抗水稻黑条矮缩病转基因材料的鉴定分析
2016-08-10尹鲜思王少岭崔益平陈秋红王国梁王志龙
尹鲜思,王少岭,崔益平,陈秋红,王国梁,2,王志龙*
(1.湖南农业大学农学院,湖南 长沙 410128;2.美国俄亥俄州立大学植物病理系,俄亥俄 哥伦布 43210)
抗水稻黑条矮缩病转基因材料的鉴定分析
尹鲜思1,王少岭1,崔益平1,陈秋红1,王国梁1,2,王志龙1*
(1.湖南农业大学农学院,湖南 长沙 410128;2.美国俄亥俄州立大学植物病理系,俄亥俄 哥伦布 43210)
针对水稻黑条矮缩病毒(RBSDV)的核心粒子外壳蛋白基因(S8)、毒质与非结构蛋白基因(S9)及其嵌合基因(S8&S9)构建了6个RNAi载体,经农杆菌介导转化武陵粳1号获得转基因植株。通过潮霉素溶液浸泡法对转基因植株进行初步筛选,剔除假阳性植株;再通过PCR从DNA水平进行转基因阳性鉴定;进一步通过qRT–PCR 从RNA水平进行转基因表达量分析。选取qRT–PCR表达量高的单拷贝纯系S8后代进行灰飞虱传毒试验,结果表明,含有S8 RNAi片段的转基因水稻材料对RBSDV具有一定抗性。
水稻;水稻黑条矮缩病;转基因株系;RNA干扰;荧光定量PCR
投稿网址:http://xb.ijournal.cn
水稻黑条矮缩病是由水稻黑条矮缩病毒(RBSDV)引起的一种病毒性病害,发病植株表现为矮化、分蘖增多,叶片短阔、僵直,叶色浓绿,叶脉和茎秆呈蜡白色,渐变为褐色短条瘤状隆起,穗小或不抽穗结实[1–3]。RBSDV的宿主主要有水稻、玉米、小麦等禾本科植株,其传播媒介为灰飞虱和白背飞虱[4–6]。水稻黑条矮缩病20世纪50年代在日本被首次报道[7],并于1968年大规模暴发,导致粮食减产绝收[8]。20世纪 60年代在中国首次被报道[9–10]。20世纪90年代末以来,在中国南方稻区再次暴发流行,造成水稻大面积减产[11]。
水稻黑条矮缩病毒属呼肠孤病毒科斐济病毒属,其病毒粒子为直径75~80 nm的二十面体,包含双层衣壳[12–13],具有外层衣壳上的 A–突起和核心粒子表面的 B–突起[14]。RBSDV全基因组由 10条双链RNA(dsRNA)组成[15],分别命名为S1~S10,其对应编码蛋白分别为P1~P10,其中,S8具有1 927 bp,P8的相对分子质量为65 000,属小核衣壳结构蛋白,在RBSDV侵染过程中抑制转录[16]。S9具有1 900 bp,含2个非重叠ORFs,分别编码P9–1和P9–2,P9–1为聚集在侵染细胞的病毒基质,参与病毒复制和聚集,P9–2为非结构蛋白[17]。
Jia等[18]针对P7–1基因进行RNA干扰,有效抑制了病毒在昆虫媒介的传播;Shimizu等[10]对P9–1进行干扰,发现可以达到抗病效果。笔者针对水稻黑条矮缩病毒核心粒子外壳蛋白基因(S8)、毒质与非结构蛋白基因(S9)及 S8&S9嵌合片段,利用两步法快速高效构建了6个RNAi载体,经农杆菌介导转化武陵粳 1号,获得阳性转基因植株,并通过qRT–PCR选择双链RNA表达量高的单拷贝纯系进行了灰飞虱传毒试验与抗性评估,现将结果报道如下。
1 材料与方法
1.1材料
水稻武陵粳1号(高感RBSDV);已感染RBSDV水稻病株及叶片(贮藏于–80 ℃冰箱,扬州大学左示敏教授惠赠);灰飞虱(湖南农业大学饲养)。
1.2方法
1.2.1RNA干扰载体的构建
使用pENTR载体和pANDA载体,参照两步法[19],构建RBSDV S8、S9及S8&S9的RNA干扰载体(PCR引物见表1)。

表1 PCR和qRT–PCR引物信息Table 1 Sequence information for PCR and qRT–PCR primers
S8的RNAi干扰载体的构建:以含有RBSDV的水稻叶片cDNA为模板,扩增 S8干扰片段,将其连接到入门载体pENTR上,经BamHI、XhoI双酶切和测序验证后,再与目的载体pANDA进行重组反应,阳性重组子经菌落PCR及KpnI、SacI双酶切验证后,用于转化农杆菌。S9的RNAi干扰载体构建与S8的相同。
S8–1&S9–2嵌合片段载体的构建:以已构建的pENTR–S8–1为模板,S8–1–F与A引物扩增嵌合片段中的S8–1部分;以pENTR–S9–2为模板,B引物与S9–2–R引物扩增嵌合片段中的S9–2部分,A引物与B引物为反向互补序列(表1),将2个片段分别切胶回收,共同作为模板,S8–1–F与S9–2–R引物扩增整个嵌合片段,再连接到入门载体pENTR上,BamH I、XhoI双酶切验证,测序正确后,与pANDA进行重组反应,重组子采用菌落PCR和质粒KpnI、SacI双酶切验证,正确重组子用于转化农杆菌。S8–2&S9–1的 RNAi干扰载体构建与S8–1&S9–2的相同。
1.2.2根癌农杆菌介导的水稻遗传转化
运用电击法,将构建的 RNAi载体 pANDA–S8/S9/S8&S9转入根癌农杆菌EHA105中,挑取阳性克隆进行PCR验证和双酶切(KpnI和SacI)验证。采用农杆菌介导的水稻转化法转化水稻黑条矮缩病高感品种武陵粳1号,采用潮霉素筛选法,获得转基因植株。
1.2.3转基因阳性植株的鉴定与表达分析
将得到的转基因 T1代植株从生根培养基上取出,洗净根部培养基,移植到大田,待植株健壮后,剪取少许叶片置于含有潮霉素溶液(质量分数0.1%)的管子中浸泡5~7 d;选取潮霉素溶液鉴定阳性(叶片仍为绿色)的植株,提取其叶片基因组 DNA,使用目的引物(表1)进行转基因PCR阳性鉴定。再对PCR鉴定为阳性的植株进行qRT–PCR表达量分析,确定插入片段的转录水平,以鉴定转基因阳性表达效率。进一步选取转基因T2代及T3代阳性植株,进行潮霉素溶液鉴定,PCR及qRT–PCR分析,筛选高表达目的片段的T3代单拷贝转基因纯系。
1.2.4灰飞虱室内传毒与转基因植株的抗性鉴定
将挑选的T3代单拷贝转基因纯系浸种发芽,水稻幼苗在温室采用长日照培养(光照 14 h,温度26 ℃,相对湿度80%;黑暗10 h,温度20 ℃,相对湿度保持不变)。在人工气候模拟培养箱中饲养灰飞虱,恒温27 ℃,相对湿度70%,光照16 h。用感染RBSDV的水稻叶片饲养灰飞虱。待T3植株长到四叶期进行灰飞虱传毒,持续20 d,期间不定时驱赶灰飞虱取食待传毒水稻叶片,保证传毒效率最大化。传毒完成后,随机抓取一定量传毒灰飞虱,提取RNA,RT–PCR检测其带毒率。将完成传毒的水稻幼苗移栽到大田中,观察水稻抽穗期表型,确定水稻是否发病。
2 结果与分析
2.1RNAi载体的构建与验证
选取编码RBSDV结构蛋白基因S8以及S9作为构建RNAi干扰载体的候选基因,利用入门载体及LR重组反应[19],构建2个S8片段S8–1和S8–2以及2个S9片段S9–1和S9–2的干扰载体(图1)。
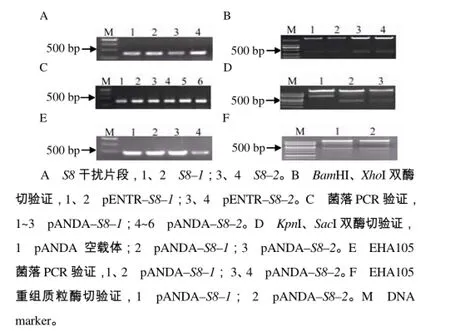
图1 RBSDV S8 RNAi载体的构建Fig. 1 The construction of RNAi vector for RBSDV S8
为了增强干扰效果,将S8基因和S9基因利用Overlapping PCR方法拼接到一起构成一个较大的RNA干扰片段,共构建2个嵌合片段 S8–1&S9–2 和S8–2&S9–1的RNAi载体,pANDA–S8&S9的构建如图2所示。
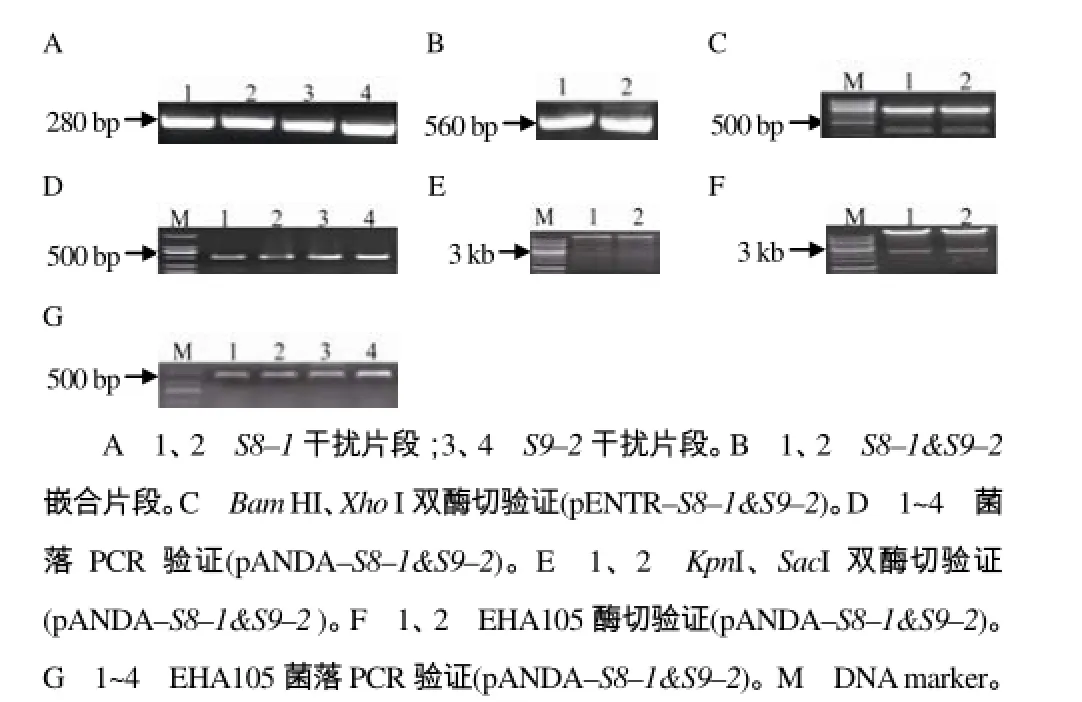
图2 RBSDV S8&S9 RNAi载体构建Fig.2 The construction of RNAi vector for RBSDV S8&S9
2.2转基因水稻植株的阳性鉴定
对经农杆菌介导法转化获得的转基因水稻植株,采取了潮霉素溶液浸泡法、PCR扩增以及qRT–PCR表达量分析3个步骤进行鉴定。
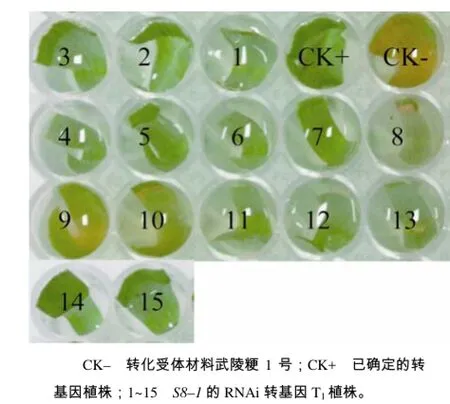
图3 RBSDV S8–1转基因植株的潮霉素浸泡鉴定结果Fig.3 The identification of transgenic plants with RBSDV S8–1 by hygromycin detection
将得到的转基因 T1代植株采用潮霉素浸泡法进行初步筛选。S8–1的RNAi转基因株系,阳性植株具有潮霉素抗性,叶片表现为绿色(如 CK+,已确定的转基因植株),未转基因的 CK–叶片呈黄色(图3),由此初步鉴定1~8号、11~15号为转基因阳性,9号和10号为转基因阴性植株。将初步鉴定的转基因阳性植株(1~8号、11~15号)提取DNA,以未转基因植株为阴性对照,载体pANDA–8–1为阳性对照,使用目的片段引物进行 PCR扩增,如图4–A所示。1~8号及11~15号的扩增片段与阳性对照大小一致,确定这13株植株中插入了目的基因片段。从中选取 7株(1、2、3、5、6、14、15号)进行qRT–PCR表达量分析,如图4–B,以武陵粳1号为对照组(WT),RBSDV基因与水稻基因组无同源性,测量S8与水稻泛素基因(Ubiquitin,UBQ)的表达量比值(S8/UBQ),与对照组相比,7株转基因材料相对表达量均有一定程度的增加,其中在8号植株中增加约60倍,在2号和14号植株中分别增加约36、41倍,进一步表明RBSDV外源片段已导入水稻中,并且可以在水稻中转录,为转基因阳性植株。对转基因阳性植株的T2代,继续采用上述3个步骤筛选,以获得目的基因单拷贝插入的高表达株系,并筛选其T3代纯系用于病毒传毒鉴定。
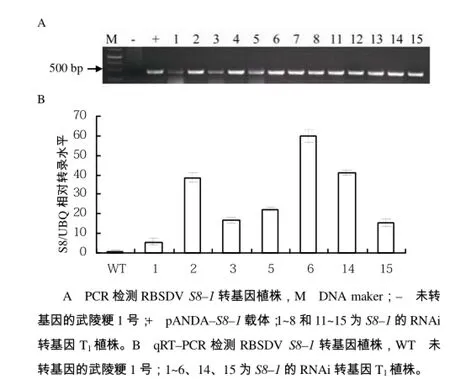
图4 RBSDV S8–1转基因植株鉴定结果Fig.4 The identification of transgenic plants with RBSDV S8–1
按照以上筛选方式,对pANDA–S9–1转基因阳性植株进行鉴定,如图 5–A所示。PCR鉴定显示14株T1转基因植株中,有12株(1~6号、9~14号)为阳性植株,2株(7、8号)为阴性植株。选取2号和5号植株的T2代高表达株系进行qRT–PCR表达量分析,结果(图5–B)显示,T2转基因植株2–4的相对表达量高达约800倍,T2转基因植株2–1和5–1的相对表达量约600倍,其余植株的相对表达量约200~400倍,这些高表达株系将用于筛选单拷贝的T3代纯系。
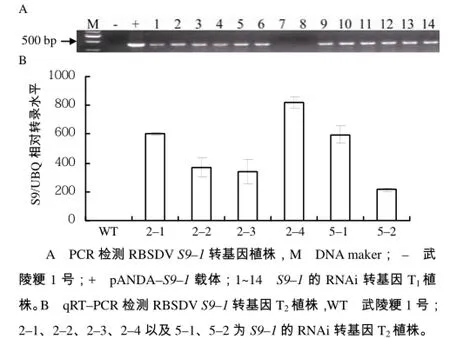
图5 RBSDV S9–1转基因植株鉴定结果Fig.5 The identification of transgenic plants with RBSDV S9–1
同样地,对pANDA–S8&S9转基因阳性植株进行鉴定,如图6–A所示。结果显示,17株T1转基因植株中,有16株(1~11号、 13~17号)为阳性植株,1株(12号)为阴性植株。选取1、25和30号植株的T2代进行qRT–PCR表达量分析,结果显示(图6–B),植株1号的3株T2代(1–2号、1–4号, 1–6 号)相对表达量约为20~60倍,其余植株的相对表达量均低于20倍,相对表达量高于20倍的株系被考虑用于筛选T3代的单拷贝纯系。
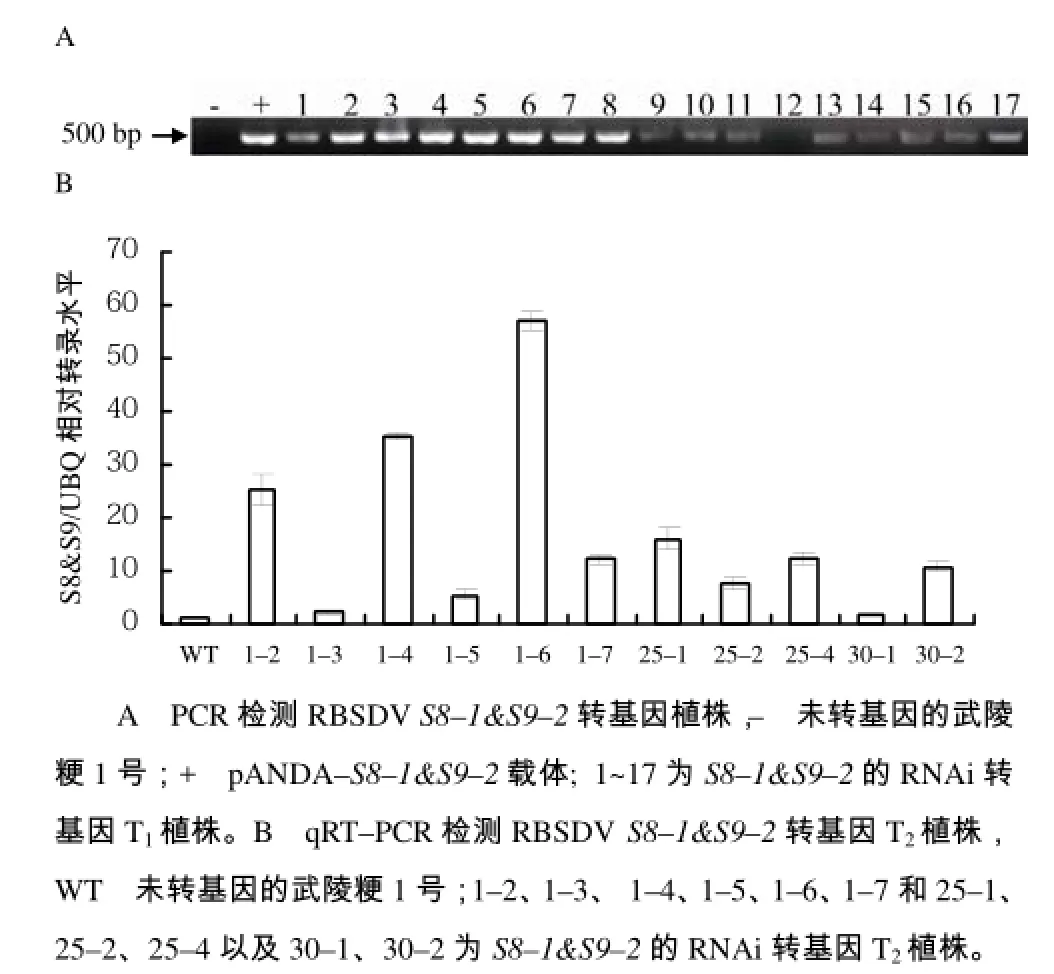
图6 RBSDV S8–1&S9–2转基因植株鉴定结果Fig.6 The identification of transgenic plants with RBSDV S8–1&S9–2
2.3转基因水稻植株的传毒接种与鉴定
在进行传毒试验之前,提取RBSDV病毒供体水稻单株材料叶片 RNA,采用 S8基因引物进行RT–PCR分析,以水稻Ubiquitin基因作为对照,检测供体材料带毒情况,如图 7–A所示。高表达RBSDV病毒S8基因的水稻单株4号和5号可以用来作为接种RBSDV的供体。

图7 感染RBSDV水稻病株鉴定Fig.7 The identification of infected plants with RBSDV and inoculation of RBSDV by Lalielphax striatellus
根据图4–B的qRT–PCR表达量分析结果,选取S8–1的6号转基因株系T3后代纯系种子,于播种后植株2~3叶期,开始采用离体叶片法,进行灰飞虱传毒,如图7–B所示。传毒完成后,通过检测传毒灰飞虱的带毒率来确定传毒效果,若带毒率高于80%便认定传毒有效。
经有效传毒后,植株在同等生长条件下,继续生长25 d,观察表型变化。已接受传毒的武陵粳1号与转基因材料相比,植株生长缓慢,部分植株干枯而死,叶片上出现条纹病斑,而携带有 RBSDV 的S8–1 RNAi干扰载体的转基因植株以及未进行传毒的武陵粳1号则正常生长,植物无枯死现象,叶片也无明显病斑(图 8–A、图 8–B)。温室接种数据初步表明,含有 S8–1 RNAi片段的转基因材料对RBSDV具有一定抗性。

图8 S8–1转基因植株传毒后的表型Fig.8 The phenotypes of S8–1 transgenic plants after RBSDV inoculation
3 结论与讨论
水稻黑条矮缩病具暴发性、间歇性和迁移性,其防控难度极大。针对RBSDV的S8、S9及S8&S9嵌合基因,通过两步法构建了6个RNAi载体,转化高感水稻品种武陵粳1号,获得了S8、S9及S8&S9 的RNAi转基因材料。由于获得的转基因材料较多,因而先采用潮霉素浸泡法(较简便有效)对转基因材料进行初步筛选,排除假阳性,以降低后续工作量。接着利用PCR鉴定目的基因是否整合到阳性植株基因组中,再通过qRT–PCR测量目的基因在转基因植株的相对表达量。通过上述对水稻转基因阳性植株的层层递进鉴定,成功筛选到了 S8、S9及 S8&S9嵌合基因的RNAi转基因阳性植株,为RBSDV室内接种及田间抗性鉴定提供了材料基础。
在进行RBSDV室内接种前,先确定田间采集的水稻矮化病株是否带毒。在接种过程中,检测了新鲜的及冻存的水稻带毒叶片,发现二者均能使灰飞虱有效传毒,特别是冻存的水稻带毒叶片,能够全年安排灰飞虱传毒试验,进行田间抗性鉴定。
在目前获得的转基因材料中,仅对S8的RNAi株系进行了室内接种鉴定及表型观察,初步结果表明,含有S8 RNAi片段的转基因水稻对RBSDV具有一定抗性,但还有待大量接种病毒后的田间群体表型鉴定。同样,含有S9以及S8&S9 RNAi片段的转基因材料也需要室内大量接种和田间群体表型鉴定。由于使用的目的载体pANDA含有筛选标记抗潮霉素基因,考虑到水稻为重要粮食作物,将在后续试验中采用无筛选标记转基因技术获取抗RBSDV的转基因材料,以便用于水稻育种与种质创新。
[1] Conti M,Lovisolo O.Tubular structures associated with maize rough dwarf virus particles in crude extracts:electron microscopy study[J].Journal of General Virology,1971,13(1):173–176.
[2] Milne R G,Lovisolo O.Maize rough dwarf and related viruses[J].Advances in Virus Research,1977,21:267–341.
[3] 王华弟,祝增荣,陈剑平,等.水稻黑条矮缩病发生流行规律监测预警与防控关键技术[J].浙江农业学报,2007,19(3):141–146.
[4] Identification of rice black streaked dwarf Fijivirus in maize with rough dwarf disease in China[J].Archives of Virology,2001,146(1):167–170.
[5] Bai F W,Yan J,Qu Z C,et al.Phylogenetic analysis
reveals that a dwarfing disease on different cereal crops in China is due to rice black streaked dwarf virus(RBSDV)[J].Virus Genes,2002,25(2):201–206.
[6] 中国科学院上海生物化学研究所病毒组.水稻黑条矮缩病病原体的研究Ⅰ.传毒灰稻虱体中类似病毒的质粒[J].中国科学:A辑,1974,17(2):158–167.
[7] Kuribayashi K,Shinkai A.On the rice black-streaked dwarf disease,a new disease of rice in Japan[J].Annals of the Phytopathological Society of Japan,1952,16:41.
[8] Ishii M,Yoshimura S.Epidemiological studies on rice black-streaked dwarf virus in Kanto–Tosan district in Japan[J].J Cent Agric Exp Sta,1973,17:61–121.
[9] Wu J,Ni Y,Liu H,et al.Development and use of three monoclonal antibodies for the detection of rice black-streaked dwarf virus in field plants and planthopper vectors[J].Virology Journal,2013,10:114.
[10] Shimizu T,Nakazono–Nagaoka E,Akita F,et al. Immunity to rice black streaked dwarf virus,a plant reovirus,can be achieved in rice plants by RNA silencing against the gene for the viroplasm component protein[J]. Virus Research,2011,160(18):400–403.
[11] Wang H D,Chen J P,Wang A G,et al.Studies on the epidemiology and yield losses from rice black-streaked dwarf disease in a recent epidemic in Zhejiang province,China[J].Plant Pathology,2009,58(5):815–825.
[12] Redolfi P,Boccaroo G.Fractionation of the doublestranded RNA of maize rough dwarf virus subviral particles[J].Virology,1974,59(1):319–322.
[13] Shikata E,Kitagawa Y.Rice black-streaked dwarf virus:its properties,morphology,and intracellular localization [J].Virology,1977,77(2):826–842.
[14] 邓金奇,戴良英.水稻黑条矮缩病毒的研究进展[J].中国农学通报,2012,28(18):209–212.
[15] 孙枫,徐秋芳,程兆榜,等.中国水稻黑条矮缩病研究进展[J].江苏农业学报,2013,29(1):195–201.
[16] Liu H,Wei C,Zhong Y,et al.Rice black-streaked dwarf virus minor core protein P8 is a nuclear dimeric protein and represses transcription in tobacco protoplasts[J]. FEBS Letters,2007,581(13):2534–2540.
[17] Isogai M,Uyeda I,Lee B C.Detection and assignment of proteins encoded by rice black streaked dwarf Fijivirus S7,S8,S9 and S10[J].Journal of General Virology,1998,79:1487–1494.
[18] Jia D,Mao Q,Chen H,et al.Virus-induced tubule:a vehicle for rapid spread of virions through basal lamina from midgut epithelium in the insect vector[J].Journal of Virology,2014,88(18):10488–10500.
[19] 王少岭,杜强,尹鲜思,等.两步法构建灰飞虱RNAi载体及水稻遗传转化[J].作物研究,2015,29(3):230–234.
责任编辑:罗慧敏
英文编辑:罗 维
Identification and analysis of transgenic lines against rice black-streaked dwarf virus
Yin Xiansi1, Wang Shaoling1, Cui Yiping1, Chen Qiuhong1, Wang Guoliang1,2, Wang Zhilong1*
(1.College of Agronomy, Hunan Agricultural University, Changsha 410128, China; 2.Department of Plant Pathology, The Ohio State University, Columbus, Ohio 43210, USA)
Genome segments S8 (encoding minor core capsid protein), S9 (encoding virplasm and non-structual protein) and their chimeric sequences S8&S9 of RBSDV were used to construct six RNAi vectors and transferred into rice cultivar Wulingjing 1 to generate transgenic plants by Agrobacterium-mediated transformation. Transgenic plants were first screened by immersion with hygromycin solution to eliminate the false-positive plants, then identified by PCR genotyping analysis from the DNA level,followed by qRT–PCR analysis of candidate genes at the RNA level. Single-copy transgenic lines with high expression of S8 detected by qRT–PCR were picked up for Laodelphax striatellus-mediated virus transmission test, and the results showed that transgenic lines containing S8 RNAi vector displayed resistance to RBSDV.
rice; rice black-streaked dwarf virus (RBSDV); transgenic lines; RNA interference (RNAi); qRT–PCR
;*通信作者,王志龙,博士,教授,主要从事作物遗传育种与水稻抗逆分子生物学研究,zhilongwang@126.com
Q786
A
1007-1032(2016)04-0380-06
2016–03–24 修回日期:2016–04–08
转基因生物新品种培育重大专项 (2012ZX08009001);教育部创新团队发展计划项目(IRT1239);湖南农业大学大学生创新性实验计划项目(XCX15145)
尹鲜思(1990—),男,湖南邵阳人,硕士研究生,主要从事水稻抗逆分子生物学研究,1614974545@qq.com;王少岭为共同
