沙地赤松光合及叶绿素a快相荧光动力学特性
2016-08-09安宇宁白雪峰
孟 鹏, 安宇宁, 白雪峰
辽宁省固沙造林研究所, 阜新 123000
沙地赤松光合及叶绿素a快相荧光动力学特性
孟鹏*, 安宇宁, 白雪峰
辽宁省固沙造林研究所, 阜新123000
摘要:沙地赤松在科尔沁沙地南缘区已有50a的引种历史,但对其生长表现还知之甚少,对其光合生理还一无所知。为深入了解其光合生理特性及机理,以同龄樟子松为对照,对沙地赤松成熟林分生长指标以及光合和叶绿素a快相荧光动力学特性进行了研究。结果表明:与樟子松相比,成林阶段沙地赤松具有较大的生长量、生物量,这与其较强的光合作用密切相关。沙地赤松最大净光合速率Pmax(10.376μmolCO2 m-2s-1)和平均净光合速率Pn(4.902μmolCO2 m-2s-1)均高于樟子松,且具有较高的光饱和点LSP和较低的光补偿点LCP。暗呼吸速率(Rd)较低,导致其光合效率(Pmax/Rd)较高。JIP-测定揭示了沙地赤松较强光合作用的内在机制,在O相至P相范围内,其相对可变荧光值(Vt)总体较低,说明其在电子传递过程中耗散能量较低,而用于光化学的能量较高。比活性参数进一步表明沙地赤松单位激发态面积反应中心数目(RC/CSo、RC/CSm)、吸收的光量(ABS/CSo、ABS/CSm)、被反应中心捕获的光量(TRo/CSo)、用于电子传递的能量均较高(ETo/CSo),也因此具有较高的性能指数,其PI(ABS/CSo/CSm)分别是樟子松的1.42、1.65和1.63倍。显然在沙地环境下沙地赤松生长表现和光合性能优于樟子松,研究结果为扩大该树种在沙地上的引种栽培提供了初步的理论依据。
关键词:科尔沁沙地;沙地赤松;光合特性;快相叶绿素荧光;动力学特性
以CO2气体交换为特征的光合作用是植物的重要生理过程之一,植物的光合性能反映着其对光能的吸收与转换能力,与生长和生产力水平密切相关。叶绿素荧光分析技术是用于光合作用机理研究的一种“内在探针”,与 “表观性”的气体交换指标相比,叶绿素荧光参数更具有反映“内在性”的特点[1]。通过高速连续激发光得到的叶绿素a快相荧光动力学曲线(OJIP曲线)能全面反映位于类囊体膜上的蛋白复合体光系统Ⅱ(PSⅡ)的光反应过程、效率和结构。典型的OJIP曲线有O、J、I、P等相,通过这些相点的荧光特征可对电子传递过程的量子效率等信息进行评估,实现无损条件下对植物光合性能的有效监测[2- 3],这种方法被称为JIP-测定[4]。JIP-测定具有方便、快速、灵敏的特点,活体离体均可测定,不受材料大小和形态限制[5]。以该技术为切入点研究一些松属树种光合作用内在机理已有报道,比如Busch等探讨了班克松(P. banksiana) 的能量耗散机制[6],Martínez-Ferri等探讨了地中海白松(P. halepensis)和腓尼基刺柏(Juniperus phoenicea)在激发能捕获效率及光抑制程度上的差异[7],Pollastrini等探讨了意大利松(P. pinea)在遭受胁迫的初期针叶内电子传递效率和光合性能[8],Porcar-Castell等研究了欧洲赤松(Pinus sylvestris,樟子松为其地理变种)春季光合作用恢复期的叶绿素荧光动力学特性[9]。沙地赤松(P.densiflora)由辽宁省固沙造林研究所(下文简称固沙所)于1965年开始陆续在科尔沁沙地南缘引种栽培,至今已有50年的引种历史,于2014年被辽宁省林木良种审定委员会审定为赤松优良新品种。20世纪90年代开始,章古台沙地个别地点引种樟子松林(P. sylvestrisvar.mongolica)就出现了衰退现象,遭受了松枯梢病,外在表现为从底轮枝开始针叶枯黄萎蔫,而同期相同地点引种的沙地赤松则未表现任何衰退特征,感病指数几乎为0[10]。2000年和2012年,这里个别地点樟子松林遭受了松毛虫、松沫蝉及松枯梢病危害,导致树势衰弱,而同龄沙地赤松仍表现良好,并可以在沙地上进行天然更新。已有学者开始了赤松光合特性的研究,但多为人工控制环境条件的盆栽苗期试验[11]。在光适应范围研究方面,HanQM等人较全面的测定了赤松一级枝在水平和垂直方向上的光梯度,从叶片发育过程中形态指标的改变揭示了成熟赤松具有较强的光适应性[12],但该研究没有从叶绿素荧光角度深入探讨光适应性的机理。综上,针对特殊自然生境下赤松成熟林分的研究较少,沙地赤松成熟林分的光合及叶绿素a快相荧光动力学特性研究还未见报道。
本研究以同龄樟子松为对照,在对成林阶段沙地赤松生长指标进行调查的基础上,采用光合及JIP-测定相结合的方法对比二者的光合指标,并深入了解沙地赤松的光合机理,为在沙地环境下推广应用该针叶树良种提供一些基础的理论依据。
1材料与方法
1.1研究区概况
位于辽宁省彰武县章古台镇固沙所三家子实验区,地处42°43′N,122°29′E,海拔226.5m,年均降水量为433mm,蒸发量1570mm,年均温6.7 ℃,无霜期154d,≥10 ℃有效积温2800—3200 ℃。
1.1.1试验林分
试验林分建于1976年4月上旬,通过引种栽植2年生实生苗形成了沙地赤松与樟子松两个同龄的相邻林分,林分间相距6.0m。两林分面积分别为2.7hm2(沙地赤松)和2.1hm2(樟子松),初植株行距为1.0m×3.0m,后经历了相同的抚育和营林措施,于2002年进行了一次间伐,株行距变化为2.0m×6.0m。此后由于樟子松遭到了病虫害,对受害严重的单株进行了清除,因此樟子松林密度(701株/hm2)略低于沙地赤松(803株/hm2)。栽植地点为平缓沙地,高程差不超过0.4m。
1.1.2土壤
试验林建立时,土壤为灰沙土,瘠薄干旱,腐殖质含量极低,pH值呈中性。土壤表层20—50cm为灰沙,其下埋有50—100cm不等的黑褐色沙土,最下面则为白沙,沙层厚30m左右。土壤地下水位为3.6m,0—50cm土层土壤含水率年变化很大,在4.0%—8.0%之间。
开展本次研究时,0—50cm土层土壤含水率年变化进一步加大,在1.5%—9.5%之间,土壤地下水位下降为5.0m。两林分间土壤水分条件没有显著差异。
1.1.3植被
试验林建立时该区植被稀疏,均为一般沙地常见的旱生植物,有的形成群落,有的则不规律的散生,物种包括黄柳(Salix gordejevii)、胡枝子(Lespedeza bicolor)、差巴嘎蒿(Artemisia halodendron)、拂子矛(Calamagrostis epigeios)、碱草(Elymus dahuricus)等。
开展本次研究时,该区植被类型有所增加,主要是以樟子松、油松(P.tabuliformis)、沙地赤松为主的固沙针叶林,林下有少量伴生植物,包括鼠李(Rhumus parvifolius)、榆树(Ulmus pumila)、桑树(Morus alba)、胡枝子、南蛇藤(Celastrus articulatus)、白蔹(Ampelopsis japonica)、锦鸡儿(Caragana microphylla)、山里红(Crataegus pinnatifida)、黄菠萝(Phellodenron amurense)、小叶朴(Celtes bungeana)、小叶杨(Populus simoni)等。
1.2生长量和生物量调查
于2013年8月上旬,在试验林内分树种各设置1块30m×30m标准地,对林分密度、树高、胸径等指标进行调查。在各自标准地内,根据平均胸径、平均树高,选择3株标准木。在测定完光合及叶绿素荧光指标后,于8月中旬将标准木伐倒,利用解析木法测定连年树高和胸径生长量,并计算连年单株材积生长量。采用分层切割法测定干、枝、叶、球果等地上生物量[13]。采用全挖法测定根生物量[14],为了避免细根的损失,挖了长8m、宽6m、深4m的坑,用自制的三齿耙十分小心的从根尖部向根基部追踪式挖掘,根部挖出后用自来水洗净,然后烘干测生物量。各器官生物量测定后,计算干叶比(干生物量与叶生物量比值)和根茎比(根生物量与地上生物量比值)。
1.3光合特性调查
测定时间为8月上旬。在每株所选标准木树冠阳面中部选择样枝并做标记,用Li- 6400光合测定系统,在不加任何人为环境控制的自然状态下测定两树种当年生针叶净光合速率(Pn)、蒸腾速率(Tr)、气孔导度(Gs)等指标。将被测针叶尽量摆满叶室,表面积由针叶长度和表面积形成的回归方程来确定[15]。测定时选择晴朗天气,测定3d,每天8:00开始测定,每2h测1次,18:00结束,每个时间点每株标准木重复测定3次。测定期内光合有效辐射(PAR)变化范围为52—1704μmolm-2s-1,均值为839.5μmolm-2s-1;气温变化范围为22—29℃,均值为26.7℃;相对湿度(RH)变化范围为45%—73%,均值为59.2%。由于测定枝距地面4m左右,所以测定时需将叶室架设于4m高铝合金人字梯上。水分利用效率(WUE)由Pn与Tr的比值求得。
光响应曲线参数的测定:在测定光合日变化的当日,于10:00叶片充分活化时进行光响应参数测定。测定时叶室水平放置。在正常大气CO2浓度、最适温度范围、RH(70±5)%条件下,利用Li- 6400光合测定系统自带的红蓝光源测定,光量子通量密度(PPFD)设置为2000、1800、1500、1200、1000、800、500、200、100、80、50、20、0μmolm-2s-1,设置最小等待时间为120s,最大等待时间为200s,测定不同PPFD下针叶的Pn。两个树种Pn与PPFD之间的关系可用SPSSV17.0软件Nonlinear模块,按下述经验方程进行拟合[16]:
(1)
式中,Pmax(μmolCO2m-2s-1)为最大净光合速率;α为弱光下光量子利用效率(即表观量子效率,AQY);C0为度量弱光下净光合速率趋近于零的指标。当PPFD为0时,Pn为负值,即是暗呼吸速率(Rd),通过公式(1)可以计算。通过适合性检验,若方程拟合效果良好,则可用下式计算光补偿点(LCP):
(2)
假定Pn达到Pmax的99%时的PPFD为光饱和点(LSP),则:
(3)
1.4叶绿素a快相荧光动力学指标测定
与光合指标同期测定。具体方法是于8:00 剪取光合测定时所选样枝附近见光良好、粗细均匀的当年生枝5条,迅速插入备好的水瓶中,并装入放有湿巾的黑塑料袋内,立即带回实验室,暗适应30min。然后用PocketPEA植物效率分析仪 (型号PPEA,HandstechInstrumentLtd. 英国)测定叶绿素a快相荧光的多相跃升,并读取相关参数[17]。当年生针叶平行排列在荧光夹中,完全覆盖4mm2的测试孔。测定光源为6 个发光二极管提供的波长为650nm的红光,光照强度为3500μmolm-2s-1的饱和光强,荧光信号的记录时程为1s,起始点为照光后10μs,每种针叶测定20 个重复。PocketPEA可以产生以时间为横坐标,原始荧光值(从最小荧光Fo至最大荧光Fm)为纵坐标的叶绿素快相荧光(OJIP)曲线。为对比树种间差异,克服原始OJIP曲线变异性较大的缺点,对原始曲线进行了标准化。本研究用Fm-Fo进行标准化,标准化的荧光信号数据用相对可变荧光(Vt)表示,Vt=(Ft-Fo) / (Fm-Fo),用Vt绘制的OJIP曲线O相的荧光强度均为0,P相的荧光强度均为1[18]。解析从OJIP曲线中得到的主要荧光参数的生理意义(表1)。

表1 叶绿素a快相荧光动力学曲线提取的参数及意义
ABS:吸收的光量Absorptionflux; RC:反应中心Reactioncenter; TR:捕获的光量Trappedenergyflux; ET:电子传递的能量Electrontransportflux; DI: 热耗散的能量Dissipatedenergyflux; CS: 激发态面积Excitedcrosssection; PI:性能指数Performanceindex
1.5叶绿素(Chl)含量测定
采用混合液法测定[19]。
1.6数据处理
所有数据均采用SPSSV17.0软件进行数理统计分析,对组间数据进行单因素方差分析(one-wayANOVA),采用Fisher的LSD法做多重比较分析;显著性水平设定为α=0.05。采用Excel2003制图。
2结果与分析
2.1沙地赤松与樟子松单株生长量及生物量对比2.1.1生长量
对标准地内沙地赤松和樟子松树高和胸径生长量进行对比(表2),发现沙地赤松树高较高,是樟子松的1.15倍;胸径较大,是樟子松的1.23倍。而且,无论是树高还是胸径指标,沙地赤松的变异系数均较小,表明沙地赤松林分生长离散度较小,单株间生长量更趋于一致。进行本次研究的当年及前一年,沙地赤松的连年单株材积生长量均显著高于樟子松。

表2 沙地赤松、樟子松生长量对比表
不同小写字母表示不同树种间差异显著;表中数据为平均值±标准差,在树高和胸径指标方面,沙地赤松n=69,樟子松n=74;连年材积生长量指标方面,两树种n=3
2.1.2生物量
表3所示,两树种生物量排序规律是一致的,即:干>根>枝>叶>果。沙地赤松总生物量较大,是同龄樟子松的1.73倍,其中干、果、根生物量显著高于樟子松,分别是樟子松的1.69、7.8和1.97倍,而枝、叶生物量与樟子松差异不显著。沙地赤松干叶比和根茎比均显著高于樟子松。
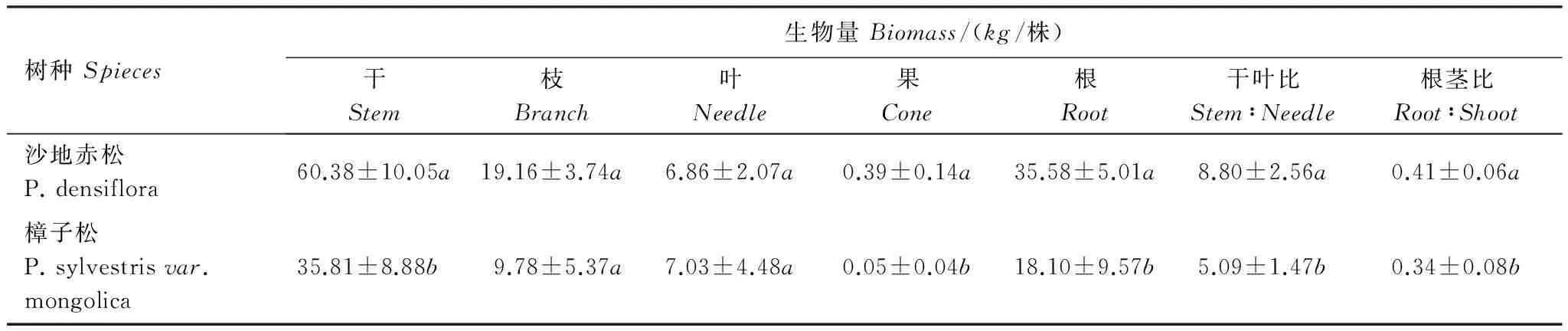
表3 沙地赤松、樟子松单株标准木各器官生物量(干重)对比表
不同小写字母表示不同树种间差异显著;表中数据为平均值±标准差,n=3
2.2光合特性对比2.2.1光合能力方面
光响应曲线反映了在其它环境因子相对稳定的条件下,植物Pn随PPFD变化的规律。利用公式(1)拟合出两树种的光响应曲线(图1右),拟合公式如下:
沙地赤松
樟子松
根据公式1—3计算出各光响应参数(表4)。Pmax/Rd的比值反应植物的光合效率,沙地赤松Pmax/Rd较高,是樟子松的1.74倍。沙地赤松Pmax较高,是樟子松的1.25倍,且Rd较低,导致Pmax/Rd较高。由光响应曲线可以看出,沙地赤松在几乎任意光强下Pn均大于樟子松(图1)。光合日变化可以看出,沙地赤松Pn日变化范围在3.290—7.349μmolCO2m-2s-1之间,平均Pn为4.902μmolCO2m-2s-1,比樟子松大36.2%,除14:00和16:00外其他时间点均显著超过樟子松(表5)。另外沙地赤松Tr比樟子松小34.8%,因此水分利用效率(WUE)显著高于樟子松,约是樟子松的2.06倍。并且,沙地赤松能在较小Gs下保持较高Pn(表6)。
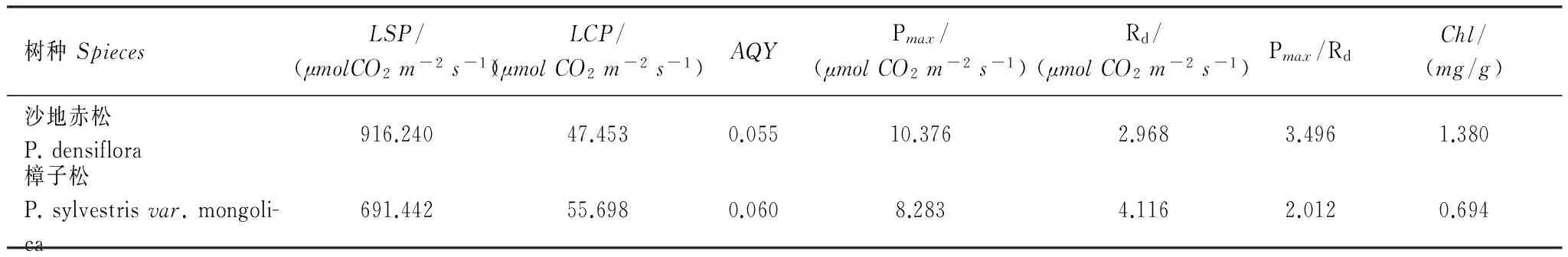
表4 两树种光响应曲线特征主要参数及叶绿素含量

图1 两树种利用公式(1)拟合前(a)和拟合后(b)的光响应曲线Fig.1 PPFD response curves of net photosynthetic rate (Pn) for the two species before (a) and after (b) fitted by equation (1)

树种TreespeciesPn/(μmolCO2m-2s-1)8:0010:0012:0014:0016:0018:00沙地赤松P.densiflora7.349±0.618a6.211±0.800a4.001±0.776a3.931±0.841a4.631±0.646a3.290±1.207a樟子松P.sylvestrisvar.mon-golica5.365±1.412b5.441±1.174b2.293±0.828b3.783±0.646a3.523±0.778a1.190±0.395b
表中数值平均值±样本标准差;每个时间点随机取20个数据,即n=20;a,b表示不同树种间差异显著

表6 沙地赤松与樟子松平均气孔导度、光合速率、蒸腾速率和水分利用效率比较
表中数值平均值±样本标准差;每个时间点随机取20个数据,所以n=120;a,b表示不同树种间差异显著
2.2.2光适应范围方面
沙地赤松具有更高的光饱和点(LSP)和更低的光补偿点(LCP)(表4)。通过实际光响应曲线(图1a),发现樟子松易于发生动态光抑制(dynamicphotoinhibition),在PPFD达1800μmolm-2s-1时,光合速率迅速下降,Pn为峰值的77.4%;而当PPFD达2000μmolm-2s-1时,Pn仅为峰值的30.5%。相比之下,沙地赤松在PPFD为1800μmolm-2s-1时,并未发生光抑制,当PPFD达2000μmolm-2s-1时,仅发生了轻微的光抑制,Pn为峰值的91.4%。本研究还发现沙地赤松针叶具有较高的Chl,比樟子松高1倍,达到1.380mg/g(表4)。
2.3 叶绿素a快相荧光动力学特征对比2.3.1特征位点比较

图2 沙地赤松与樟子松由Vt和ΔVt绘制的叶绿素快相荧光动力学曲线Fig.2 Fast chlorophyll fluorescence transients of P. densiflora and P. sylvestris var. mongolica based on Vt and ΔVtP1、P2分别为沙地赤松和樟子松的P相点
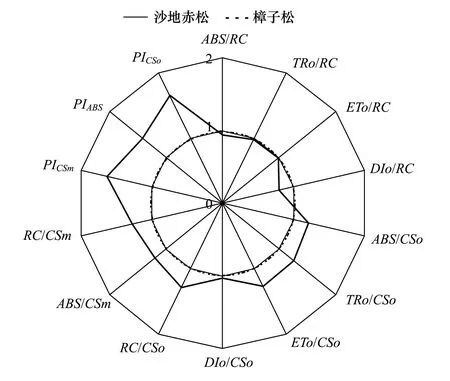
图3 沙地赤松与樟子松叶绿素a荧光比较 Fig.3 Radar plot of fluorescence data of P. densiflora and P. sylvestris var. mongolicaABS:吸收的光量Absorption flux; RC:反应中心 Reaction center; TR:捕获的光量Trapped energy flux; ET:电子传递的能量Electron transport flux; DI: 热耗散的能量Dissipated energy flux; CS: 激发态面积Excited cross section; PI:性能指数Performance index
由Vt绘制的OJIP曲线可以看出(图2),两树种均为典型的叶绿素快相荧光动力学曲线,具有O、J、I、P等相点。它们的曲线形态比较相似,但到达P相点的时间(Tfm)不相同,沙地赤松为700ms(P1位点),而樟子松为900ms(P2位点)。该图亦可看出,两树种针叶OJIP曲线的差异主要在J相(2ms)附近。为更清楚的观察包括J相在内各相荧光的差异,以樟子松Vt绘制的OJIP曲线为对照,然后用沙地赤松曲线与对照曲线的标准化荧光差值(ΔVt)重新作图(图2)。结果显示,从O相到接近P相的过程中,沙地赤松标准化荧光差值(ΔVt)大部分时间点均低于樟子松,两树种差异主要出现在300μs—30ms范围内,即J相附近区域,其中最大差异出现在J—I相之间(时间约为104μs)。在快接近P相时,沙地赤松标准化荧光值出现了高于樟子松的一个窄峰。
2.3.2荧光参数比较
比活性参数是指活跃的单位反应中心(RC)或单位受光面积(CS)的各种量子效率(ABS/RC、TRo/RC、ETo/RC、DIo/RC、ABS/CSo、TRo/CSo、ETo/CSo、DIo/CSo、ABS/CSm)以及单位面积上的反应中心的数量(RC/CSo、RC/CSm)(表1)。图3 是以樟子松的荧光参数作为对照(参数值为1),用沙地赤松的荧光参数与对照参数的比值绘制成的雷达图。由图3可见除ABS/RC与DIo/RC小于樟子松外,沙地赤松其它比活性参数指标均等于或大于樟子松,尤其是RC/CSo、RC/CSm、ABS/CSo、ABS/CSm、TRo/CSo、ETo/CSo显著高于樟子松。图3还表明,沙地赤松性能指数PI(AB/CSo/CSm)较高,分别是樟子松的1.42、1.65和1.63倍。
3讨论
固沙所从1965年开始就陆续在科尔沁沙地南缘地区开展了沙地赤松引种造林试验,目前沙地赤松林树势健康,生长稳定。通过本研究发现在同样立地条件下,虽然沙地赤松密度较大,单株营养面积和受光照条件不及樟子松,但其单株生长量和生物量均大于樟子松,尤其是在测定光合指标的当年及前一年的连年材积生长量显著大于樟子松,说明沙地赤松可以固定较多的CO2,并积累较多的光合产物用于生长。由于植物固定CO2的能力和其光合性能及吸收光能后的电子传递具有相关性[20],推断沙地赤松光合性能和光能利用效率可能好于樟子松。
为验证这一推论,本研究采用光合生理指标和叶绿素a快相荧光动力学指标相结合的方法,对沙地赤松光合水分生理特性进行了剖析。研究发现沙地赤松具有较强的光合能力,Pmax和平均Pn均高于樟子松,能在较小Gs下保持较高Pn。这意味着沙地赤松可以产生较多的光合产物。在光合产物分配方面,沙地赤松也具有自己的优势:干叶比较高,说明其把更多的光合产物积累到了树干中,也因此连年材积生长较高;根茎比较高,应归因于沙地赤松较强的光合能力导致地上部分能够向根系供给充足的碳氢化合物[21],用于了根系的横向和垂直向的生长[22],提高了自身的吸水能力和抗旱性,也因此更有利于适应土壤长期水分亏缺的沙地环境。沙地环境的另一个特点是多风,在章古台地区树木生长季常以南及西南风为主,最大瞬间风速32m/s,年5m/s以上的风可达240次。在这样的多风环境下,植物的蒸腾耗水量增加,极易使植物处于干旱胁迫之下。而沙地赤松能够保持较小Tr和较高的WUE,增加了其在沙地环境下长期生存生长的稳定性。另外暗呼吸速率(Rd)较低保证了沙地赤松对光合产物的低消耗[23],同时形成了较高的光合效率(Pmax/Rd)。与樟子松相比,沙地赤松具有更高的LSP,因此在较高光强(2000μmolm-2s-1)下仅发生轻微的光抑制;具有更低的LCP,说明其具有较强的利用弱光的能力,这和其较高的叶绿素含量(Chl)有关,较高的Chl有利于叶绿体对光能的吸收与转化,也保障了其较高的光合能力[24]。
前人研究表明,比活性参数可以更确切的反映植物的光合器官对光能的吸收、转化和耗散等状况[25],长期多次干旱胁迫会造成沙地植物在ETo/CSo等指标的下降,从而降低用于电子传递的能量[26]。本研究发现沙地赤松大部分比活性参数高于樟子松,包括RC/CSo、RC/CSm、ABS/CSo、ABS/CSm、TRo/ CSo、ETo/CSo等,表明其在沙地环境条件下单位激发态面积反应中心数目(RC/CSo、RC/CSm)、吸收的光量(ABS/CSo、ABS/CSm)、被反应中心捕获的光量(TRo/CSo)、用于电子传递的能量(ETo/CSo)均保持较高水平。性能指数(PI(ABS/CSo/CSm))包含了3 个参数[RC/CS、φPo(PSⅡ 最大光化学效率) 和ψo(表示电子越过QA的能量占反应中心捕获能量的比例)],这3 个参数相互独立[27],所以性能指数和推动力(即性能指数的对数)可以更准确地反映植物光合机构的状态[28]。沙地赤松性能指数PI(ABS/CSo/CSm)较高,证明沙地赤松光反应活性较强。沙地赤松光合机构的这些优良特性使其能在较小Gs下保持较高Pn,也使其在弱光下能够吸收更多的光能,导致较低的LCP;在强光下能利用较多的光能,从而避免激发能的积累,产生光抑制,保持较高的LSP。以上研究揭示了沙地赤松对光具有很强的适应性,也揭示了沙地赤松生长量和生物量均较大的内在生理机制。
在P相时荧光水平达到了最大值(Fm),Fm是在1s内获得的,它代表了PSⅡ反应中心的关闭,代表了PSⅡ和质体醌A(QA)所有原初电子受体的完全还原,及PSⅡ受体侧电子流的饱和[29]。在快接近P相时,沙地赤松标准化荧光值出现了高于樟子松的一个窄峰,表明电子传递到电子传递链的慢还原型质体醌(PQ)库的末端时耗散能量较大[30]。这说明沙地赤松并非所有的光合机构性能指标都好于樟子松,只是总体性能优于樟子松而已。
4结论
4.1与樟子松相比,成林阶段沙地赤松具有较大的生长量、生物量,这与其较强的光合作用密切相关。沙地赤松Pmax和平均Pn均高于樟子松,且光适应范围较大,具有较高的LSP和较低的LCP,暗呼吸速率(Rd)较低,光合效率(Pmax/Rd)较高。
4.2JIP-测定揭示了沙地赤松较强光合作用的内在机制,在O相至P相范围内,其相对可变荧光值(Vt)总体较低,说明其在电子传递过程中耗散能量较低,而用于光化学的能量较高。比活性参数进一步表明沙地赤松单位激发态面积反应中心数目、吸收的光量、被反应中心捕获的光量、用于电子传递的能量均较高,也因此具有较高的性能指数,其PI(ABS/CSo/CSm)分别是樟子松的1.42、1.65和1.63倍。
4.3在沙地环境下,沙地赤松生长表现和光合性能优于樟子松,研究结果为扩大该树种在沙地上的引种栽培提供了光合理论依据。
参考文献(References):
[1]KrauseGH,WeisF.Chlorophyllfluorescenceandphotosynthesis:thebasics.AnnualReviewofPlantPhysiologyandPlantMolecularBiology, 1991, 42: 313- 349.
[2]Demmig-AdamsB,AdamsWWIII.Theroleofxanthophyllcyclecarotenoidsintheprotectionofphotosynthesis.TrendsinPlantScience, 1996, 1(1): 21- 26.
[3]张谧, 王慧娟, 于长青. 超旱生植物沙冬青高温胁迫下的快速叶绿素荧光动力学特征. 生态环境学报, 2009, 18(6): 2272- 2277.
[4]MereuS,GerosaG,MarzuoliR,FusaroL,SalvatoriE,FincoA,SpanoD,ManesF.GasexchangeandJIP-testparametersoftwoMediterraneanmaquisspeciesareaffectedbyseasprayandozoneinteraction.EnvironmentalandExperimentalBotany, 2011, 73: 80- 88.
[5]StirbetA,Govindjee.OntherelationbetweentheKautskyeffect(chlorophyllafluorescenceinduction)andphotosystemⅡ:BasicsandapplicationsoftheOJIPfluorescencetransient.JournalofPhotochemistryandPhotobiologyB:Biology, 2011, 104(1/2): 236- 257.
[6]BuschF,HünerNPA,EnsmingerI.Increasedairtemperatureduringsimulatedautumnconditionsdoesnotincreasephotosyntheticcarbongainbutaffectsthedissipationofexcessenergyinseedlingsoftheevergreenconiferjackpine.PlantPhysiology, 2007, 143(3): 1242- 1251.
[7]Martínez-FerriE,BalaguerL,ValladaresF,ChicoJM,ManriqueE.Energydissipationindrought-avoidinganddrought-toleranttreespeciesatmiddayduringtheMediterraneansummer.TreePhysiology, 2000, 20(2): 131- 138.
[8]PollastriniM,LuchiN,MichelozziM,GerosaG,MarzuoliR,BussottiF,CaprettiP.EarlyphysiologicalresponsesofPinus pineaL.seedlingsinfectedbyHeterobasidionsp.pl.inanozone-enrichedatmosphericenvironment.TreePhysiology, 2015, 35(3): 331- 340.
[9]Porcar-CastellA,PfündelE,KorhonenJFJ,JuurolaE.AnewmonitoringPAMfluorometer(MONI-PAM)tostudytheshort-andlong-termacclimationofphotosystemIIinfieldconditions.PhotosynthesisResearch, 2008, 96(2): 173- 179.
[10]宋晓东, 陈江燕, 刘桂荣, 徐贵军, 李淑华. 樟子松枯梢病的侵染发生规律. 森林病虫通讯, 2000, (4): 14- 17.
[11]金永焕, 李敦求, 姜好相. 不同土壤水分对赤松光合作用与水分利用效率的影响研究. 中国生态农业学报, 2007, 15(1): 71- 74.
[12]HanQM,KawasakiT,KatahataS,MukaiY,ChibaY.HorizontalandverticalvariationsinphotosyntheticcapacityinaPinus densifloracrowninrelationtoleafnitrogenallocationandacclimationtoirradiance.TreePhysiology, 2003, 23(12): 851- 857.
[13]林思祖, 杨梅, 曹子林, 刘洪波, 陈艳. 不同强度人为干扰对马尾松地上部分生物量及生产力的影响. 西北植物学报, 2004, 24(3): 516- 522.
[14]RaufS,SadaqatHA.Effectsofvariedwaterregimesonrootlength,drymatterpartitioningandendogenousplantgrowthregulatorsinsunflower(Helianthus annuusL.).JournalofPlantInteractions, 2007, 2(1): 41- 51.
[15]MyersDA,ThomasRB,DeluciaEH.Photosyntheticcapacityofloblollypine(Pinus taedaL.)treesduringthefirstyearofcarbondioxideenrichmentinaforestecosystem.Plant,CellandEnvironment, 1999, 22(5): 473- 481.
[16]BassmanJH,ZwierJC.GasexchangecharacteristicsofPopulus trichocarpa, Populus deltoidesandPopulus trichocarpa × P. deltoidesclones.TreePhysiology, 1991, 8(2): 145- 159.
[17]PerveenS,ShahbazM,AshrafM.RegulationingasexchangeandquantumyieldofphotosystemⅡ (PSⅡ)insalt-stressedandnon-stressedwheatplantsraisedfromseedtreatedwithtriacontanol.PakistanJournalofBotany, 2010, 42(5): 3073- 3081.
[18]ZeliouK,ManetasY,PetropoulouY.TransientwinterleafreddeninginCistus creticuscharacterizesweak(stress-sensitive)individuals,yetanthocyaninscannotalleviatetheadverseeffectsonphotosynthesis.JournalofExperimentalBotany, 2009, 60(11): 3031- 3042.
[19]陈福明, 陈顺伟. 混合液法测定叶绿素含量的研究. 林业科技通讯, 1984, (2): 4- 8.
[20]MaxwellK,JohnsonGN.Chlorophyllfluorescence—apracticalguide.JournalofExperimentalBotany, 2000, 51(345): 659- 668.
[21]HossainMD,HanafiMM,SalehG,ForoughiM,BehmaramR,NooriZ.Growth,photosynthesisandbiomassallocationofdifferentkenaf(Hibiscus cannabinusL.)accessionsgrownonsandysoil.AustralianJournalofCropScience, 2012, 6(3): 480- 487.
[22]GregoryPJ.Rootsandthephysico-chemicalenvironment.In:GregoryPJ,ed.PlantRoots:Growth,Activity,andInteractionwithSoils.Oxford:BlackwellPublishing, 2006: 131- 173.
[23]HeimannM,ReichsteinM.Terrestrialecosystemcarbondynamicsandclimatefeedbacks.Nature, 2008, 451(7176): 289- 292.
[24]CayónSG.Theevolutionofphotosynthesis,transpirationandchlorophyllduringthedevelopmentofleavesofplantain(MusaAABSimmonds).Infomusa, 2001, 10(1): 12- 15.
[25]李鹏民, 高辉远,StrasserRJ. 快速叶绿素荧光诱导动力学分析在光合作用研究中的应用. 植物生理与分子生物学学报, 2005, 31(6): 559- 566.
[26]LuoYY,ZhaoXY,ZhangJH,LiYL,ZuoXA,SunDC.PhotosynthesisofDigitaria ciliarisduringrepeatedsoildroughtandrewatering.SciencesinColdandAridRegions, 2015, 7(1): 81- 87.
[27]StrasserRJ,Tsimilli-MichaelM,SrivastavaA.Analysisofthechlorophyllafluorescencetransient.In:PapageorgiouG,Govindjee,eds.AdvancesinPhotosynthesisandRespiration.Netherlands:Springer, 2004: 321- 362.
[28]vanHeerdenPDR,StrasserRJ,KrügerGHJ.ReductionofdarkchillingstressinN2-fixingsoybeanbynitrateasindicatedbychlorophyllafluorescencekinetics.PhysiologiaPlantarum, 2004, 121(2): 239- 249.
[29]SchanskerG,TóthSZ,StrasserRJ.MethylviologenanddibromothymoquinonetreatmentsofpealeavesrevealtheroleofphotosystemIintheChlafluorescenceriseOJIP.BiochimicaetBiophysicaActa(BBA) -Bioenergetics, 2005, 1706(3): 250- 261.
[30]Govindjee.Sixty-threeyearssinceKautsky:Chlorophyllafluorescence.AustralianJournalofPlantPhysiology, 1995, 22(2): 131- 160.
基金项目:林业公益性行业科研专项(201004023);辽宁省科学事业公益研究基金项目(GY2013- 13-011);辽宁省农业科技重点计划项目(2011207002,2011207004)
收稿日期:2015- 03- 23;
修订日期:2015- 11- 30
*通讯作者
Corresponding author.E-mail: mengpeng18@163.com
DOI:10.5846/stxb201503230552
PhotosyntheticcharacteristicsandchlorophyllafluorescenceinductionparametersofPinus densifloraonsandysoil
MENGPeng*,ANYuning,BAIXuefeng
Liaoning Province Sand-Fixation and Afforestation Research Institute, Fuxin 123000, China
Abstract:Pinus densiflora has grown in the southern Horqin Sandy Land, China, for 50 years, but little is known about its field performance in this region, and nothing is known about its photosynthetic physiology. To understand the physiological characteristics and mechanisms of photosynthesis in P. densiflora, mature trees were compared with the same-aged trees of Pinus sylvestris var. mongolica (the control), and the growth characteristics of both species were investigated. Photosynthetic characteristics were analyzed with a Li- 6400 system, and fast chlorophyll a fluorescence transients (OJIP) were analyzed with a Pocket PEA plant efficiency analyzer. Growth and biomass of mature P. densiflora was greater than that of P. sylvestris var. mongolica. This was closely related to stronger photosynthesis in P. densiflora, including a higher maximum net photosynthetic rate (Pmax, 10.376 μmol CO2 m-2s-1) and a large range of light adaptation with high light saturation point and low light compensation point. The dark respiration rate (Rd) of P. densiflora was low, leading to higher photosynthetic efficiency (Pmax/Rd). Diurnal changes in the net photosynthetic rate (Pn) of P. densiflora ranged from 3.290 to 7.349 μmol CO2 m-2s-1, and the mean Pn was 4.902 μmol CO2 m-2s-1, which was 36.2% higher than that of P. sylvestris var. mongolica. The Pn of P. densiflora at each time point was significantly higher than that of P. sylvestris var. mongolica, except at 14:00 and 16:00. The transpiration rate of P. densiflora was 34.8% lower than that of P. sylvestris var. mongolica, and its water use efficiency was 2.06-fold greater than that of P. sylvestris var. mongolica. The Pn of P. densiflora remained high when stomatal conductance was low. Compared with P. sylvestris var. mongolica, P. densiflora was less susceptible to dynamic photoinhibition, showing slight photoinhibition only at a PPFD of 2000 μmol m-2s-1, when Pn was 91.4% of the maximum. The chlorophyll content in P. densiflora needles was 1.380 mg/g, double that in needles of P. sylvestris var. mongolica. The JIP test was used to reveal the mechanism for the strong photosynthesis of P. densiflora and showed that in the range from O to P phase, the relative variable fluorescence value of P. densiflora was generally low. This low value indicated that little energy was dissipated via the electron transfer chain, so that more energy could be used for photochemistry. The two species reached fluorescence maxima at different times under saturating illumination: 700 ms for P. densiflora and 900 ms for P. sylvestris var. mongolica. The phenomenological energy fluxes per excited cross section (CS) were calculated; these indexes showed that, compared with P. sylvestris var. mongolica, P. densiflora had a higher density of reaction centers (RC/CSo, RC/CSm), a greater proportion of photon flux absorbed by the antenna pigments (ABS/CSo, ABS/CSm), higher trapping flux to the reaction center (TRo/CSo), and used more energy for electron transfer (ETo/CSo). These attributes led to its higher performance indexes. The PI(ABS/CSo/CSm)of P. densiflora were 1.42-, 1.65-, and 1.63-fold those of P. sylvestris var. mongolica, respectively. Together, these results suggest that the growth and photosynthetic performance of P. densiflora are better than those of P. sylvestris var. mongolica in the Horqin Sandy Land environment. This conclusion provides the theoretical basis for the expansion of P. densiflora cultivation in this region.
Key Words:Horqin Sandy Land; Pinus densiflora; photosynthetic characteristics; transient chlorophyll fluorescence; kinetics
孟鹏, 安宇宁, 白雪峰.沙地赤松光合及叶绿素a快相荧光动力学特性.生态学报,2016,36(11):3469- 3478.
MengP,AnYN,BaiXF.PhotosyntheticcharacteristicsandchlorophyllafluorescenceinductionparametersofPinus densifloraonsandysoil.ActaEcologicaSinica,2016,36(11):3469- 3478.
