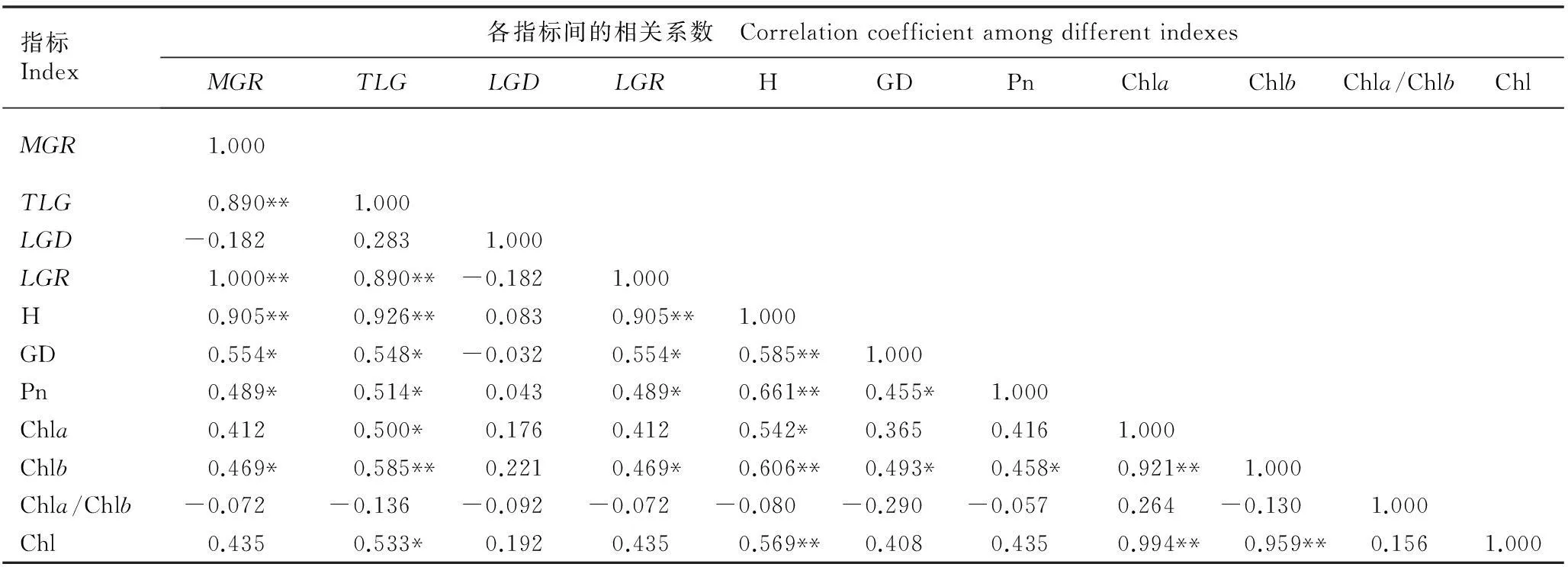
相关性分析结果表明:幼苗的最大生长速率(MGR)与线性生长总量(TLG)、线性生长速率(LGR)和株高(H)均呈极显著正相关(P<0.01),与地径(GD)、净光合速率(Pn)和叶绿素b含量(Chlb)均呈显著正相关(P<0.05);TLG与LGR、H和Chlb均呈极显著正相关,与GD、Pn、叶绿素a含量(Chla)和总叶绿素含量(Chl)均呈显著正相关;线性生长持续时间(LGD)与各指标间均无显著相关性;LGR与H呈极显著正相关,与GD、Pn和Chlb均呈显著正相关;H与
表5南方红豆杉幼苗株高生长节律参数、株高和地径、叶片净光合速率和叶绿素含量间的相关系数1)
Table 5Correlation coefficients among height growth rhythm parameters, height and ground diameter of seedlings, net photosynthetic rate and chlorophyll content in leaf ofTaxuswallichianavar.mairei(Lemée et H. Lév.) L. K. Fu et Nan Li1)
指标Index各指标间的相关系数 CorrelationcoefficientamongdifferentindexesMGRTLGLGDLGRHGDPnChlaChlbChla/ChlbChlMGR1.000TLG0.890**1.000LGD-0.1820.2831.000LGR1.000**0.890**-0.1821.000H0.905**0.926**0.0830.905**1.000GD0.554*0.548*-0.0320.554*0.585**1.000Pn0.489*0.514*0.0430.489*0.661**0.455*1.000Chla0.4120.500*0.1760.4120.542*0.3650.4161.000Chlb0.469*0.585**0.2210.469*0.606**0.493*0.458*0.921**1.000Chla/Chlb-0.072-0.136-0.092-0.072-0.080-0.290-0.0570.264-0.1301.000Chl0.4350.533*0.1920.4350.569**0.4080.4350.994**0.959**0.1561.000
1)MGR: 最大生长速率 The maximum growth rate;TLG: 线性生长总量 Total linear growth;LGD: 线性生长持续时间 Linear growth duration;LGR: 线性生长速率 Linear growth rate; H: 株高 Height; GD: 地径 Ground diameter; Pn: 净光合速率 Net photosynthetic rate; Chla: 叶绿素a含量 Content of chlorophylla; Chlb: 叶绿素b含量 Content of chlorophyllb; Chla/Chlb: 叶绿素a与叶绿素b的含量比值Content ratio of chlorophyllato chlorophyllb; Chl: 总叶绿素含量 Content of total chlorophyll. *:P<0.05; **:P<0.01.
GD、Pn、Chlb和Chla/Chlb均呈极显著正相关,与Chla呈显著正相关;GD与Pn,以及Chlb与GD和Pn均呈显著正相关;Chla与Chlb以及Chl与Chla和Chlb均呈极显著正相关。
2.4不同产地南方红豆杉各家系的聚类分析
依据欧氏距离对不同产地南方红豆杉20个家系进行聚类分析,结果见图2。
由图2可以看出:在欧氏距离10处,供试的20个南方红豆杉家系被分成4个类型,其中,第Ⅰ类包括FY23、LQ02、JO12和JO14共4个家系;第Ⅱ类包括JO17、MX21和LQ03共3个家系;第Ⅲ类包括FY02、FY17、LQ05、JO13、JO22、MX07和MX20共7个家系;第Ⅳ类包括LQ09、LQ11、FY21、FY26、MX12和MX18共6个家系。
对南方红豆杉上述4个类型家系幼苗的株高和地径、株高生长节律参数、叶片净光合速率和叶绿素含量进行综合比较,结果见表6。
综合比较结果表明:第Ⅰ类家系的南方红豆杉幼苗生长最差,其植株相对矮小和纤细,最大生长速率(MGR)、线性生长速率(LGR)和线性生长总量(TLG)均较低,并且叶片的净光合速率(Pn)和叶绿素含量也相对较低;第Ⅱ类家系的南方红豆杉幼苗生长较好,其MGR和LGR均较高,Pn也较高,而叶绿素含量却最低;第Ⅲ类家系的南方红豆杉幼苗生长较差,其MGR、苗高、地径和Pn均相对较弱,但叶绿素含量却较高;第Ⅳ类家系的南方红豆杉幼苗生长最佳,其苗高、地径、MGR、LGR、TLG、Pn和叶绿素含量均最高。

FY: 江西分宜 Fenyi of Jiangxi; LQ: 浙江龙泉 Longquan of Zhejiang; JO: 福建建瓯 Jian’ou of Fujian; MX: 福建明溪 Mingxi of Fujian.
图2不同产地南方红豆杉各家系的聚类图
Fig. 2Dendrogram of each family ofTaxuswallichianavar.mairei(Lemée et H. Lév.) L. K. Fu et Nan Li from different locations
表6不同产地南方红豆杉4个类型家系的幼苗株高、地径和部分株高生长节律参数以及叶片光合生理指标的比较1)
Table 6Comparison on height, ground diameter and part height growth rhythm parameters of seedlings and photosynthetic physiological indexes in leaf of four type families ofTaxuswallichianavar.mairei(Lemée et H. Lév.) L. K. Fu et Nan Li from different locations1)

类型TypeH/cmGD/mmMGR/cm·d-1LGR/cm·d-1TLG/cmLGD/dPn/μmol·m-2·s-1含量/mg·g-1 ContentChlaChlbChlChla/ChlbⅠ57.856.480.330.2924.9486.901.811.110.441.552.51Ⅱ69.87.320.430.3828.9676.932.191.100.431.532.54Ⅲ63.647.010.370.3226.1081.172.181.290.501.792.57Ⅳ76.007.520.440.3832.3784.672.641.310.541.852.43
1)H: 苗高 Height; GD: 地径 Ground diameter;MGR: 最大生长速率 The maximum growth rate;LGR: 线性生长速率 Linear growth rate;TLG: 线性生长总量 Total linear growth;LGD: 线性生长持续时间 Linear growth duration; Pn: 净光合速率 Net photosynthetic rate; Chla: 叶绿素aChlorophylla; Chlb: 叶绿素bChlorophyllb; Chl: 总叶绿素Total chlorophyll; Chla/Chlb: 叶绿素a与叶绿素b的含量比值Content ratio of chlorophyllato chlorophyllb.
3 讨论和结论
佘诚棋等[20]认为林木变异存在多种模式,并且不同性状变异在种源和家系水平上差异较大。本研究中,不同产地南方红豆杉各家系幼苗的株高和地径、叶片的净光合速率和叶绿素含量均存在一定差异,并且各指标在同一产地内不同家系间差异极显著,而在不同产地间则无显著差异。这一研究结果与焦月玲等[21]得出的“南方红豆杉的生长变异在产地间较大”的研究结果明显不同,推测这可能是由于本研究涉及的各产地的家系数量相对较少,不能完全代表各产地的所有家系。而麻栎(QuercusacutissimaCarruth.)[22]和青钱柳〔Cyclocaryapaliurus(Batal.) Iljinsk.〕[20]的表型性状变异主要来源于种源内或家系间,本研究结论与这一结论相似。
光合作用与植物的生长和产量密切相关,培育高光效林木已成为当今育种专家关注的焦点[23-27]。同一产地内不同家系间南方红豆杉的净光合速率存在极显著差异,为选育高光效的南方红豆杉家系提供了遗传改良的基础。南方红豆杉各家系的叶片叶绿素a与叶绿素b的含量比值均为2.5左右,而阳生植物的该比值通常在3.0左右[28],说明南方红豆杉具有喜阴的生物学特性。相关研究结果也显示红豆杉属植物适宜生长在荫蔽环境中[29-30]。因此,建议在南方红豆杉的培育过程中采取适当的遮阳措施。
本研究采用Logistic拟合回归分析法对不同产地南方红豆杉各家系的株高生长节律进行拟合分析,结果表明其株高生长呈现“S型”曲线,符合树木生长的一般规律[31-32]。并且,南方红豆杉的线性生长阶段(即速生期)为5月中旬或下旬至8月上旬。在此时段内,气温相对较高,光照也较强,及时实施浇水和遮阳等管理措施不仅能够减轻幼苗受到的胁迫程度,而且还能够使幼苗的生长效率达到最大。南方红豆杉各家系最大生长速率(MGR)、线性生长速率(LGR)和线性生长总量(TLG)的最大值较最小值的增幅均超过60%,并且产自浙江龙泉和福建明溪的南方红豆杉幼苗的MGR和LGR均最高。Petterle等[33]认为地理位置的光周期是季节变化的主要指标,能够调控植物的生长节律。据此认为,浙江龙泉和福建明溪的光照条件和气候环境可能更适宜南方红豆杉生长。
相关性分析结果表明:南方红豆杉TLG与MGR和LGR极显著正相关,与线性生长持续时间(LGD)无显著相关性,说明南方红豆杉的株高生长量主要取决于生长速率,而与生长时间无关,这一研究结果与张萍等[14]对木荷(SchimasuperbaGardn. et Champ.)株高生长节律的相关研究结果相似。净光合速率(Pn)与MGR和LGR均显著正相关,说明光合作用能够有效加快南方红豆杉的生长速率;株高(H)与地径(GD)以及二者与TLG、MGR、LGR、Pn和叶绿素b含量均存在显著或极显著正相关,可见南方红豆杉幼苗的株高和地径生长同样取决于其生长速率。从LGD与GD呈负相关可以看出线性生长持续时间过长不利于南方红豆杉幼苗地径的生长。
聚类分析结果表明:供试的20个家系被分成4个类型,以第Ⅳ类家系的幼苗株高和地径生长最佳,并且其LGR、MGR、TLG、Pn和叶绿素含量均最高,具有较强的后期生长潜力。据此筛选出南方红豆杉的6个优良家系,分别为产自浙江龙泉的家系LQ09和LQ11、产自江西分宜的家系FY21和FY26及产自福建明溪的家系MX12和MX18。并且,在选择家系时建议重点关注线性生长速率。
[1]洪伟, 王新功, 吴承祯, 等. 濒危植物南方红豆杉种群生命表及谱分析[J]. 应用生态学报, 2004, 15(6): 1109-1112.
[2]李志辉, 李柏海, 祁承经, 等. 我国南方珍贵用材树种资源的重要性及其发展策略[J]. 中南林业科技大学学报, 2012, 32(11): 1-8.
[3]张蕊, 周志春, 金国庆, 等. 南方红豆杉种源遗传多样性和遗传分化[J]. 林业科学, 2009, 45(1): 50-56.
[4]ZHANG D Q, ZHOU N. Genetic diversity and population structure of the endangered coniferTaxuswallichianavar.mairei(Taxaceae) revealed by Simple Sequence Repeat (SSR) markers[J]. Biochemical Systematics and Ecology, 2013, 49: 107-114.
[5]李乃伟, 贺善安, 束晓春, 等. 基于ISSR标记的南方红豆杉野生种群和迁地保护种群的遗传多样性和遗传结构分析[J]. 植物资源与环境学报, 2011, 20(1): 25-30.
[6]张蕊, 周志春, 余能健, 等. 不同种源南方红豆杉幼林生长和紫杉醇含量的研究[J]. 林业科学研究, 2011, 24(1): 56-62.
[7]王艺, 张蕊, 冯建国, 等. 不同种源南方红豆杉生长差异分析及早期速生优良种源筛选[J]. 植物资源与环境学报, 2012, 21(4): 41-47.
[8]焦月玲, 周志春, 李因刚, 等. 南方红豆杉种源光合特性差异及光环境效应[J]. 林业科学研究, 2007, 20(5): 731-735.
[9]LUKKARINEN A J, RUOTSALAINEN S, PELTOLA H, et al. Annual growth rhythm ofLarixsibiricaandLarixgmeliniiprovenances in a field trial in southern Finland[J]. Scandinavian Journal of Forest Research, 2013, 28: 518-532.
[10]SHEFFIELD M C P, GAGNON J L, JACK S B, et al. Phenological patterns of mature longleaf pine (PinuspalustrisMiller) under two different soil moisture regimes[J]. Forest Ecology and Management, 2003, 179: 157-167.
[11]QI D L, YANG C, XIE G S, et al. Preliminary study on seedling growth rhythm and grey correlation analysis of rubber (Heveabrasiliensis) seedlings in Danzhou District, Hainan[J]. American Journal of Plant Sciences, 2014, 5: 3866-3872.
[12]魏润鹏. 桉树无性系顶梢伸长节律及其在良种选择中的潜在作用[J]. 林业科学, 2012, 48(7): 153-160.
[13]曹健康, 方乐金. 光皮桦幼林生长节律的观察研究[J]. 林业科学研究, 2006, 19(3): 404-407.
[14]张萍, 周志春, 金国庆, 等. 木荷种源苗高生长参数变异研究[J]. 林业科学研究, 2006, 19(1): 61-65.
[15]李秋荔, 黄寿先, 李志先, 等. 中国马褂木不同种源苗期生长规律研究[J]. 广西植物, 2012, 32(3): 355-361.
[16]郑益兴, 冯永刚, 彭兴民, 等. 印楝1年生苗木生长节律与数量分级标准[J]. 南京林业大学学报: 自然科学版, 2008, 32(3): 25-30.
FTL2 expression with growth rhythm in Scots pine (Pinussylvestris)[J]. New Phytologist, 2014, 204: 159-170.
[18]李合生. 植物生理生化实验原理和技术[M]. 北京: 高等教育出版社, 2000: 134-137.
[19]朱仁海, 杨琪瑜, 沈文瑛. 统计分析方法[M]. 北京: 中国林业出版社, 1990.
[20]佘诚棋, 杨万霞, 方升佐, 等. 青钱柳种源间苗期性状变异分析[J]. 南京林业大学学报: 自然科学版, 2010, 34(1): 34-38.
[21]焦月玲, 周志春, 余能健, 等. 南方红豆杉苗木性状种源分化和育苗环境对苗木生长的影响[J]. 林业科学研究, 2007, 20(3): 363-369.
[22]张元燕, 虞木奎, 方炎明. 麻栎不同种源的表型性状变异分析[J]. 植物资源与环境学报, 2014, 23(3): 36-44.
[23]侯爱菊, 徐德昌. 植物高光效基因工程育种[J]. 中国生物工程杂志, 2005, 25(9): 19-23.
[24]张斌斌, 姜卫兵, 韩键, 等. 桃光合性能杂种优势的秋季日变化特征[J]. 中国生态农业学报, 2011, 19(1): 82-86.
[25]BIRADAR S, SALIMATH P M, PATIL B C. Land race as a source for improving photosynthetic rate and productivity in cowpea (VignaunguiculataW.)[J]. Electronic Journal of Plant Breeding, 2010, 1: 872-877.
[26]BELANE A K, DAKORA F D. Assessing the relationship between photosynthetic C accumulation and symbiotic N nutrition in leaves of field-grown nodulated cowpea (VignaunguiculataL. Walp.) genotypes[J]. Photosynthetica, 2015, 53: 562-571.
[27]褚延广, 苏晓华, 黄秦军, 等. 欧洲黑杨基因资源光合生理特征与生长的关系[J]. 林业科学, 2010, 46(7): 77-83.
[28]王沙生, 高荣孚, 吴贯明. 植物生理学[M]. 2版. 北京: 中国林业出版社, 1991: 136.
[29]孙佳音, 杨逢建, 庞海河, 等. 遮荫对南方红豆杉光合特性及生活史型影响[J]. 植物研究, 2007, 27(4): 439-444.
[30]PERRIN P M, MITCHELL F J G. Effects of shade on growth, biomass allocation and leaf morphology in European yew (TaxusbaccataL.)[J]. European Journal of Forest Research, 2013, 132: 211-218.
[32]WEI R P, HAN S D, DHIR N K, et a1. Population variation in growth and 15-year-old shoot elongation along geographic and climatic gradients in black spruce in Alberta[J]. Canadian Journal of Forest Research, 2004, 34: 1691-1702.
[33]PETTERLEA,KARLBERGA,BHALERAOR P. Daylength mediated control of seasonal growth patterns in perennial trees[J]. Current Opinion in Plant Biology, 2013, 16: 301-306.
(责任编辑: 佟金凤)
Analyses on differences in seedling growth, photosynthetic physiology and height growth rhythm of each family ofTaxuswallichianavar.maireifrom different locations
XIAO Yao1,2, CHU Xiuli1,①, YIN Zengfang2, JIANG Junma3, WANG Hui3, ZHOU Zhichun1
(1. Research Institute of Subtropical Forestry, Chinese Academy of Forestry, Fuyang 311400, China; 2. College of Biology and Environment, Nanjing Forestry University, Nanjing 210037, China; 3. Fuxi Forest Farm of Zhejiang Province, Chun’an 311700, China),J.PlantResour. &Environ., 2016, 25(1): 34-42
In order to fully understand differences in seedling growth, photosynthetic physiology and height growth rhythm of each family ofTaxuswallichianavar.mairei(Lemée et H. Lév.) L. K. Fu et Nan Li from different locations, taking two-year-old container seedlings of 20 families from Fenyi of Jiangxi (FY), Longquan of Zhejiang (LQ), Jian’ou of Fujian (JO) and Mingxi of Fujian (MX) as research objects, height and ground diameter of seedlings, and net photosynthetic rate and chlorophyll content in leaf were compared, and Logistic fitting regression analysis on height growth of each family was carried out and fitting curve parameters and height growth rhythm parameters were calculated and analyzed. On this basis, correlation among different indexes was analyzed and cluster analysis of 20 families tested was carried out. The results show that there is no significant difference in height, ground diameter, chlorophyll content and net photosynthetic rate ofT.wallichianavar.maireiamong different locations, but there are extremely significant differencesinthoseindexesamongdifferentfamilies (P<0.01). The maximum values of height, ground diameter, net photosynthetic rate (Pn) and contents of chlorophylla(Chla), chlorophyllb(Chlb) and total chlorophyll (Chl) of each family are higher than their minimum values by 52.99%, 29.63%, 115.65%, 39.81%, 30.95% and 37.24%, respectively. Height growth of four locations all appear obvious “S-shape” curve. The maximum values of the maximum growth rate (MGR), linear growth rate (LGR), total linear growth (TLG) and linear growth duration (LGD) of each family are higher than their minimum values by 60.00%, 61.54%, 63.73% and 22.73%, respectively. From the beginning (t1) and final (t2) days of linear growth, fast growing stage of each family is from middle or later May to early August. There are extremely significantly positive correlations of seedling height with indexes ofMGR,TLG,LGR, ground diameter, Pn, Chlband Chl, et al, and significantly positive correlation with Chla, while there are significantly positive correlations of ground diameter with indexes ofMGR,TLG,LGR, Pn and Chlb, et al, but there is no significant correlation of both height and ground diameter withLGD. The cluster analysis result shows that atEuclideandistanceof 10, 20 families tested are divided into four types, in which, seedling growth of 6 families of the Ⅳ type is the best and their growth potential is the highest. It is suggested that genetic variance of each family ofT.wallichianavar.maireifrom different locations is rich, and its height and ground diameter growth are mainly affected by growth rate. Overall, growth potential of seedlings of family LQ09, LQ11, FY21, FY26, MX12 and MX18 is higher, so they can be used as superior family ofT.wallichianavar.maireifor spreading.
Taxuswallichianavar.mairei(Lemée et H. Lév.) L. K. Fu et Nan Li; height growth rhythm; photosynthetic physiology; Logistic fitting regression analysis; correlation analysis; cluster analysis
10.3969/j.issn.1674-7895.2016.01.05
2015-08-17
浙江省农业新品种选育重大科技专项竹木育种协作组重点项目(2012C12908-5)
肖遥(1990—),男,四川成都人,硕士研究生,主要从事珍贵树种遗传育种方面的研究工作。
Q945; S791.49.01
A
1674-7895(2016)01-0034-09
