干旱胁迫下广东石漠化地区造林树种光合和耗水特性
2016-08-09吴俊文李吉跃白晶晶
吴俊文, 刘 珊, 李吉跃, 何 茜, 苏 艳, 邱 权, 白晶晶, 王 燕, 贾 朋
华南农业大学林学与风景园林学院, 广州 510642
干旱胁迫下广东石漠化地区造林树种光合和耗水特性
吴俊文, 刘珊, 李吉跃*, 何茜, 苏艳, 邱权, 白晶晶, 王燕, 贾朋
华南农业大学林学与风景园林学院, 广州510642
摘要:石漠化地区土层稀薄、干旱贫瘠、植被破坏、生态恶化等问题较为突出,人工恢复森林植被是一项快速有效的生态恢复途径。采用盆栽苗木称重法和Li- 6400光合系统测定方法分别测定3个常见石漠化造林树种浙江润楠(Machilus leptophylla),枫香(Liquidambar formosana)和亮叶含笑(Michelia fulgens Dandy)苗木在不同土壤水分条件下耗水特性和光合特性,为进一步筛选和评价石漠化地区造林树种抗旱特性提供理论依据。研究结果表明:(1)整个干旱胁迫期间,3种幼苗叶片的净光合速率(Pn)、气孔导度(Gs)、蒸腾速率(Tr)和胞间CO2浓度(Ci)持续下降。重度干旱时,浙江润楠光合和水分利用效率分别是枫香的2.5倍、89.6倍和亮叶含笑的1.9倍、26.3倍。(2)干旱胁迫中期和后期,枫香和亮叶含笑净光合速率(Pn)和气孔导度(Gs)下降而胞间CO2浓度(Ci)升高,说明Pn下降的主要原因已经由气孔因素转变为非气孔因素的限制。(3)3个石漠化树种在不同干旱胁迫下日耗水总量和日平均耗水速率均存在显著性差异(P<0.05),白天耗水量占总耗水量74%—92%之间。浙江润楠在重度干旱时期还对水分要求还比较大,维持一个相对较高的耗水速率,是枫香和亮叶含笑的3.7倍和2.2倍。(4)不同干旱胁迫下,3个石漠化树种耗水速率日变化均表现出单峰曲线,不同干旱胁迫下峰值点会发生变化。(5)综合来看,浙江润楠和枫香是相对高光合和高水分利用效率树种。隶属函数结果表明,在正常、轻度和重度干旱下抗旱能力均为枫香>浙江润楠>亮叶含笑,重度干旱下为浙江润楠>枫香>亮叶含笑。
关键词:石漠化造林树种;干旱胁迫;光合;耗水
石漠化是指在脆弱的喀斯特环境之下,人类不合理的活动造成了森林破坏,最终出现大面积裸露的岩石的土地退化现象[1]。广东省的石灰岩面积约有1.45万km2,岩溶地区主要分布在北部和西部,大多分布在边远山区,生态环境恶化,交通不便,植被稀少,水土流失严重,生活用水贫乏,农村贫困问题突出。石漠化是岩溶区域最主要的生态问题,也是制约经济和社会发展的瓶颈[2]。植被恢复是改善这种恶劣生态环境的重要手段,这个问题受到国内外学者广泛关注[3- 6]。针对粤北特殊的地理条件也有一系列报道关于石漠化造林树种筛选、造林技术和相关生理生态研究[7- 9]。然而造林面临的一个问题是石漠化地区土层较薄,保水能力较差,植被经常面临周期性干旱环境。所以石漠化地区造林首要解决的一个问题就是筛选适合这种生境的抗旱树种,只有了解了造林树种本身的耗水规律和抗旱能力,才能做到适地适树,提高造林成功率,对广东地区石漠化治理和生态恢复具有重要意义。
浙江润楠(Machilus leptophylla),枫香(Liquidambar formosana)和亮叶含笑(Michelia fulgensDandy)是粤北石灰岩地区造林树种之一,有学者研究了几个树种在粤北石灰岩山地造林早期的生长表现[10],也有学者已经研究过其他造林树种任豆Zenia insignis、车轮梅Rhaphiolepis indica、楝叶吴茱萸Evodia glabrifolia等的抗旱表现[9,11- 14],然而对于这3个树种抗旱方面研究还鲜有报道。本文选择浙江润楠、枫香和亮叶含笑幼苗为研究材料,采用盆栽苗木称重法和Li-6400光合系统测定方法,测定耗水特性和光合生理特性,研究不同水分条件下3个石漠化树种在水分消耗和光合特点,为进一步研究和筛选石漠化抗旱树种提供理论依据。
1材料与方法
1.1实验地点和材料
选择广东石漠化地区常见造林树种枫香、浙江润楠和亮叶含笑3年生实生苗,2013年7月上盆,所用花盆规格为 200mm× 150mm, 基质土壤取自华南农业大学树木园, 其田间持水量为(24.02±2.10) %, 容重为(1.15±0.09)g/cm3。试验期间定期除草浇水,防治病虫害,保证苗木正常生长。实验地点在华南农业大学林学与风景园林学院6楼温室,通过计算不同干旱时期温湿度日变化计算得到平均温度和湿度分别是(24.53±1.82)℃和(53.97±9.36)%。实验开始前测定苗木基本情况如表1所示。
1.2实验测定项目和方法1.2.1实验设计
2013年10月27开始到12月4日结束。每种选择 30 株生长正常、长势接近的苗木,对所有供试苗木浇透水后,停止浇水进行自然干旱处理,即用保鲜膜进行覆盖密 封盆栽土壤,同时将苗盆用塑料袋完全套住(从苗木根茎处覆盖整个表面, 并密封花盆底部),防止水分通过土壤表面蒸发,使得水分散失的唯一途径是通过植物蒸腾作用。土壤含水量采用FOM/mts便携式土壤湿度计进行连续监测, 即封盆后每种苗木固定 6盆,每间隔2—4d于 18: 00 测定盆栽苗木的土壤体积含水量,根据容重换算成土壤质量含水量,土壤质量含水量(%)=体积含水量(%)/土壤容重(g/cm3)(通常认为1g水的体积就是1cm3)。整个实验持续43d,直至苗木枯萎死亡。干旱梯度的划分参考其他学者土壤干旱胁迫划分方法[13,15- 17],设定土壤体积含水量占田间持水量80%以上为正常水平,50%—70%为轻度干旱,30%—50%为中度干旱,低于30%为重度干旱。

表1 苗木生长情况(平均值±标准差)

表2 各时期苗木土壤含水量占田间持水量比例 %(平均值±标准差)
1.2.2耗水量测定
本次研究中耗水量测定方法为盆栽苗木称重法,以防止土壤的水分的蒸发影响苗木蒸腾耗水的研究。每种苗木挑选生长中等的苗木6 盆测定单株耗水量,8:00—20:00每隔2h用SP- 30 电子天平 (美国) 称盆重1次,第2天8:00再称重1次(8:00—20:00测定结果为白天耗水量,20:00—次日8:00测定结果为夜晚耗水量)。注意在每个干旱期间挑选2—3个晴朗天气,避开天阴和下雨天气进行耗水量测定。每次称盆的同时采用便携式手持气象站记录每次称盆时的环境温、湿度,每次3个重复。
1.2.3单株叶面积
每株树用坐标纸比对将差不多大小的叶片分为一级,每级记录叶片数量,并找出各级典型叶片计算出叶面积,以分级叶片数×叶面积再相加之总和得到单株叶面积。
1.2.4耗水速率
耗水速率 = 每个时间段的耗水量 /(单株叶面积 × 时间)
1.2.5光合生理特性测定
在整个干旱胁迫进程中进行苗木叶片光合生理测定。于2013年10月27开始在每个干旱期间选择3个典型晴天于于 09: 00—11: 30 用Li- 6400 便携式光合作用分析系统(美国) 测定苗木叶片净光合速率 (Pn) 和蒸腾速率(Tr),每种苗木选择4 株, 每株测定3 片功能叶,其中测定过程中使用LI- 6400- 2B红蓝光源,光强设置为1200μmolm-2s-1,温度 35℃,相对湿度为(51.5±6.3)%,大气CO2浓度(399±9.2)μmol/mol。水分利用效率:WUE= 净光合速率Pn /蒸腾速率Tr。测定一直持续到到12月4日实验结束。
1.3数据分析
采用WPS2013和Excel2010 进行作图,SPSS19对实验数据进行ANOVA方差分析、 多重比较(Duncans法) 和相关性分析。采用隶属函数法对苗木抗旱性进行分析,这种方法采用Fuzzy数学中隶属函数的方法对树种各个抗旱指标值得隶属函数值进行累加,求平均数以评价抗旱性。抗旱性隶属函数值的计算方法如下:
如果某个指标与抗旱性呈现出正相关关系,可以用公式:X(μ)=(X-Xmin)/(Xmax-Xmin),如果某一指标与抗旱性呈现出负相关关系,用公式:X(μ)=1-(X-Xmin)/(Xmax-Xmin),公式中X(μ)为抗旱性隶属函数值,X为某一指标的测定值,Xmax为某一指标测定中的最大值,Xmin为某一指标测定中的最小值。
2结果与分析
2.1干旱胁迫对3个石漠化树种光合特性的影响2.1.1干旱胁迫对幼苗净光合速率的影响
如图1所示,不同树种在不同干旱胁迫时期以及同一干旱胁迫时期不同树种之间净光合速率(Pn)存在显著性差异(P<0.05)。轻度干旱LD时期,枫香、浙江润楠分别较对照CK降低了22.8%、37.3%,而亮叶含笑不变,分别是CK的77.2%、62.6%和104.1%。中度干旱MD时期,枫香、浙江润南和亮叶含笑分别较对照CK降低了42.3%、41.2%和51.3%。重度干旱SD时期,枫香、浙江润南和亮叶含笑分别较对照CK降低了91.7%、78.2%和99.6%。3个树种光合进行种间差异分析(图1)表明,正常水分条件下,亮叶含笑光合显著低于浙江润楠和枫香,轻度干旱下,枫香合显著高于浙江润楠和亮叶含笑。中度干旱下,亮叶含笑光合显著低于浙江润楠和枫香。重度干旱下,浙江润楠还维持较高光合,分别是枫香和亮叶含笑的2.5倍和89.6倍。
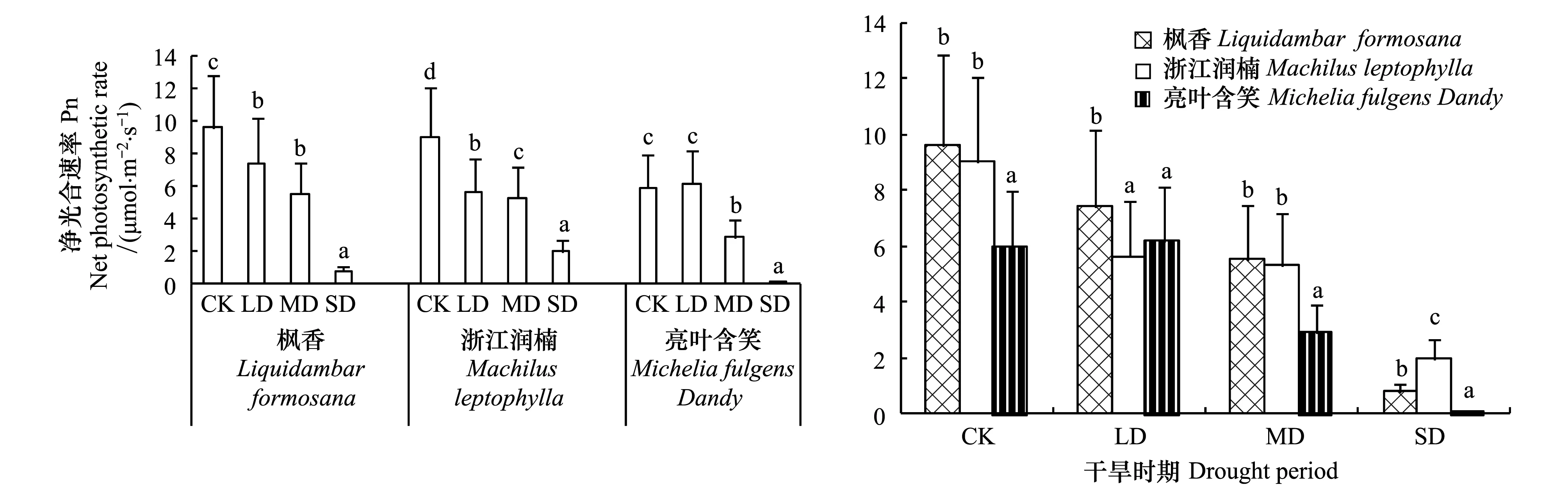
图1 3个石漠化树种不同干旱时期净光合速率(不同字母表示差异显著)Fig.1 Net photosynthetic rate of three rocky desertification tree species under different drought stress (different letters indicated significant difference)CK: 正常水分; LD: 轻度干旱; MD: 中度干旱; SD: 重度干旱
2.1.2干旱胁迫对幼苗胞间CO2浓度(Ci)、蒸腾速率(Tr)和气孔导度(Cond)的影响
如图2所示,枫香和亮叶含笑在干旱期间胞间CO2浓度(Ci)有显著差异(P<0.05)。不同干旱胁迫时期,3种幼苗气孔导度和蒸腾速率Tr均存在显著差异(P<0.05)。胞间CO2浓度(Ci)差异主要体现在中期MD和后期SD,枫香在中度干旱下增加了55.8%,亮叶含笑在中期和后期分别较CK增加了15.3%和119.6%。气孔导度(Cond)对水分胁迫比较敏感,轻度干旱时期,枫香和浙江润楠显著降低,而亮叶含笑则无显著变化。中度和重度干旱时期,枫香无显著差异,而浙江润楠和亮叶含笑则表现出显著差异。重度干旱下,枫香、浙江润楠和亮叶含笑分别降至CK的15.3%、25.2%和21.1%。蒸腾速率(Tr)方面,枫香在干旱前期没有表现出差异,直到重度干旱时期才表现出显著差异,较CK降低了74.9%。浙江润楠在轻度干旱胁迫时期就表现出差异,较CK降低了53.3%,之后则无明显差异。亮叶含笑在中期和后期表现出差异,分别降低了27.8%和81%。
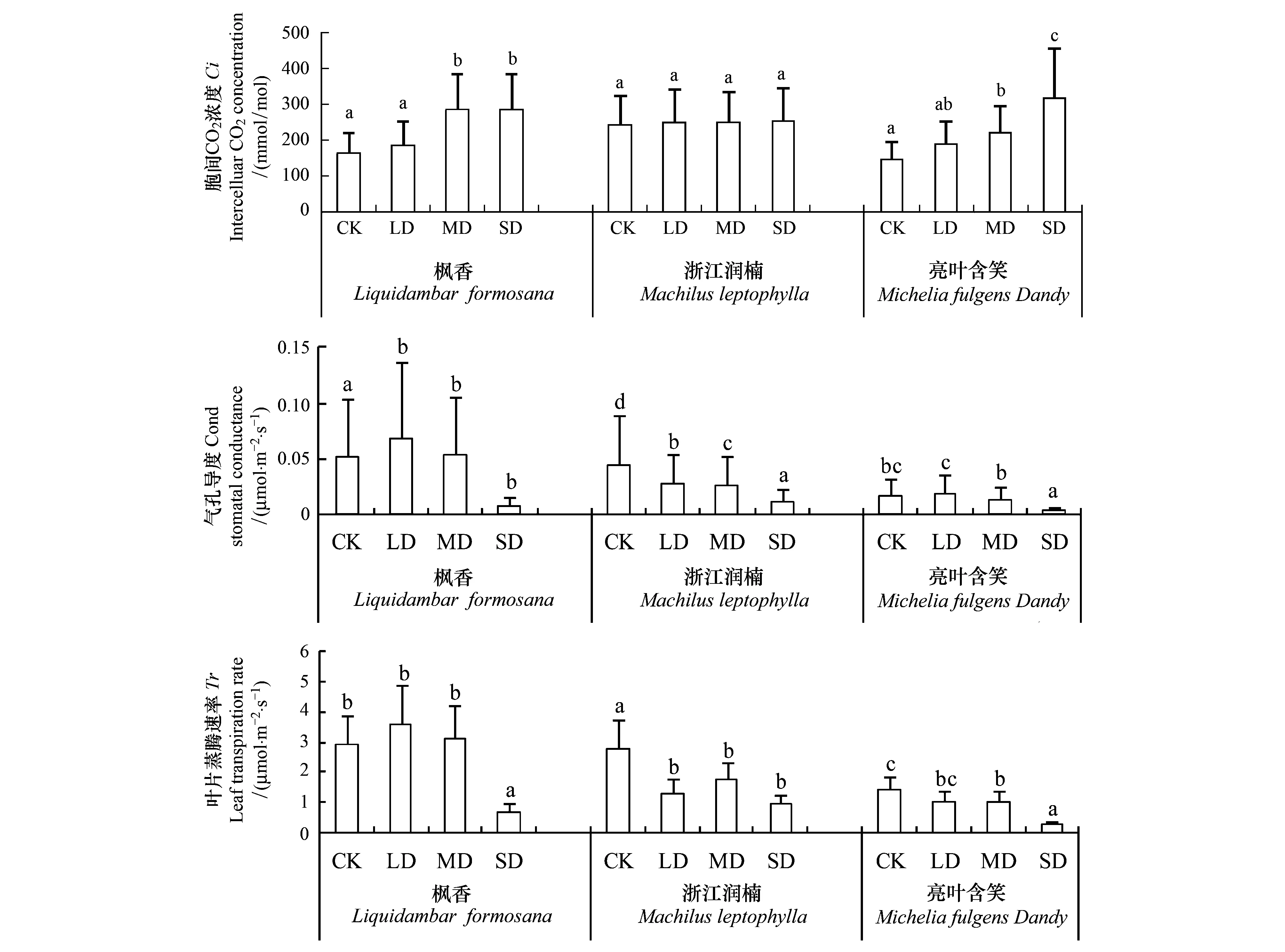
图2 3个石漠化树种不同干旱时期胞间CO2浓度Ci、叶片蒸腾速率Tr和气孔导度CondFig.2 Intercelluar carbon dioxide (Ci), leaf transpiration rate (Tr) and stomatal conductance (Cond) of three rocky desertification tree species under different drought stress
2.1.3干旱胁迫对幼苗水分利用效率(WUE)的影响
图3表明,不同干旱时期和同一干旱时期3个树种水分利用效率WUE均存在显著差异(P<0.05)。枫香和浙江润楠水分利用效率均随土壤含水量的下降而降低,而亮叶含笑则是先升高而后降低。轻度干旱时期枫香水分利用效率显著降低,降低了38.5%,浙江润楠无差异,亮叶含笑则显著增加了38.7%。中度干旱时期枫香降低了46.7%,浙江润楠水分利用效率无显著变化,而亮叶含笑则降低了32.7%。重度干旱时期,枫香和亮叶含笑都与对照有显著差异,分别降低了67.5%和98.2%,而浙江润楠则与对照无显著差异。种间差异方面,浙江润楠一直维持一个较高的水分利用效率,枫香次之,亮叶含笑则在轻度和中度时候水分利用效率较高,重度干旱集聚下降,仅为CK的1.8%,且重度干旱下,浙江润楠水分利用效率分别是枫香和亮叶含笑的1.9和26.3倍。
2.2干旱胁迫对3个石漠化树种蒸腾耗水特性的影响
2.2.1耗水量比较
蒸腾耗水特性是植物利用水分状况最直接体现,本实验中苗木盆用膜覆盖密封处理,苗木蒸腾耗水是树苗向外界失水的唯一途径。由图4可以看出,3个树种不同土壤水分条件下全天耗水量和耗水速率有显著差异(P<0.0.5)。3个树种在正常水分CK和轻度干旱LD存在显著差异,降幅在33%—35%之间,而在中度MD和重度SD干旱下则表现出无显著差异。白天耗水占全天耗水比例在74%—92%之间,夜间耗水占全天耗水比例在7%—17%之间。
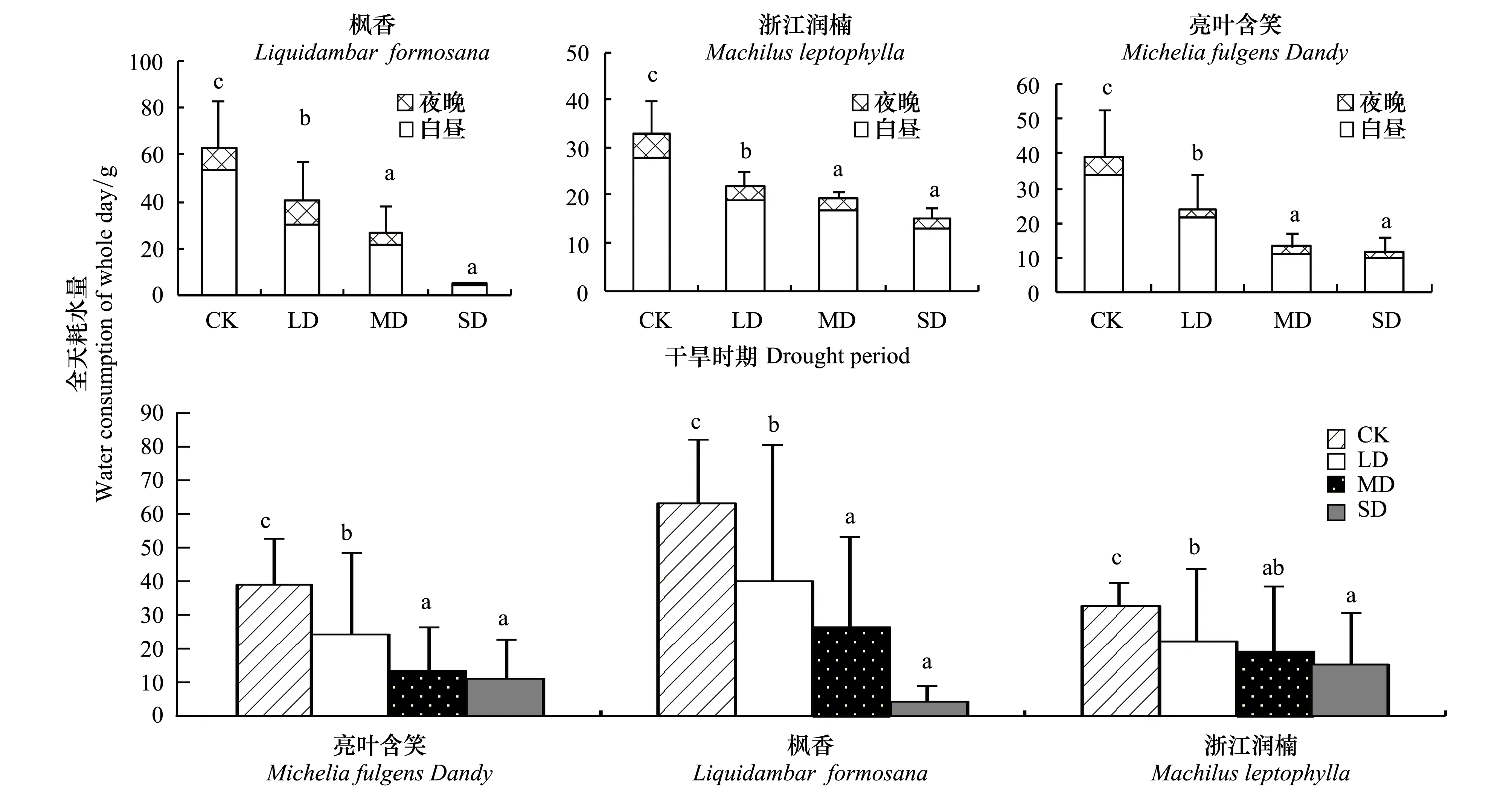
图4 3个石漠化树种不同干旱时期全天耗水量比较Fig.4 Water consumption of the whole day of three rocky desertification tree species
2.2.2耗水速率日变化
苗木蒸腾耗水量可以在一定程度上表现不同苗木蒸腾耗水差异,但是由于叶面积的不同,耗水量不能够完整反映出耗水能力和节水能力的强弱。要比较不同树种的耗水和节水性能需要考虑苗木的叶面积,即以蒸腾耗水速率来比较,蒸腾速率是由耗水量和叶面积来决定的。图5可以看出4个不同水分条件下3个树种的耗水速率均呈单峰曲线。
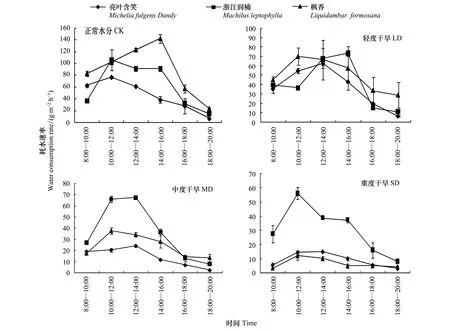
图5 不同土壤水分条件下苗木白天各时段耗水速率(±标准误差)Fig.5 Water consumption rate (±standard error) of seedlings in each period in the daytime under different soil moisture (mean ± SD)

图6 不同干旱胁迫下3个石漠化树种白天平均耗水速率Fig.6 Average water consumption rate of three rocky desertification tree species under different drought stress
正常水分条件下,浙江润楠和亮叶含笑峰值出现在10:00—12:00,枫香峰值出现在14:00—16:00,最大耗水速率排序为枫香(142.13±7.57)gm-2h-1>浙江润楠(106.1±17.40)gm-2h-1>亮叶含笑(76.9±1.40)gm-2h-1,日耗水速率种间差异较大。轻度水分胁迫下,亮叶含笑峰值时间段不变,浙江润楠变为14:00—16:00,枫香峰值提前到10:00—12:00,最大耗水速率排序为浙江润楠(73.2±3.11)gm-2h-1>枫香(69.9±9.03)gm-2h-1>亮叶含笑(62.6±2.47)gm-2h-1,种间相差不大,变化范围介于41.1—62.1gm-2h-1。中度干旱胁迫下,亮叶含笑峰值时间段不变,浙江润楠峰值时间段变为10:00—12:00,枫香没有变依然为10:00—12:00,最大耗水速率排序为浙江润楠(67.2±0.87)gm-2h-1>枫香(37.6±2.73)gm-2h-1>亮叶含笑(24.2±1.07)gm-2h-1,变化范围介于21.5—59.4gm-2h-1。重度干旱胁迫下,3个树种的峰值点均是10:00—12:00,最大耗水速率排序为浙江润楠(56.1±4.36)gm-2h-1>亮叶含笑(14.2±1.46)gm-2h-1>枫香(12.2±2.94)gm-2h-1,变化范围介于11.8—48.1gm-2h-1。枫香、浙江润楠和亮叶含笑在轻度干旱时期最大耗水速率分别降低了69.9%、73.2%和62.6%,中度干旱时期分别降低了73.6%、36.6%和68.5%,重度干旱时期分别降低了91.4%、47.1%和81.6%。
2.2.3白天平均耗水速率
由图6可知,同一树种在不同干旱时期白天平均耗水速率有显著差异(P<0.05)。枫香和亮叶含笑在正常水分CK和轻度干旱LD下白天平均耗水速率无显著差异,中度干旱后显著降低,浙江润楠则是轻度干旱就显著降低。轻度干旱LD时期,枫香、亮叶含笑和浙江润楠分别占正常水分的72.2%、67.2%和74.8%,降幅在25.5%—32.7%之间。中度干旱MD时期,枫香、亮叶含笑和浙江润楠分别占正常水分的52.8%、47.4%和70.6%,较CK下降幅度在29.4%—52.6%之间。重度干旱SD时期,枫香、亮叶含笑和浙江润楠分别占正常水分的10.2%、30.4%和50.8%,较CK下降幅度在49.2%—89.8%之间。
同一干旱时期(轻度和重度)不同树种白天平均耗水速率有显著差异(P<0.05),轻度干旱时期枫香白天平均耗水速率显著高于亮叶含笑和浙江润楠,重度干旱SD时期则是浙江润楠显著高于枫香和亮叶含笑。枫香在前期有较高的白天耗水速率,后面降得很低,浙江润楠则是在重度干旱时期还具有较大的耗水速率,分别是枫香和亮叶含笑的3.7倍和2.2倍。
2.3耗水速率与温度和湿度的相关系数
有研究表明,耗水速率和环境因子如温度湿度有密切联系[18- 21]。如图3所示,不同土壤含水量下不同树种蒸腾速率受环境温度和湿度游戏那个不一致。亮叶含笑在整个干旱胁迫期间与环境温度显著正相关,除重度干旱外与环境湿度呈显著负相关。浙江润楠和枫香除重度胁迫外均与环境温度呈显著正相关,除浙江润楠在正常水分条件下与环境湿度相关性不显著外,整个干旱期间两个树种均与环境湿度显著负相关。说明在整个干旱期间,3个树种的耗水速率均与环境温度和湿度显著相关,受环境因子影响较大,而浙江润楠和枫香在重度干旱时期与环境温度相关关系不显著则说明重度干旱时期环境因子可能不是主导因子。可能与重度干旱时期植物的内部生理代谢功能受损有关,对外界的环境因子变化不敏感。土壤水分充足时,环境因子湿度和温度是影响耗水速率的重要因子,当干旱加剧时,环境因子湿度和温度影响不显著。然而环境湿度与环境温度谁是主要影响因子则规律不明显,与环境因子对尾巨桉和竹柳的耗水速率研究结果不一致[22]。
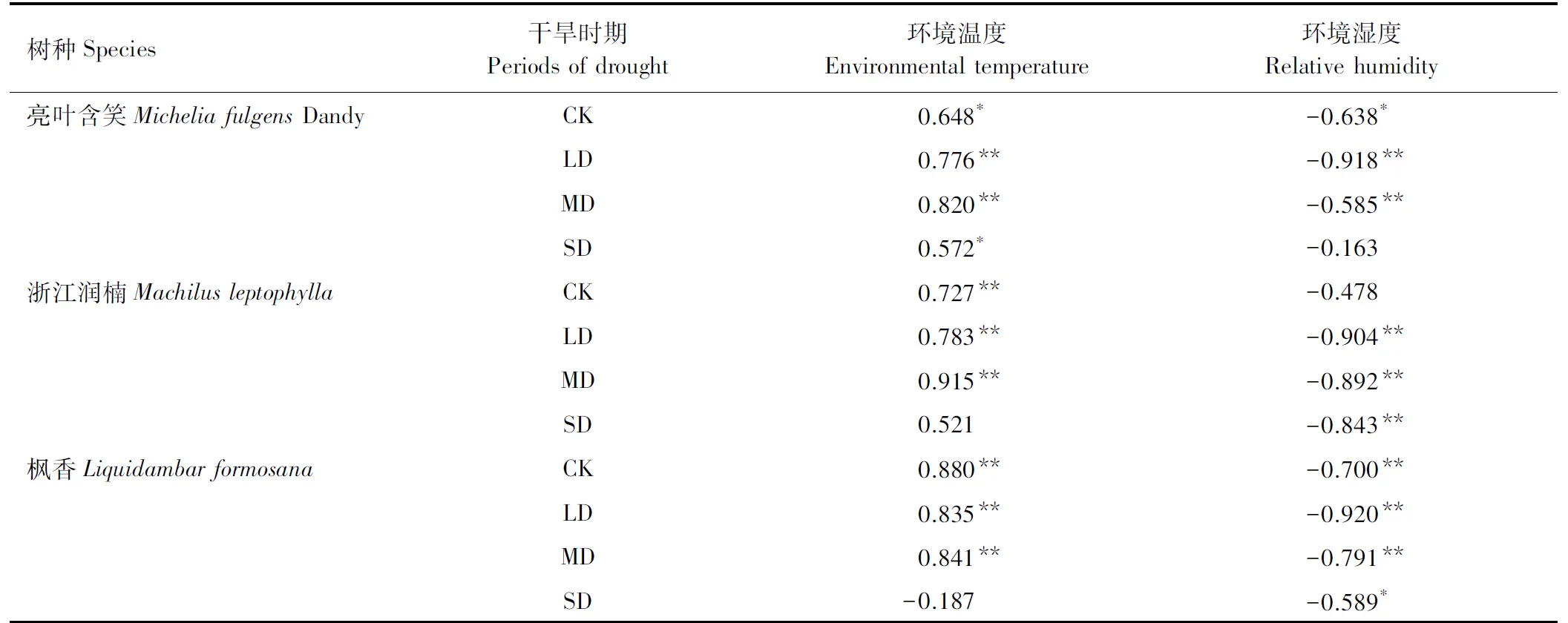
表3 耗水速率与环境温度和湿度相关系数
* 表示在 0.05 水平上显著相关; ** 表示在 0.01 水平上显著相关
2.43种苗木抗旱能力综合评价
方差分析结果表明(表4),净光合速率(Pn)、胞间CO2浓度(Ci)、气孔导度(Cond)、蒸腾速率(Tr)、日耗水量、耗水速率均呈现出种间差异(P<0.05)。而且不同干旱时期不同树种之间也存在差异(P<0.05)。故可以选择这几种指标分不同干旱梯度进行综合分析。如表4所示,在正常水分CK、轻度干旱LD、中度干旱MD时期,隶属函数值排名均是枫香>浙江润楠>亮叶含笑,而在重度干旱SD时期抗旱排名则是浙江润楠>枫香>亮叶含笑。
3结论与讨论
3.1不同土壤水分对石漠化树种耗水特性的影响
耗水量和耗水速率在抗旱树种筛选时经常作为重要评价指标,可以很好表征树种耗水能力差异[23- 26]。耗水速率是指植物单位时间单位面积的耗水量,作为植物内部的水分生理特征,具有遗传稳定性,可以用来评价比较不同植物固有的耗水能力[23]。本研究表明,在干旱胁迫过程中,耗水量和耗水速率均呈现出大幅下降趋势。且3个树种在整个干旱胁迫过程中耗水速率日变化均表现为单峰曲线,与其他树种桉树无性系、秋枫、竹柳、黄连木表现出一致的特征[22,27- 28],但是与欧洲云杉、奥地利黑松双峰曲线不同[29- 30],这可能是针叶树种与阔叶树种叶片内部结构有差异所致。
表43个石漠化树种不同干旱时期抗旱生理指标隶属函数值及综合评价
Table4Subordinatefunctionvaluesofthreerockydesertificationtreespeciesandtheircomprehensiveevaluationresultsofdroughtresistanceunderdifferentdroughtstress
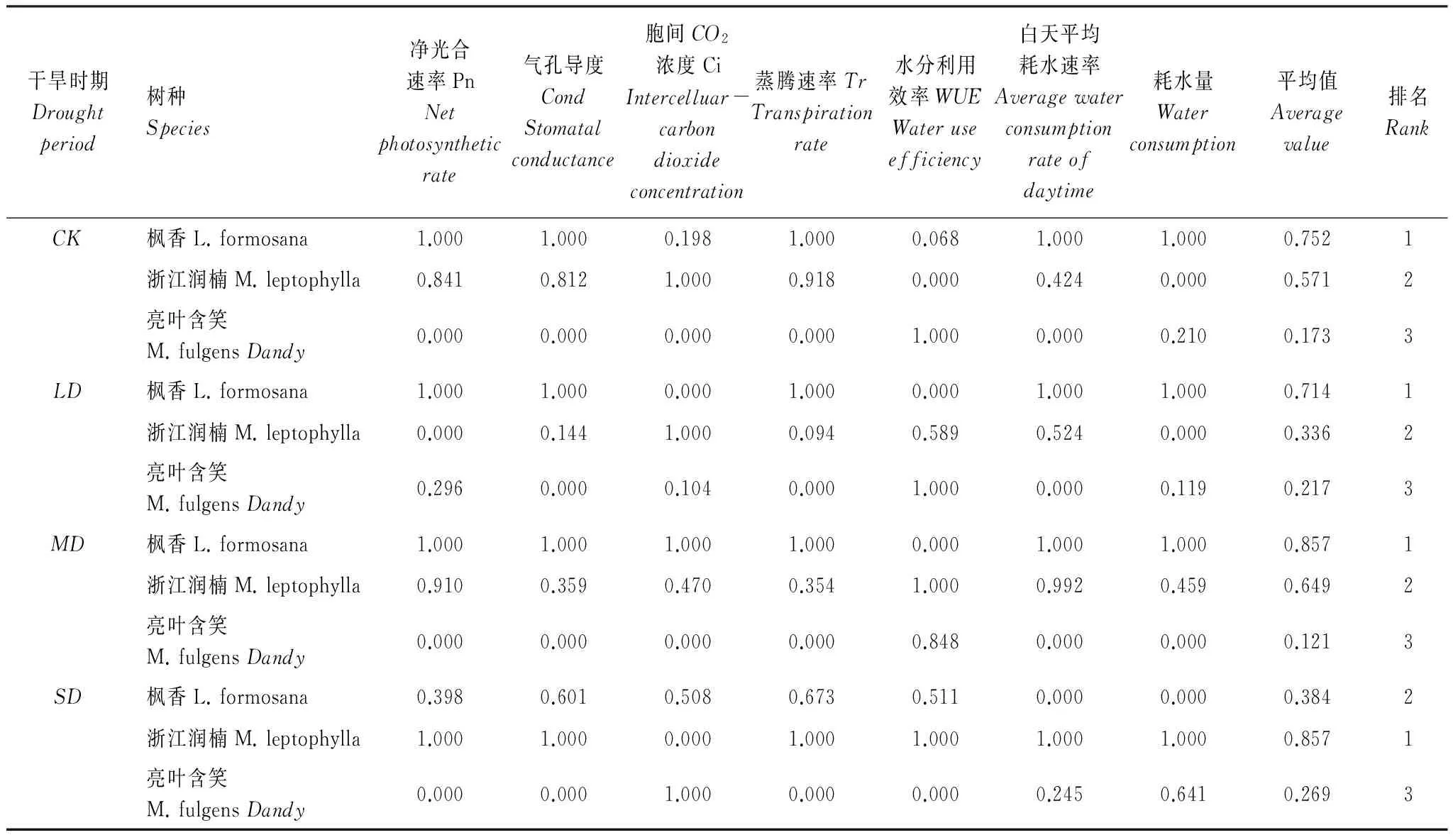
干旱时期Droughtperiod树种Species净光合速率PnNetphotosyntheticrate气孔导度CondStomatalconductance胞间CO2浓度CiIntercelluar-carbondioxideconcentration蒸腾速率TrTranspirationrate水分利用效率WUEWateruseefficiency白天平均耗水速率Averagewaterconsumptionrateofdaytime耗水量Waterconsumption平均值Averagevalue排名RankCK枫香L.formosana1.0001.0000.1981.0000.0681.0001.0000.7521浙江润楠M.leptophylla0.8410.8121.0000.9180.0000.4240.0000.5712亮叶含笑M.fulgensDandy0.0000.0000.0000.0001.0000.0000.2100.1733LD枫香L.formosana1.0001.0000.0001.0000.0001.0001.0000.7141浙江润楠M.leptophylla0.0000.1441.0000.0940.5890.5240.0000.3362亮叶含笑M.fulgensDandy0.2960.0000.1040.0001.0000.0000.1190.2173MD枫香L.formosana1.0001.0001.0001.0000.0001.0001.0000.8571浙江润楠M.leptophylla0.9100.3590.4700.3541.0000.9920.4590.6492亮叶含笑M.fulgensDandy0.0000.0000.0000.0000.8480.0000.0000.1213SD枫香L.formosana0.3980.6010.5080.6730.5110.0000.0000.3842浙江润楠M.leptophylla1.0001.0000.0001.0001.0001.0001.0000.8571亮叶含笑M.fulgensDandy0.0000.0001.0000.0000.0000.2450.6410.2693
整个干旱时期,3个树种白天耗水量占74%—92%,可见3个树种耗水主要发生在白天,因此可以将白天的耗水规律作为主要研究内容,这和欧洲云杉、竹柳、尾巨桉、毛白杨等大部分树种的研究结果类似[22,24,29]。干旱条件下气孔关闭是植物蒸腾下降的重要因素[31]。气孔关闭主要是因为环境因子的影响,对比不同水分下,环境因子湿度和温度的影响来看,到干旱后期,环境因子对耗水速率的影响有所减弱,可能是植物关闭大部分气孔,从而环境因子失去了对植物耗水特性的间接控制。
重度干旱时期,枫香的白天平均耗水速率只占正常条件下的10%,而浙江润楠占到CK的50.7%,枫香最大耗水速率只占CK的8.6%,而浙江润楠占到CK的52.9%。重度干旱时期,枫香的日耗水量只占CK的7.2%,而浙江润楠占CK的46%。这说明浙江润楠在严重干旱时期依然对水分需求较大,而枫香对水分需求较小。从耗水需求这个角度来说,3个树种的耗水需求在轻度干旱和中度干旱差异不大,而重度干旱时期水分极度缺乏,浙江润楠可能在干旱后期会因为得不到充足的水分而死亡。
石漠化地区造林,水分是主要限制因子,土层较薄,保水能力较差,所以造林最基本最核心的要求不是要高生产力而应该是有较高的成活率,轻度和中度干旱条件下,树种之间差异主要体现在生长,但面临重度干旱时候,很多树种的耐旱能力直接决定了造林成败。石漠化地区特殊的生态地理条件,研究的重心应该转向重度干旱下石漠化树种的表现。进一步开展石漠化树种干旱与死亡机理方面的研究,丰富和验证目前国际上关于干旱与植物死亡的相关假说[32- 35],进一步从机理上解释石漠化地区造林死亡与干旱之间的关系。
3.2干旱胁迫对石漠化树种光合特征影响
叶片光合速率(Pn)是光合作用中的一个重要指标,有研究表明,随着干旱胁迫的加剧,光合(Pn)随之下降[35]。轻度干旱胁迫下,植物自身渗透作用调节,光合下降不是很明显,而在中度和重度干旱下,光合作用下降明显[36]。有研究表明,抗旱性强的植物Pn降低幅度较抗旱性弱的小[37]。因此干旱胁迫下植物净光合速率变化可以作为重要的评价指标之一来衡量植物抗旱能力的大小[38]。
3种石漠化树种幼苗在轻度干旱下,Pn与对照相比,亮叶含笑下降幅度最小,枫香次之,浙江润楠下降幅度最大,说明在轻度干旱胁迫下,亮叶含笑的抗旱能力强于枫香和浙江润楠。中度胁迫下,亮叶含笑较对照降低最大,这与轻度干旱表现形成反差。重度干旱下,较对照降低最大的是亮叶含笑,其次是枫香和浙江润楠,说明重度干旱时候浙江润楠抗旱性最强,其次是枫香和亮叶含笑。故在野外条件栽培种植时候可以根据当地遭受水分胁迫程度来优先选择适合的树种,亮叶含笑和枫香较适合于轻度水分胁迫环境种植,却不适合于经常遭受严重水分胁迫地区种植。浙江润楠可以在水分极度匮乏情况下依然可以生长。
3.3气孔限制与非气孔限制
干旱条件下,植物的光合速率受到气孔和非气孔限制的双重影响[39- 40]。在轻度干旱胁迫下,光合速率的降低由于气孔限制(Cs)引起的,中度或是重度干旱下是由于非气孔因素引起的[41]。有学者提出[42],检查气孔是否是光合速率下降的原因,既要看光合气孔限制值的大小,又需要看Ci的变化。当光合速率下降时,如果Ci下降而Cs升高,说明光合速率降低主要是由于气孔限制。反过来,如果Ci升高而Cs下降说明叶肉细胞光合下降,是非气孔限制引起。本研究中,枫香和亮叶含笑Ci在轻度干旱胁迫下保持一个较低水平,可能是轻度干旱时期气孔关闭所致。随着干旱程度加剧,内部代谢紊乱,Ci浓度增大,说明在中后期,非气孔限制是光合下降的主要原因。裴斌[43]研究沙棘苗木在8个土壤水分梯度下的光合发现,土壤相对含水量在38.9%—70.5%范围内,随干旱胁迫加重,沙棘的净光合速率(Pn)、气孔导度(Gs)和胞间CO2浓度(Ci)明显下降,而气孔限制值(Ls)显著上升,即Pn下降主要是由气孔限制造成的;当RWC<38.9%时,干旱胁迫继续加剧,Pn和Ls都降低,而Ci显著上升,即Pn下降的主要原因已经转变为非气孔因素的限制。这一观点也得到类似研究的支持[40]。 所以光合下降原因是气孔限制还是非气孔限制的判断还取决于干旱胁迫的程度。
3.4干旱胁迫度水分利用效率的影响
植物水分利用效率一直是研究中比较关注的问题,了解植物的水分利用效率可以掌握植物生存策略,最终可以认为控制有限的水资源来获得最高的产量和经济效益。植物水分利用效率(WUE)也是国内外干旱、半干旱以及湿润地区农业、林业、生物学研究讨论的热点问题之一[30,44]。WUE涉及不同的尺度(叶片、单株、林分,群落)问题,但较多的研究还是关注于叶片上的水分利用效率,叶片上的水分利用效率可以揭示植物内在的耗水机制,可以更好的为植被的合理供水、用水提供相应的科学依据,有利于干旱区植被的恢复和保育方面的研究[45]。WUE是反应植物耐旱性的一个有效指标,表征植物有效利用水分的能力,一般来说,在相同条件下,水分利用效率越高的植物抗旱能力也越强[46]。
本研究中,枫香和浙江润楠水分利用效率随着干旱加剧而降低,亮叶含笑则是先升高后降低,说明3个石漠化树种可以调节WUE来适应不同的水分条件。另外,3个树种的不同表现特征说明了不同石漠化树种在水分利用效率调节上的种间差异。浙江润楠和亮叶含笑在轻度干旱和中度干旱下依旧保持着较高的水分利用效率,说明在轻度和中度干旱下亮叶含笑和浙江润楠具有较强的抗旱能力,然而亮叶含笑却在重度干旱下水分利用效率几乎降为零,浙江润楠依然维持着一个较高的水分利用效率,说明浙江润楠可以适应极度干旱环境。有学者研究发现欧洲云杉无性系WUE在轻度和中度干旱胁迫下上升[29],与亮叶含笑表现出类似的适应机制。
4结论
(1)随着土壤干旱的加剧,3个树种光合均有不同程度下降。枫香和亮叶含笑在轻度干旱下,Ci较低,光合作用降低的主要原因是气孔因素限制,中度和重度干旱胁迫下,Ci显著上升,降低的主要原因变为非气孔限制,浙江润楠表现不是很明显。在以后的研究中可以进一步探讨浙江润楠光合下降的原因。亮叶含笑和枫香适合种植于经常受轻度和中度水分胁迫地区,浙江润楠适合种植于易遭受重度干旱胁迫地区。
(2)不同土壤水分条件下,3个石漠化树种耗水量、耗水速率和耗水日变化均存在差异,不同树种之间响应程度也不同。随着土壤干旱胁迫的加剧,耗水速率受环境因子湿度和温度影响不显著,3个树种耗水速率峰值点响应也不同,说明重度干旱时候植物内部代谢受损。浙江润楠和亮叶含笑在轻度和中度干旱下维持相对较高的水分利用效率,表现出较强的抗旱能力。亮叶含笑却在重度干旱下水分利用效率几乎降为零无法适应较为严重的水分胁迫,而浙江润楠却有相对较高的水分利用效率可以适应极度干旱环境。
(3)隶属函数抗旱评价结果表明,在正常、轻度和重度干旱下抗旱能力均为枫香>浙江润楠>亮叶含笑,重度干旱下为浙江润楠>枫香>亮叶含笑。综合来看,浙江润楠和枫香是相对高光合和高水分利用效率树种。在将来可以加强不同水分条件下不同植物抗旱表现研究对比,因为不同地区遭受的水分胁迫程度不一样,进一步筛选适合该地区干旱特点的树种。同时也可以加强研究重度干旱下植物表现,探讨植物干旱死亡机理。
参考文献(References):
[1]张殿发, 王世杰, 周德全, 李瑞玲. 贵州省喀斯特地区土地石漠化的内动力作用机制. 水土保持通报, 2001, 21(4): 1- 5.
[2]冯汉华, 熊育久. 广东岩溶地区石漠化现状及其综合治理措施探讨. 中南林业调查规划, 2011, 30(1): 15- 19.
[3]王月海, 房用, 隋日光, 鲍竹芳. 山东石灰岩山地荒山植被恢复技术的研究. 水土保持研究, 2006, 13(4): 240- 242.
[4]DobrindtL,StrohH,IsselsteinJ,VidalS.Infected-notinfected:FactorsinfluencingtheabundanceoftheendophyteNeotyphodium loliiinmanagedgrasslands.Agriculture,Ecosystems&Environment, 2013, (175): 54- 59.
[5]LiuWJ,LiPJ,DuanWP,LiuWY.Dry-seasonwaterutilizationbytreesgrowingonthinkarstsoilsinaseasonaltropicalrainforestofXishuangbanna,SouthwestChina.Ecohydrology, 2014, 7(3): 927- 935.
[6]胡宁, 袁红, 蓝家程, 袁道先, 傅瓦利, 文志林. 岩溶石漠化区不同植被恢复模式土壤无机磷形态特征及影响因素. 生态学报, 2014, 34(24): 7393- 7402.
[7]张祝平, 何道泉, 敖惠修, 杨伟机, 伍辉民. 粤北石灰岩山地主要造林树种的生理生态学特性. 植物生态学与地植物学学报, 1993, 17(2): 133- 142.
[8]吴焕忠. 影响广东省石灰岩地区造林成活率的因素分析. 中南林业调查规划, 2002, 21(4): 19- 22.
[9]邱权, 何茜, 李吉跃, 苏艳, 潘昕. 干旱胁迫对华南地区三种苗木蒸腾耗水的影响. 广东林业科技, 2012, 28(1): 19- 24.
[10]吴永彬, 朱明, 黄川腾, 秦新生, 郑明轩, 莫慧芝, 徐瑞晶, 庄雪影. 13个阔叶树种在粤北石灰岩山地造林的早期生长表现. 广东林业科技, 2013, 29(2): 11- 16.
[11]薛立, 薛晔, 任向荣, 史小玲, 冯慧芳.PEG模拟干旱条件下尾叶桉和枫香苗木的生理响应. 生态环境学报, 2009, 18(2): 614- 620.
[12]冯慧芳, 薛立, 任向荣, 傅静丹, 郑卫国, 史小玲. 4种阔叶幼苗对PEG模拟干旱的生理响应. 生态学报, 2011, 31(2): 371- 382.
[13]吴俊文, 白晶晶, 何茜, 李吉跃, 邱权, 潘昕. 华南地区5种苗木叶水势与土壤含水量的关系. 福建林业科技, 2014, 41(3): 16- 21.
[14]邱权, 潘昕, 何茜, 李吉跃, 苏艳, 林雯. 华南地区3种苗木生长旺盛时期光合特性及蒸腾耗水日变化规律的比较. 华南农业大学学报, 2012, 33(4): 524- 528.
[15]潘昕, 邱权, 李吉跃, 苏艳, 何茜. 干旱胁迫下华南地区3种苗木渗透调节物质的动态变化. 华南农业大学学报, 2012, 33(4): 519- 523.
[16]韩刚, 赵忠. 不同土壤水分下4种沙生灌木的光合光响应特性. 生态学报, 2010, 30(15): 4019- 4026.
[17]潘昕, 邱权, 李吉跃, 王军辉, 何茜, 苏艳, 马建伟, 杜坤. 干旱胁迫对青藏高原6种植物生理指标的影响. 生态学报, 2014, 34(13): 3558- 3567.
[18]郭卫华, 李波, 张新时, 王仁卿. 水分胁迫对沙棘(Hippophae rhamnoides)和中间锦鸡儿(Caragana intermedia)蒸腾作用影响的比较. 生态学报, 2007, 27(10): 4132- 4140.
[19]岳广阳, 张铜会, 赵哈林, 牛丽, 刘新平, 黄刚. 科尔沁沙地黄柳和小叶锦鸡儿茎流及蒸腾特征. 生态学报, 2006, 26(10): 3205- 3213.
[20]李红丽, 董智, 丁国栋, 张国盛, 王林和, 郝云龙. 浑善达克沙地植物蒸腾特征的研究. 干旱区资源与环境, 2003, 17(5): 135- 140.
[21]郭连生, 田有亮. 9种针阔叶幼树的蒸腾速率、叶水势与环境因子关系的研究. 生态学报, 1992, 12(1): 47- 52.
[22]邱权, 潘昕, 李吉跃, 何茜, 苏艳, 林雯. 速生树种尾巨桉和竹柳幼苗耗水特性和水分利用效率. 生态学报, 2014, 34(6): 1401- 1410.
[23]何茜. 毛白杨抗旱节水优良无性系评价与筛选[D]. 北京: 北京林业大学, 2008.
[24]何茜, 李吉跃, 陈晓阳, 张志毅. 毛白杨不同无性系苗木耗水量及其昼夜分配. 华南农业大学学报, 2010, 31(1): 47- 50.
[25]周平, 李吉跃, 招礼军. 北方主要造林树种苗木蒸腾耗水特性研究. 北京林业大学学报, 2002, 24(S1): 50- 55.
[26]胡红玲, 张健, 万雪琴, 陈洪, 易万洋, 周永春. 巨桉与5种木本植物幼树的耗水特性及水分利用效率的比较. 生态学报, 2012, 32(12): 3873- 3882.
[27]华雷, 何茜, 李吉跃, 刘珊, 余斐. 桉树无性系和华南乡土树种秋枫苗木耗水特性的比较. 应用生态学报, 2014, 25(6): 1639- 1644.
[28]陈婧, 马履一, 段劼, 彭祚登, 贾黎明, 杨欣超, 杨腾. 不同土壤含水量下黄连木苗木生长及耗水特性研究. 中南林业科技大学学报, 2014, 34(3): 68- 72.
[29]林雯, 何茜, 苏艳, 李吉跃, 王军辉, 邱权. 干旱胁迫对欧洲云杉水分生理特征的影响. 西北农林科技大学学报: 自然科学版, 2014, 42(6): 69- 77.
[30]康博文, 侯琳, 王得祥, 刘建军, 韩黎明. 几种主要绿化树种苗木耗水特性的研究. 西北林学院学报, 2005, 20(1): 29- 33.
[31]黄颜梅, 张健, 罗承德. 树木抗旱性研究(综述). 四川农业大学学报, 1997, 15(1): 52- 57.
[32]董蕾, 李吉跃. 植物干旱胁迫下水分代谢、碳饥饿与死亡机理. 生态学报, 2013, 33(18): 5477- 5483.
[33]SevantoS,McdowellNG,DickmanLT,PangleR,PockmanWT.Howdotreesdie?Atestofthehydraulicfailureandcarbonstarvationhypotheses.Plant,Cell&Environment, 2014, 37(1): 153- 161.
[34]QuirkJ,McDowellNG,LeakeJR,HudsonPJ,BeerlingDJ.Increasedsusceptibilitytodrought-inducedmortalityinSequoia sempervirens (Cupressaceae)treesunderCenozoicatmosphericcarbondioxidestarvation.AmericanJournalofBotany, 2013, 100(3): 582- 591.
[35]WoodwardI.Tansleyreviews.NewPhytologist, 2004, 163(3): 453.
[36]曹慧, 兰彦平, 王孝威, 曹琴, 杜俊杰. 果树水分胁迫研究进展. 果树学报, 2001, 18(2): 110- 114.
[37]敖红, 张羽. 水分胁迫对云杉光合特性的影响. 植物研究, 2007, 27(4): 445- 448.
[38]李吉跃. 植物耐旱性及其机理. 北京林业大学学报, 1991, 13(3): 92- 100.
[39]姚庆群, 谢贵水. 干旱胁迫下光合作用的气孔与非气孔限制. 热带农业科学, 2005, 25(4): 84- 89.
[40]王孝威, 段艳红, 曹慧, 郑王义. 水分胁迫对短枝型果树光合作用的非气孔限制. 西北植物学报, 2003, 23(9): 1609- 1613.
[41]EarlHJ.Stomatalandnon-stomatalrestrictionstocarbonassimilationinsoybean(Glycine max)linesdifferinginwateruseefficiency.EnvironmentalandExperimentalBotany, 2002, 48(3): 237- 246.
[42]FarquharGD,SharkeyTD.Stomatalconductanceandphotosynthesis.AnnualReviewofPlantPhysiology, 1982, 33: 317- 345.
[43]裴斌, 张光灿, 张淑勇, 吴芹, 徐志强, 徐萍. 土壤干旱胁迫对沙棘叶片光合作用和抗氧化酶活性的影响. 生态学报, 2013, 33(5): 1386- 1396.
[44]RichardsRA,RebetzkeGJ,CondonAG,vanHerwaardenAF.Breedingopportunitiesforincreasingtheefficiencyofwateruseandcropyieldintemperatecereals.CropScience, 2002, 42(1): 111- 121.
[45]曹生奎, 冯起, 司建华, 常宗强, 卓玛错, 席海洋, 苏永红. 植物叶片水分利用效率研究综述. 生态学报, 2009, 29(7): 3882- 3892.
[46]SobradoMA.Relationofwatertransporttoleafgasexchangepropertiesinthreemangrovespecies.Trees, 2000, 14(5): 258- 262.
基金项目:高等学校博士学科点专项科研基金项目(20124404110007); 广东省林业科技创新专项资金项目(2012KJCX014-01)
收稿日期:2015- 05- 06;
修订日期:2015- 11- 30
*通讯作者
Corresponding author.E-mail: ljyymy@vip.sina.com
DOI:10.5846/stxb201505060937
PhotosyntheticandwaterconsumptionoftreespeciesutilizedforafforestationofrockydesertinGuangdongProvince
WUJunwen,LIUShan,LIJiyue*,HEQian,SUYan,QIUQuan,BAIJingjing,WANGYan,JIAPeng
College of Forestry and Landscape Architecture, South China Agricultural University, Guangzhou 510642,China
Abstract:In rocky regions affected by desertification, prominent problems of thin soil, destruction of arid vegetation, and ecological deterioration present a big challenge to afforestation. Artificial recovery of forests is a quick and effective approach to restore the ecology of a region. Pot seedlings weight and Li- 6400 photosynthetic system measuring method were adopted to study water consumption and photosynthetic characteristics under drought stress in Machilus leptophylla, Liquidambar formosana, and Michelia fulgens. The results were as follows: (1) The net photosynthetic rate (Pn), stomatal conductance (Gs), transpiration rate (Tr), and intercellular carbon dioxide concentration (Ci) declined in the three karst species. During severe drought, photosynthesis and water use efficiency of Machilus leptophylla were 2.5- and 89.6-fold higher than those of Liquidambar formosana and 1.9- and 26.3-fold higher those of Michelia fulgens, respectively. (2) Drought stress in the middle and late growth season decreased leaf Pn and Gs in Liquidambar formosana and Michelia fulgens but increased the Ci, suggesting that non-stomatal limitations contributed to the decrease in Pn. (3) The total water consumption and daily average water consumption rate of the three species showed significant differences (P < 0.05). The daytime water consumption was 74%—92% of the total daily water consumption. Machilus leptophylla showed a high demand for water with a relatively high water consumption rate that was 3.7- and 2.2-folds higher than that of Liquidambar formosana and Michelia fulgens. (4) Diurnal variation in water consumption rate showed a single peak curve in all three tree species, and the peak changed under different drought stress. (5) Over all, Machilus leptophylla and Liquidambar formosana are species with relatively high photosynthetic rate and water use efficiency. The subordinate function results showed the drought resistance ability in the following order: Liquidambar formosana > Machilus leptophylla > Michelia fulgens, under normal, mild, and severe drought stress conditions, and Machilus leptophylla > Liquidambar formosana > Michelia fulgens, under severe drought stress.
Key Words:rocky desertification afforestation tree species; drought stress; photosynthetic; water consumption
吴俊文, 刘珊, 李吉跃, 何茜, 苏艳, 邱权, 白晶晶, 王燕, 贾朋.干旱胁迫下广东石漠化地区造林树种光合和耗水特性.生态学报,2016,36(11):3429- 3440.
WuJW,LiuS,LiJY,HeQ,SuY,QiuQ,BaiJJ,WangY,JiaP.PhotosyntheticandwaterconsumptionoftreespeciesutilizedforafforestationofrockydesertinGuangdongProvince.ActaEcologicaSinica,2016,36(11):3429- 3440.
