荒漠绿洲区不同种植年限人工梭梭林土壤化学计量特征
2016-08-09苏永中刘婷娜
张 珂,苏永中,王 婷,刘婷娜
1 中国科学院寒区旱区环境与工程研究所临泽内陆河流域研究站,兰州 730000 2 中国科学院大学,北京 100049
曾清苹1,何丙辉1,*,毛巧芝1,秦华军2,李 源3,黄 祺4
1 西南大学资源环境学院,三峡库区生态环境教育部重点实验室,重庆 400715 2 贵州省生物研究所,贵阳 550009 3 重庆市涪陵区林业局,重庆 408000 4 中国电建集团贵阳勘测设计研究院有限公司,贵阳 550009
荒漠绿洲区不同种植年限人工梭梭林土壤化学计量特征
张珂1,2,苏永中1,*,王婷1,2,刘婷娜1
1 中国科学院寒区旱区环境与工程研究所临泽内陆河流域研究站,兰州730000 2 中国科学院大学,北京100049
摘要:为阐明梭梭建立对林下土壤养分化学计量特征的影响,分析了2、5、9、13、16、31、39a荒漠绿洲区梭梭林灌丛下和流沙区(0a)土壤有机碳(SOC)、全氮(TN)、全磷(TP)、碳酸钙(CaCO3)、速效磷(Available P)含量及其化学计量特征变化规律。结果显示:1)SOC和TN在9a后出现显著的成层化分布,而TP的这一特征相对滞后;不同土壤深度SOC、TN均随林龄增加而显著增加,而TP未表现出明显变化。2)C∶P和N∶P在9a后表现出明显的成层化分布且不同土层C∶P和N∶P随林龄增加显著增加,而C∶N保持相对稳定。3)较低含量的Available P在2a后即表现出0—5 cm含量大于5—20 cm且表层Available P和CaCO3随林龄增加而显著增加。而CaCO3∶Available P在不同林龄间并未发生显著变化。 4)随林龄增加,土壤表层风蚀可蚀性极显著降低且与土壤表层养分含量呈显著的负相关关系。以上结果表明,梭梭的建立提高了土壤SOC和TN含量且随林龄增加变化更显著,而变化较小的C∶N说明土壤氮的形成需要固定比例的碳。变化幅度较小的TP说明主要来源于岩石风化的磷素受时间作用的限制,而较低含量的Available P和变化幅度较小的CaCO3∶Available P则表明梭梭的生长受P的限制且有限的Available P易被CaCO3固定。另外,土壤养分含量与土壤风蚀可蚀性显著的负相关关系,进一步说明梭梭的建立改善了土壤质地,增加了土壤养分含量,这对荒漠绿洲区土壤恢复和植被建设有极大的促进和指导作用。
关键词:人工梭梭林;林龄;土壤化学计量特征;荒漠-绿洲区
生态化学计量学作为近年来对生态系统养分循环研究的一种重要工具,是分析多重化学元素(尤其是碳(C)、氮(N)、磷(P))平衡关系及其生态系统交互影响的理论依据[1- 3]。目前,生态化学计量学研究的重点主要集中在对植物叶片中C∶N∶P化学计量特征的研究,其中,C∶N和C∶P反映了植物在同化CO2同时对N、P的吸收利用情况,N∶P临界值则是判断土壤对植物养分供应状况的指标[4- 5]。作为影响植物养分特征的重要因子,土壤是一个比较复杂的生态系统,由于其不可移动性,且受植被类型、盖度、气候、地形、土壤母质、生物多样性及发育时间等因素的影响,使得土壤生态系统具有较高的空间异质性,且不同区域土壤养分循环特征也有显著差异[6-8]。如,在较高纬度易受风蚀影响的荒漠区,植物体生命活动不仅受水分短缺的影响,还受土壤贫瘠养分以及胁迫环境的限制,从而影响着植物叶片中的养分元素含量[9- 11]。因此,对不同区域不同植被类型土壤化学计量特征的研究显得尤为重要。
在干旱荒漠区建立人工固沙植被是生态恢复的重要措施之一[12]。研究表明,人工固沙林的建立能显著提高了土壤表层有机质和N、P等养分含量,且随着人工植被建立时间的延长土壤肥力显著改变[13- 15]。另外,人工植被对流沙的固定不仅为物种繁衍和生物多样性的恢复创造必要的环境条件,而且促进了土壤的形及空间异质性的改变,进一步影响着植被格局的变化[13]。自20世纪70年代以来,为保护绿洲环境及恢复由沙化导致的退化的植被-土壤系统,梭梭作为重要的人工固沙植被在河西走廊中段荒漠绿洲带被广泛的种植[16]。随着不同时间梭梭人工防护林的建立,形成了不同林龄的防护林系统并起到了极大的防风固沙作用[16]。现有的对梭梭的研究显示,当梭梭林龄大于10a时,由于很难利用到深达20 m的地下水及干旱区有限的降水,梭梭林出现自疏现象,这对梭梭在干旱区固沙可持续发展产生重要的影响[17-18]。然而,随梭梭林龄的增加,土壤养分特征如何变化?土壤状况是否发生显著改变?土壤状况的改变与土壤养分特征的关系如何?对上述问题的研究将丰富对梭梭的研究并对荒漠绿洲区人工梭梭林的建立和生态恢复提供一定的理论指导。
本研究以荒漠绿洲区不同种植年限梭梭林为研究对象,考虑到梭梭自身独特的泌盐特性及其他生物因子的共同作用导致土壤环境具有相对较高的盐分含量,而盐分离子中高含量的Ca2+又对土壤P的有效性有显著的影响[19-21],因此,本研究将比较不同种植年限梭梭灌丛下土壤有机碳(SOC)、全氮(TN)、全磷(TP)、速效磷(Available P)、碳酸钙(CaCO3)含量,分析不同林龄土壤化学计量特征及其影响因子,从而揭示荒漠-绿洲区土壤养分平衡和循环,为梭梭林在荒漠绿洲区的可持续发展、环境保护和生态恢复提供理论依据及参考资料。
1研究区概况
研究区位于黑河中游甘肃省临泽县中科院临泽内陆河流域研究站区(39°21′N,100°07′E,海拔1350 m)东北部,为近几十年来由绿洲向荒漠扩展的过渡区。绿洲外围与巴丹吉林沙漠南缘和戈壁荒漠相接。该区域为典型的荒漠气候,年均降雨量为117 mm且多集中在7—9月份,年蒸发量2390 mm,年均气温7.6℃,无霜期165 d。西北风向为主,年均风速为3.2 m/s,大风日数(> 17 m/s)为15 d,风沙活动主要集中在3—5月[16]。地下水深度为4—10 m。该区地带性土壤为灰棕漠土,由于受风沙长期侵袭,在绿洲边缘有大片的表层结构疏松且有机质含量很低的风沙土发育,极易发生风蚀。绿洲边缘主要荒漠植物种有红砂(Reaumuriasoongarica)、珍珠(Salsolapasserina)、泡泡刺(Nitrariatangutorum)、沙拐枣(Calligonummongolicum)等灌木半灌木和碱蓬(Suaedaprzewalskii)、沙米(Agriophyllumsquarrosum)、雾冰藜(Bassiadasyphylla)、白茎盐生草(Halogetonarachnoideus)等1年生草本植物[16,22]。

图1 不同林龄梭梭林采样点分布图Fig.1 Distribution of sampling sites of Haloxylon ammodendron in different plantation ages
2材料与方法
2.1植物-土壤样品采集
于2014年8月初在研究区选择通过植苗建立的2、 5、 9、 13、 16、 31、39年的梭梭林为研究对象(图1)。在每个年代梭梭林中分别建立3个10 m × 10 m的研究样地,每个样地间隔约10 m。在每个样地分别选择5株大小长势相似的梭梭 (表1),以每株梭梭根部为中心,在距每株梭梭根部5 cm处的4个方向(东、西、南、北)上采集0—5 cm和5—20 cm土壤样品并混合。同时,在流动沙丘处采集土壤样品作为参考(0a)。
2.2土壤养分测定
土壤样品于室内自然风干,研磨过100目筛制成供试样品。土壤有机碳(SOC)采用重铬酸解外加热法测定,土壤全碳(TC)和全氮(TN)用德国元素分析仪进行测定(a vario MACRO CUBE elementar,Germany),土壤全磷(P)和速效磷(Available P)分别采用H2SO4-H2O2消煮法和NaHCO3提取法。土壤CaCO3含量利用土壤无机碳含量转化而来,公式为CaCO3(mg/g) = (TC-SOC) × 8.33[23]。
2.3数据处理
土壤风蚀可蚀性结合了土壤物理和化学性质,利用公式EF(Erodible Fraction) (%) = 0.29 + 0.31Sa + 0.17Si + 0.33Sc-2.59OM-0.95CaCO3(R2= 0.67)来评价土壤对于风力引起的剥离和搬运的敏感程度,式中,Sa、Si、Sc和OM分别代表土壤砂粒含量、土壤粉粒含量、土壤黏粒含量和土壤有机质含量,所有指标均用%表示[24-25]。
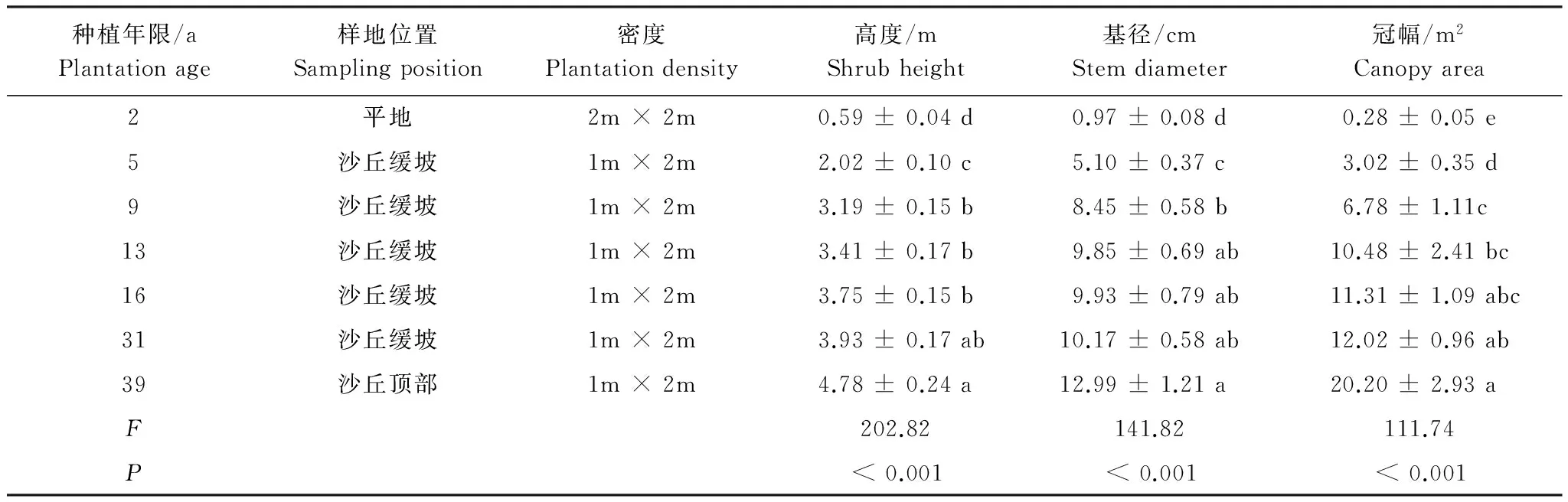
表1 不同林龄梭梭林形态特征
数据分析采用SPSS16.0软件进行分析,用ORINGIN 9.2做图。土壤SOC、TN、TP含量均采用质量含量,C∶N、C∶P、N∶P、CaCO3∶Available P采用质量比。用One-Sample T test 对0—5 cm和5—20 cm的土壤养分及化学计量比进行比较;利用One-Way ANOVA对不同林龄梭梭生长特征、土壤SOC、TN、TP、Available P、CaCO3、C∶N、C∶P、N∶P、CaCO3∶Available P和风蚀可蚀性的比较;SOC、TN和TP的相关性用Linear regression analysis进行分析; 利用Person product-moment correlation analysis对土壤变量和影响因子进行相关性分析。所有数据均经过K-S检验(One-Sample Kolmogorv-Smirnov Test)。
3结果与分析
3.1不同林龄土壤养分成层化分布特征
如表1所示,SOC和TN含量在5a之前随土壤深度增加无显著变化趋势(P>0.05),在5a之后表层(0—5 cm)SOC和TN含量则显著高于5—20 cm的含量(P<0.05)。TP含量在13a之后0—5cm含量显著高于5—20 cm含量(P<0.05)。表层Available P含量在2a之后表现为显著高于深层(P<0.05),而CaCO3含量并未随林龄增加表现出明显的成层化分布(P>0.05)。
土壤C∶N在2、16、31a均表现为0—5 cm显著高于5—20 cm(P<0.05),其余林龄下无显著差异(P>0.05)。垂直方向上C∶P和N∶P变化趋势一致,即在5a之后表层显著高于深层(P<0.05)。CaCO3∶Available P则在不同林龄处(除0a)均表现为深层比值显著高于表层(P<0.05)(表2)。
3.2不同林龄土壤养分水平分布化学计量特征
随梭梭种植年限的增加,水平方向上0—5 cm和5—20 cm的SOC和TN含量均表现为显著的增加(P<0.05),而TP含量并未随林龄的增加发生显著变化(P>0.05)。另外,Available P和CaCO3含量仅在土壤表层随林龄增加显著增加(P<0.05)且Available P的最大值(0.016 mg/g)仅占TP(0.36 mg/g)的5%(表2)。
水平方向上,随梭梭林龄的增加,土壤C∶N和CaCO3∶Available P在0—5 cm和5—20 cm均未有显著变化(P>0.05);C∶P和N∶P在表层则随林龄显著增加(P<0.05),5—20 cm处的C∶P和N∶P在31a之前均未有显著变化,而在39a处达到最大值(表2)。
3.3土壤养分相关性及其与影响因子的关系
分析不同林龄土壤SOC, TN及TP的相关关系发现,三者均呈显著的正相关关系(P<0.001)(图2)。Pearson相关分析表明,土壤pH与SOC,TN,C∶P和N∶P均呈极显著的正相关关系(P<0.01);土壤电导率(Electrical conductivity, EC)与SOC,TN, C∶P和N∶P均呈极显著的正相关关系(P<0.01),与TP和Available P呈显著的相关关系(P<0.05);除与CaCO3呈显著的负相关关系(P<0.05),与CaCO3∶Available P呈显著的正相关关系(P<0.05)及与C∶N无显著关系(P> 0.05)外,土壤砂粒与其他土壤变量均呈极显著的负相关关系(P<0.01);土壤粉粒和粘粒均与SOC,TN,TP,C∶P,N∶P呈极显著的正相关关系(P<0.01),与CaCO3呈显著的正相关关系(P<0.05),与CaCO3∶Available P呈显著的负相关关系(P<0.05)。另外,Available P 与SOC,TN和粉粒呈极显著的正相关关系(P<0.01),与粘粒呈显著正相关(P<0.05),且SOC与CaCO3,N∶P,和CaCO3∶Available P均有显著的相关关系(P<0.05)。总体来看,土壤砂粒、粉粒和粘粒是影响土壤养分及其化学计量特征的主要因子(表3)。
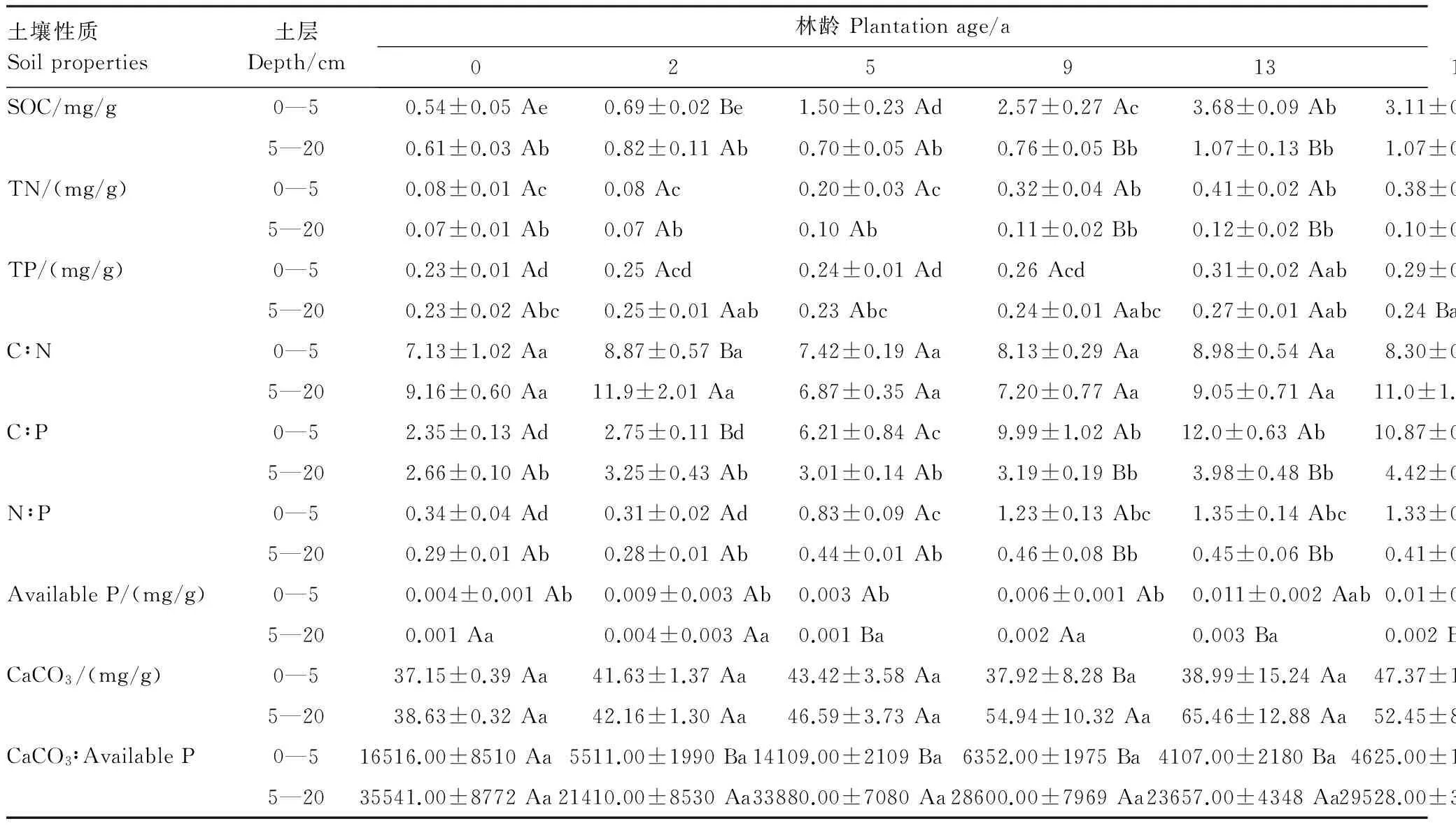
表2 不同林龄土壤养分成层化分布特征
同列同一元素不同大写字母表示差异显著(P<0.05);同行同一元素不同小写字母表示差异显著(P<0.05)
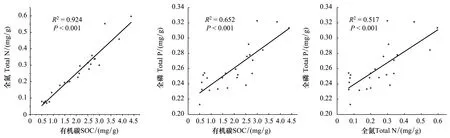
图2 土壤有机碳、全氮和全磷的关系Fig.2 Relationship between SOC, TN, and TP

变量Variable有机质SOC全氮TN全磷TP速效磷AvailableP碳酸钙CaCO3pH电导率EC砂粒Sand粉粒Silt粘粒Clay有机质SOC―0.678**0.817**-0.885**0.910**0.669**全氮TN―0.678**0.885**-0.900**0.909**0.700**全磷TP――0.3160.444*-0.625**0.687**0.416**速效磷AvailableP0.667**0.579**――0.2750.429*-0.678**0.726**0.476*碳酸钙CaCO30.406*0.3890.2960.354―0.3020.338-0.503*0.475*0.433*C∶N――0.1890.356-0.056-0.368-0.4030.176-0.185-0.127C∶P―0.952**――0.405*0.749**0.867**-0.905**0.895**0.727**N∶P0.933**―――0.3820.738**0.926**-0.907**0.883**0.746**CaCO3∶AvailableP-0.482*-0.404――― -0.234-0.3490.511*-0.444*-0.489*
**表示极显著相关(P<0.01); *表示显著相关(P<0.05); ― 表示存在自相关,不宜进行相关性分析
3.4土壤风蚀可蚀性与土壤养分的关系
结果表明,随林龄的增加,土壤表层(0—5 cm)的风蚀可蚀性极显著的减弱,并且表层SOC、TN、TP、Available P和CaCO3含量均随土壤风蚀可蚀性的减弱而显著增加(P≤ 0.001)(表4,图3)。而5—20 cm深度的土壤风蚀可蚀性呈显著的减弱(P<0.05)(表4),且仅与土壤CaCO3含量有关(表5)(P= 0.001)。

表4 土壤风蚀可蚀性(EF)随林龄变化特征
同列同一元素不同小写字母表示差异显著(P<0.05)
4讨论
植被-土壤是一个相互作用的过程,土壤SOC、N、P元素作为植物正常生长发育所必需的养分,在植物生长过程中发挥着重要的作用,其含量的多少及成分组合状况,均会受到土壤养分含量的影响,而植被的建立对土壤环境也有一定的反馈作用,如:植物凋落物的分解、根系分泌物及根际微生物的积累等,对土壤质地的改善和土壤养分的积累有极大的促进作用[26-29]。
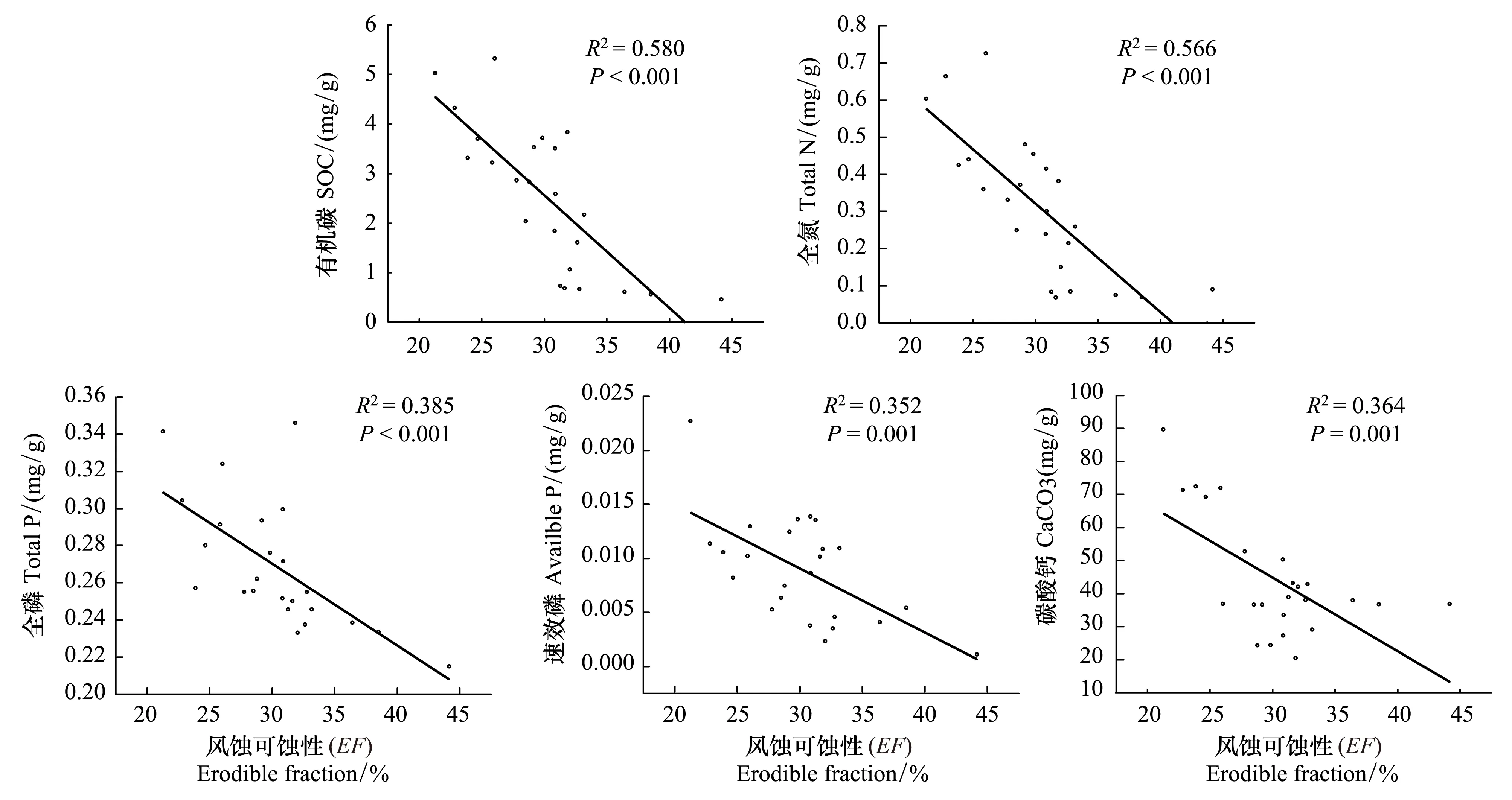
图3 土壤风蚀可蚀性 (0—5 cm) 与土壤养分 (0—5 cm) 的关系Fig.3 Relationship between soil erodible fraction with soil nutrients in 0—5 cm
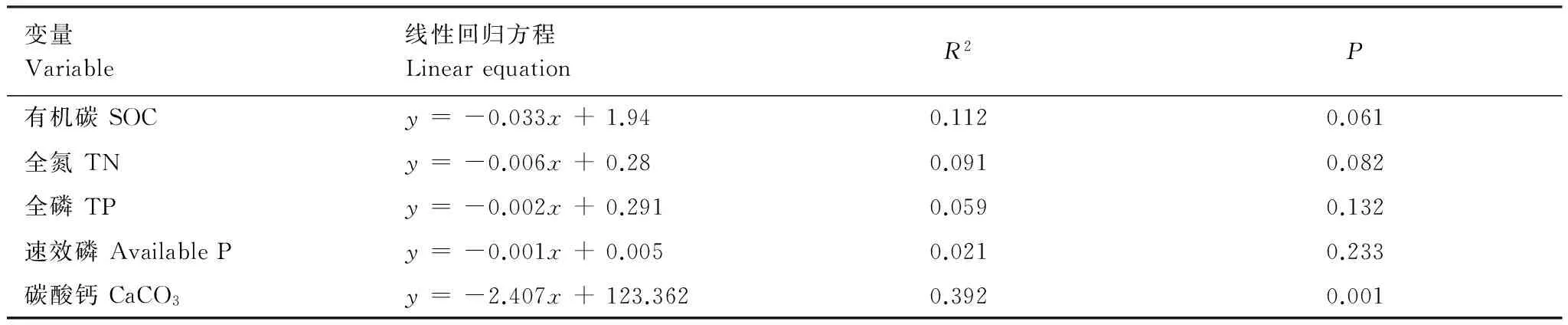
变量Variable线性回归方程LinearequationR2P有机碳SOCy=-0.033x+1.940.1120.061全氮TNy=-0.006x+0.280.0910.082全磷TPy=-0.002x+0.2910.0590.132速效磷AvailablePy=-0.001x+0.0050.0210.233碳酸钙CaCO3y=-2.407x+123.3620.3920.001
与流动沙丘相比,随着梭梭的建立,SOC和TN均显著增加并在梭梭建立5a后0—5 cm土壤SOC和N显著高于5—20 cm土壤,一方面说明梭梭建立后能够降低风速和减少地面的粗糙程度,不断的改善土壤表面的物理性质,为土壤养分的积累提供必要条件,并且随着梭梭林龄的增加,凋落物的不断增加直接影响着SOC和TN的积累;另一方面也说明了在受水分限制的干旱区土壤养分的表层积聚作用[30-32]。相对于SOC和TN,TP含量随梭梭林龄的增加没有显著变化且直到13a后0—5 cm TP含量才显著高于5—20 cm,这是由于TP的增加主要是通过来源相对固定且时间漫长的岩石风化作用,且表层土壤更易受岩石风化的影响,这就导致土壤P成层化分布的形成相对滞后[11]。 随林龄的增加,SOC和TN在0—5 cm和5—20 cm 均显著增加,而TP并未发生显著的变化,一方面说明梭梭的建立为SOC和TN的积累创造了有利的环境条件,而TP的变化则直接受土壤恢复时间的影响;另一方面表明梭梭的生长可能受P的限制。
C∶N∶P是衡量土壤有机质组成和营养平衡的一个重要指标,是确定土壤C、N、P平衡特征的重要参数[7,33]。与SOC和TN相比,土壤C∶N在不同深度和不同林龄间均维持相对稳定,这与Tian等[8]对中国土壤C∶N∶P的研究相一致[8],即SOC和TN含量具有较大的空间变异性,而受气候的影响较小的C∶N相对稳定,同时验证了不同深度土壤C∶N相对稳定的基本原则,即有机物质的形成需要一定数量的N与其相应的相对固定比率的C[2]。另外,相比于稳定的C∶N,土壤C∶P和N∶P随林龄的增加而显著增加,这与Tian等[8]、Cleveland等[7]分别从区域尺度和全球尺度上探讨的天然生态系统土壤C∶N、C∶P和N∶P化学计量比得出的结论一致,即:由于C元素和P元素来源的差异性,土壤C∶P和N∶P随气候、土壤类型、土壤深度和风化阶段而呈现显著的差异性。同时表明天然生态系统土壤C∶N、C∶P和N∶P的变化特征也适用于研究区人工梭梭林土壤。
对C、N和P的相关性分析显示三者间均有极显著的相关性,且C和N、C和P、N和P的相关系数分别为0.92、0.65和0.52。这与Tian等[8]对中国土壤8000个土壤层C、N和P的相关分析结果(相关系数分别为0.93、0.62和0.51)相似,这说明相对于大尺度上有机质丰富的土壤,人工梭梭林建立后SOC含量显著增加,且逐渐恢复的土壤中C、N和P也存在显著的耦合关系,这与最近的研究相符[34-35],表明小尺度上研究C、N和P的耦合关系也具有重要的生态指示作用。另外,人工梭梭林建立后土壤养分及化学计量特征与土壤砂粒、粉粒和粘粒显著的相关关系,进一步表明土壤质地的改善促进了土壤养分含量的提高[14]。
由于钙离子在植物抗旱性中起重要作用,同时速效磷是植物直接可吸收利用的磷素直接影响着植物体蛋白质的合成[1-2,36-37],因此,土壤CaCO3和速效磷对植物生长有重要的影响。然而,研究表明[38],土壤中高含量的CaCO3会大大降低土壤磷的有效性,进而影响植物生长。并且衡量土壤碳酸钙对速效磷固持指标的土壤碳酸钙/速效磷(钙磷比)化学计量特征对植物生物量、密度变化的影响显著大于土壤碳酸钙、速效磷含量的影响。本研究中,随林龄增加,表层Available P显著增加且在0—5 cm和5—20 cm间表现出明显的成层化现象,而Available P的最大值(0.016 mg/g)仅占总磷 (0.36 mg/g)的5%,说明尽管梭梭的建立提高了土壤Available P的含量,而较低的Available P含量进一步说明梭梭的生长受P的限制。此外,CaCO3在表层随林龄的增加而增加,而CaCO3∶Available P随林龄增加并未发生显著变化,说明土壤CaCO3含量与Available P增加的同步性,从而使得土壤中有限的速效磷被吸附固持。
土壤风蚀是导致干旱半干旱区土壤退化的一个普遍存在的现象[39]。土壤风蚀可蚀性临界值显示[24-25],EF<40%表明研究区较弱的风蚀可行性,40%
本研究在一定程度上揭示了梭梭建立对荒漠绿洲区土壤恢复的促进作用,为该区生态恢复提供了理论依据和参考。然而,植被-土壤是一个交互作用的复杂生态系统,因此,梭梭如何在有限的水分条件下实现可持续发展、梭梭群落特征及草本物种如何响应土壤环境变化将是今后的研究重点。
参考文献(References):
[1]Elser J J, Sterner R W, Gorokhova E, Fagan W F, Markow T A, Cotner J B, Harrison J F, Hobbie S E, Odell G M, Weider L J. Biological stoichiometry from genes to ecosystems. Ecology Letters, 2000, 3(6): 540- 550.
[2]Sterner R W, Elser J J. Ecological stoichiometry: the biology of elements from molecules to the biosphere. Princeton, New Jersey, USA∶Princeton University Press, 2002.
[3]Ågren G I. The C∶N∶P stoichiometry of autotrophs-theory and observations. Ecology Letters, 2004, 7(3): 158- 191.
[4]Koerselman W, Meuleman A F M. The vegetation N∶P ratio: a new tool to detect the nature of nutrient limitation. The Journal of Applied Ecology, 1996, 33(6): 1441- 1450.
[5]Rong Q Q, Liu J T, Cai Y P, Lu Z H, Zhao Z Z, Yue W C, Xia J B. Leaf carbon, nitrogen and phosphorus stoichiometry ofTamarixchinensisLour in the Laizhou Bay coastal wetland, China. Ecological Engineering, 2015, 76: 57- 65.
[6]Chadwick O A, Derry L A, Vitousek P M, Huebert B J, Hedin L O. Changing sources of nutrients during four million years of ecosystem development. Nature, 1999, 397(6719): 491- 497.
[7]Cleveland C C, Liptzin D. C∶N∶P stoichiometry in soil: is there a “Redfield ratio” for the microbial biomass? Biogeochemistry, 2007, 85(3): 235- 252.
[8]Tian H Q, Chen G S, Zhang C, Melillo J M, Hall C A S. Pattern and variation of C∶N∶P ratios in China′s soils: a synthesis of observational data. Biogeochemistry, 2010, 98(1/3): 139- 151.
[9]He M Z, Dijkstra F A. Drought effect on plant nitrogen and phosphorus: a meta-analysis. New Phytologist, 2014, 204(4): 924- 931.
[10]Drenovsky R E, Richards J H. Critical N∶P values∶Predicting nutrient deficiencies in desert shrublands. Plant and Soil, 2004, 259(1/2): 59- 69.
[11]Delgado-Baquerizo M, Maestre F T, Gallardo A, Bowker M A, Wallenstein M D, Quero J L, Ochoa V, Gozalo B, Garcia-Gómez M, Soliveres S, García-Palacios P, Berdugo M, Valencia E, Escolar C, Arredondo T, Barraza-Zepeda C, Bran D, Carreira J A, Chaieb M, Conceição A A, Derak M, Eldridge D J, Escudero A, Espinosa C I, Gaitán J, Gabriel Gatica M, Gómez-González S, Guzman E, Gutiérrez J R, Florentino A, Hepper E, Hernández R M, Huber-Sannwald E, Jankju M, Liu J S, Mau R I, Miriti M, Monerris J, Naseri K, Noumi Z, Polo V, Prina A, Pucheta E, Ramírez E, Ramírez-Collantes D A, Romão R, Tighe M, Torres D, Torres-Díaz C, Ungar E D, Val J, Wamiti W, Wang D L, Zaady E. Decoupling of soil nutrient cycles as a function of aridity in global drylands. Nature, 2013, 502(7473): 672- 676.
[12]李新荣, 赵洋, 回嵘, 苏洁琼, 高艳红. 中国干旱区恢复生态学研究进展及趋势评述. 地理科学进展, 2014, 33(11): 1435- 1443.
[13]李新荣. 干旱沙区土壤空间异质性变化对植被恢复的影响. 中国科学(D辑:地球科学), 2005, 35(4): 361- 370.
[14]苏永中, 赵哈林, 张铜会, 李玉霖. 科尔沁沙地不同年代小叶锦鸡儿人工林植物群落特征及其土壤特性. 植物生态学报, 2004, 28(1): 93- 100.
[15]翁伯琦, 郑祥洲, 丁洪, 王煌平. 植被恢复对土壤碳氮循环的影响研究进展. 应用生态学报, 2013, 24(12): 3610- 3616.
[16]Su Y Z, Zhao W Z, Su P X, Zhang Z H, Wang T, Ram R. Ecological effects of desertification control and desertified land reclamation in an oasis-desert ecotone in an arid region: A case study in Hexi Corridor, northwest China. Ecological Engineering, 2007, 29(2): 117- 124.
[17]Ma Q L, Wang J H, Zhu S J. Effects of precipitation, soil water content and soil crust on artificialHaloxylonammodendronforest. Acta Ecologica Sinica, 2007, 27(12): 5057- 5067.
[18]Zhu Y J, Jia Z Q. Soil water utilization characteristics ofHaloxylonammodendronplantation with different age during summer. Acta Ecologica Sinica, 2011, 31(6): 341- 346.
[19]Song J, Feng G, Tian C Y, Zhang F S. Strategies for adaptation ofSuaedaphysophora,HaloxylonammodendronandHaloxylonpersicumto a saline environment during seed-germination stage. Annals of Botany, 2005, 96(3): 399- 405.
[20]von Wandruszka R. Phosphorus retention in calcareous soils and the effect of organic matter on its mobility. Geochemical Transactions, 2006, 7: 6.
[21]Cross A F, Schlesinger W H. Biological and geochemical controls on phosphorus fractions in semiarid soils. Biogeochemistry, 2001, 52(2): 155- 172.
[22]陈隆亨, 李福兴, 邸醒民, 张继贤. 中国风沙土. 北京: 科学出版社, 1998.
[23]鲍士旦. 土壤农化分析(第三版). 北京: 中国农业出版社, 2000.
[24]Borrelli P, Ballabio C, Panagos P, Montanarella L. Wind erosion susceptibility of European soils. Geoderma, 2014, 232- 234: 471- 478.
[25]Fryrear D W, Krammers C A, Williamson D L, Zobeck T M. Computing the wind erodible fraction of soils. Journal of Soil and Water Conservation, 1994, 49(2): 183- 188.
[26]McGroddy M E, Daufresne T, Hedin L O. Scaling of C∶N∶P stoichiometry in forests worldwide: Implications of terrestrial Redfield-type ratios. Ecology, 2004, 85(9): 2390- 2401.
[27]贾晓红, 李新荣, 王新平, 樊恒文, 赵金龙. 流沙固定过程中土壤性质变异初步研究. 水土保持学报, 2003, 17(4): 46- 50.
[28]苏永中, 赵哈林, 张铜会. 几种灌木、半灌木对沙地土壤肥力影响机制的研究. 应用生态学报, 2002, 13(7): 802- 806.
[29]李从娟, 雷加强, 高培, 徐新文, 王永东, 范敬龙. 人工防护林作用下风沙土成土过程的研究进展. 土壤学报, 2012, 49(6): 1227- 1234.
[30]Jobbágy E G, Jackson R B. The distribution of soil nutrients with depth: Global patterns and the imprint of plants. Biogeochemistry, 2001, 53(1): 51- 77.
[31]He M Z, Dijkstra F A, Zhang K, Li X R, Tan H J, Gao Y H, Li G. Leaf nitrogen and phosphorus of temperate desert plants in response to climate and soil nutrient availability. Scientific Reports, 2014, 4: 6923.
[32]Schlesinger W H, Reynolds J F, Cunningham G L, Huenneke L F, Jarrell W M, Virginia R A, Whitford W G. Biological feedbacks in global desertification. Science, 1990, 247(4946): 1043- 1048.
[33]杨阳, 刘秉儒, 杨新国, 韩丛丛. 荒漠草原中不同密度人工柠条灌丛土壤化学计量特征. 水土保持通报, 2014, 34(5): 67- 73.
[34]王维奇, 王纯, 曾从盛, 仝川. 闽江河口不同河段芦苇湿地土壤碳氮磷生态化学计量学特征. 生态学报, 2012, 32(13): 4087- 4093.
[35]周正虎, 王传宽, 张全智. 土地利用变化对东北温带幼龄林土壤碳氮磷含量及其化学计量特征的影响. 生态学报, 2015, 35(20): 6694- 6702.
[36]Larkindale J, Knight M R. Protection against heat stress-induced oxidative damage in Arabidopsis involves calcium, abscisic acid, ethylene, and salicylic acid. Plant Physiology, 2002, 128(2): 682- 695.
[37]Kawano T, Sahashi N, Takahashi K, Uozumi N, Muto S. Salicylic acid induces extracellular superoxide generation followed by an increase in cytosolic calcium ion in tobacco suspension culture: The earliest events in salicylic acid signal transduction. Plant and Cell Physiology, 1998, 39(7): 721- 730.
[38]赵雪莱, 何兴东, 薛苹苹, 张宁, 邬畏, 李荣, 慈华聪, 徐静静, 高玉葆, 赵哈林. 土壤碳酸钙/有效磷化学
[39]南岭, 杜灵通, 展秀丽. 土壤风蚀可蚀性研究进展. 土壤, 2014, 46(2): 204- 211.
重庆缙云山两种林分土壤呼吸对模拟氮沉降的季节响应差异性
基金项目:国家自然科学基金项目(41271291);国家林业局“948”项目(2015- 4- 42);重庆市林业重点科技攻关项目(渝林科研2015- 7);重庆市应用开发计划重点项目(CSTC2014yykfB80015);西南大学资源环境学院“光炯”创新实验项目(2计量特征对油蒿群落植物密度的影响. 科学通报, 2012, 57(1): 80- 87.
0150104)
曾清苹,何丙辉,毛巧芝,秦华军,李源,黄祺.重庆缙云山两种林分土壤呼吸对模拟氮沉降的季节响应差异性.生态学报,2016,36(11):3244- 3252.
Zeng Q P, He B H, Mao Q Z, Qin H J, Li Y, Huang Q.Seasonal responses of soil respiration to simulated nitrogen deposition in a citrus plantation and masson pine forest in Mt. Jinyun, Chongqing, China.Acta Ecologica Sinica,2016,36(11):3244- 3252.
重庆缙云山两种林分土壤呼吸对模拟氮沉降的季节响应差异性
曾清苹1,何丙辉1,*,毛巧芝1,秦华军2,李源3,黄祺4
1 西南大学资源环境学院,三峡库区生态环境教育部重点实验室,重庆400715 2 贵州省生物研究所,贵阳550009 3 重庆市涪陵区林业局,重庆408000 4 中国电建集团贵阳勘测设计研究院有限公司,贵阳550009
摘要:氮沉降对土壤呼吸的影响仍然存在着争论,需要进一步研究。选择重庆缙云山的马尾松林和柑橘林开展了氮添加实验,分别设置3个氮添加水平(低氮T5:20 gN m-2a-1,中氮T10:40 gN m-2a-1和高氮T15:60 gN m-2a-1)和对照(T0:0 gN m-2a-1)共4个水平的处理,各林分每个处理各9次重复,每个处理量分4次,在每个季度开始各施1次。采用ACE(Automated Soil CO2Exchange Station, UK)自动土壤呼吸监测系统测定两林分土壤表层(0—10 cm)的呼吸、温度和湿度,分别在当年的7月、9月、11月、第2年的1月、2月、3月、5月、6月各连续测定4d,每天(8:00—18:00)4次,以揭示两种林分土壤呼吸对模拟氮沉降的季节动态响应及其差异性。结果表明:(1)柑橘林与马尾松林林下土壤表层呼吸表现出一致的季节变化动态趋势:夏季>春季>秋季>冬季,但柑橘林土壤呼吸显著高于马尾松林(P<0.05)。(2)总体上氮沉降抑制了2种林分土壤表层呼吸,而N沉降量大抑制程度越高。只在冬季土壤湿度低的马尾松林下氮沉降促进了土壤呼吸。(3)土壤温度与土壤呼吸有极显著的正相关指数关系(P<0.01),而土壤水分与土壤呼吸有显著的二次模型拟合关系,但均受到氮沉降量处理的影响。综合分析表明,在亚热带山区2类森林下的典型案例结果支持氮沉降抑制土壤呼吸的认识。
关键词:氮沉降;土壤呼吸;土壤温度;土壤湿度;柑橘林;马尾松林
Abstract: Disagreements over the effects of nitrogen deposition on soil respiration still exist, thereby necessitating further studies. In the present study, we designed an experiment to explore the seasonal responses of soil respiration to various levels of nitrogen deposition (T0: 0 g N m-2a-1as the control; T5: 5 g N m-2a-1as the low N treatment; T10: 10 g N m-2a-1as the moderate nitrogen treatment; and T15: 15 g N m-2a-1as the high nitrogen treatment. All treatments were performed with nine replicates both in the Masson pine forest and in the Citrus plantation at Mt. Jinyun, Chongqing, China. The different levels of N addition were applied four times at the beginning of each season from May 2014. We measured soil respiration, soil temperature, and soil moisture simultaneously by using the Automated Soil CO2Exchange Station (ACE, UK) for four consecutive days in July, September, November, January, February, March, May, and June, respectively, at four times each day between 8:00 and 6:00. The results showed that:(1) the two forests underwent similar seasonal changes in soil respiration, with the most change being observed during the summer, followed by spring, autumn, and winter, which yielded the least change. However, the citrus plantation presented significantly higher levels of soil respiration than the Masson pine forest across all seasons (P< 0.05). (2) Generally, nitrogen deposition suppressed soil respiration in both forests and these inhibitory effects were strengthened with increasing levels of nitrogen deposition. The only exception in which nitrogen deposition enhanced soil respiration was in the Masson pine forest during the winter, while soil moisture was very low. (3) Soil respiration showed significantly positive exponential relationships with soil temperature (P< 0.01) and significant quadratic relationship with soil moisture (P< 0.05). Both factors (soil temperature and moisture) are influenced by the levels of nitrogen deposition in subtropical forests. Thus, the results of our study on subtropical forests confirm the theory that nitrogen deposition inhibits soil respiration.
Key Words: nitrogen deposition; soil respiration; soil temperature; soil moisture; citrus plantation; Masson pine forest
近年来,全球变暖逐渐成为各界关注的热点问题,IPCC第5次评估报告(AR5)指出2003—2012年平均温度较1850—1900年上升0.78℃,并表现出加剧的趋势。CO2作为导致全球变暖最主要的气体之一,2011年其浓度达到391μL/L比1750年高40%[1- 2]。而土壤呼吸(Rs)是大气CO2的重要来源,约占整个生态系统呼吸的50%—80%,主要由土壤动物、微生物和根系等呼吸产生[3- 4],是陆地生态系统中最重要的碳循环过程[5],其产生速度受自然、生物以及各种干扰等因子影响[6]。
随着人类社会发展,氮肥施用、畜牧业快速发展及矿物燃料燃烧等活动使大气含氮化合物增多,导致大气氮沉降量加剧,且变化量有增无减[7]。氮素作为林木生长的养分限制因子,它直接或间接影响着森林土壤呼吸。目前,氮沉降引起的土壤呼吸改变逐渐成为各领域学者研究的重点,其中最常用的方法为外源氮添加模拟氮沉降试验,然而研究区域、研究时间不同所得结论各异。吴迪等[8]、刘益君等[9]、孙素琪等[10]及Sun等[11]模拟氮沉降试验表明氮沉降对土壤呼吸有抑制作用,Tu等[12]的研究结果表明氮沉降对土壤呼吸有促进作用,Hasselquist等[13]则发现氮沉降并没有明显改变林地土壤呼吸。可见,氮沉降对土壤呼吸的影响存在很大的不确定性,需要进一步的深入研究。氮沉降的土壤效应深受植被性质、土壤类型与气候差异的影响[8-13],不同的土壤水热环境也会使土壤呼吸有较大差异。北美[14]、欧洲[15]关于氮沉降试验开展较早,而国内氮沉降对森林土壤呼吸影响的试验起步较晚,且大部分集中在温带森林系统,涉及热带、亚热带森林相关研究相对较少。
本文选取我国亚热带地区广泛分布的马尾松和柑橘林为研究对象,开展人工氮添加模拟氮沉降试验。探讨不同氮沉降水平对马尾松林和柑橘林土壤呼吸及水热环境的影响,探索该区域马尾松林和柑橘林土壤呼吸的动态特征以及不同林分对氮沉降的响应,为不同林地利用途径的土壤呼吸研究提供基础数据。
1材料与方法
1.1研究区概况
研究区位于重庆市北碚区境内缙云山国家级自然保护区(106°22′E,29°45′N)内,距市中心60 km。缙云山占地面积76 km2,海拔350—951.5 m,属典型中亚热带温暖湿润季风气候。年平均温度13.6℃,最热月(8月)平均温度24.3℃,最高温43℃,最冷月(1月)平均温度3.1℃,最低温-1℃;年平均相对湿度87%,年平均降雨量1161.8 mm,年均日照1293.9 h,干湿季分明。雨季是4—9月(降雨量1243.8 mm),其中7—8月是伏旱期(蒸发量255.4 mm),降雨量小,蒸发量大。土壤以酸性黄壤及水稻土为主,伴有少量零星分布的紫色土。区内植物资源丰富,植被种类繁多,森林覆盖率达96.6%。试验地建立在海拔为350—500 m的柑橘林和马尾松林内,其中柑橘林林下无灌木层,草本层为优势群落,主要优势草本为:兔儿酸(PolygonumamphibiumL.)、糯米团(Gonostegiahirta(Bl.) Miq.)、九层塔(OcimumbasilicumL.)、加拿大飞蓬(Conyzacanadensis(L.) Cronq.)、杠板归(PolygonumperfoliatumL.)、稗草(Echinochloacrusgalli(L.) Beauv.);马尾松林主要优势灌木为:杉木(Cunninghamialanceolata(Lamb.) Hook.)、盐肤木(RhuschinensisMill.)、四川山矾(SymplocossetchuensisBrand)、异叶榕(FicusheteromorphaHemsl.)、青冈(Cyclobalanopsisglauca(Thunb) Oersted)、野桐(Mallotusjaponicus(Thunb.) Muell. Arg.)、毛桐(Mallotusbarbatus(Wall.) Muell. Arg.)、朴树(CeltissinensisPers.)、黄牛奶树(Symplocoslaurina(Retz.) Wall.)、钝叶柃(EuryaobtusifoliaH. T. Chang)、白栎(QuercusfabriHance)、慈竹(Neosinocalamusaffinis(Rendle) Keng f.)等,主要优势草本为:边缘鳞盖蕨(Microlepiamarginata(Houtt.) C. Chr.)、中华里白(Hicriopterischinensis(Ros.)Ching)、粽叶狗尾草(Setariapalmifolia(Koen.) Stapf)、菝葜(SmilaxchinaLinn.)、山莓(RubuscorchorifoliusLinn. f.)、尖叶清风藤(SabiaswinhoneiHemsl.)、香花崖豆藤(MillettiadielsianaHarms)等。
1.2实验设计
2014年5月,在海拔相近的马尾松林和柑橘林内分别选取3块面积为20 m×20 m的代表性样地,基本情况如表1所示。在每个样地沿对角线确定3个点,于每个点各设置4个1 m×1 m的小样地(间隔>1 m防相互干扰),作为氮添加处理(不考虑大气氮沉降)样方。氮添加处理设置了4个水平:对照(T0,0 gN m-2a-1)、低氮(T5,20 gN m-2a-1),中氮(T10,40 gN m-2a-1)和高氮(T15,60 gN m-2a-1)。为消除流水等对样地氮含量造成影响,施氮时保证高浓度样方在低浓度样方之下。同时将长为16 cm内径为23 cm的PVC管随机埋入小样地中并确保管顶距土壤表面3 cm,去除管内植物和凋落物。在每个季度初期选择晴天将不同浓度NH4NO3溶于1L水中,采用手提式喷雾器均匀喷洒在每个小样地内,为排除试验中溶解NH4NO3所需水分对土壤呼吸的影响,在对照样地喷洒了等体积水。
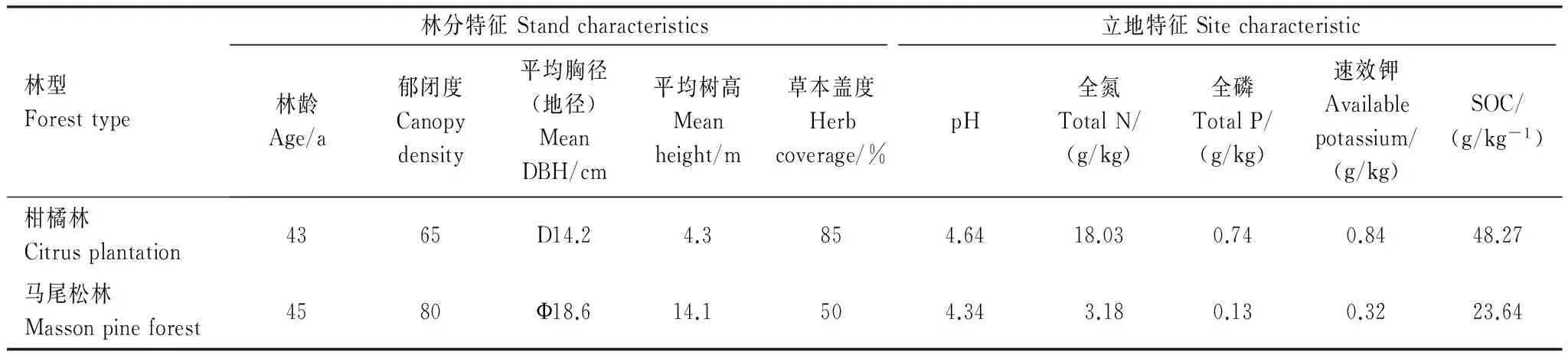
表1 重庆缙云山2种实验林(马尾松林和柑橘林)的林分与土壤特征基本信息
1.3土壤呼吸的测定
2014年7月至2015年7月,夏、秋、冬、春4个季节(夏季用7月和9月所得数据平均值表示,秋季用11月和1月所得数据平均值表示,冬季用2月和3月所得数据平均值表示,春季用5月和6月所得数据平均值表示),在施肥2d后(所施氮溶液完全渗入土壤后)选择晴朗天气,采用ACE自动土壤呼吸监测系统(Automated Soil CO2 Exchange Station, UK)于7月、9月、11月、1月、2月、3月、5月、6月对两林地土壤呼吸进行测定,各测定4d(连续),各个点每天共测定4次,测定时间为8:00—18:00(每次测定间隔2.5 h)。为了减小安放PVC管对土壤呼吸的影响,每个季度埋管与测定土壤呼吸时间间隔保持一致,并在每次测定前一天,将测定管内的地表植被自土壤表层彻底剪除,尽量不破坏土壤,以减少土壤扰动和根系损伤对测量结果的影响。呼吸圈埋好后固定1a。
在测定土壤呼吸的同时用ACE自动土壤呼吸监测系统配带的2个土壤温度热电阻探头和2个土壤水分探头SM300分别对土壤10cm深处的温度和湿度进行同步测量。
1.4数据处理与统计分析
以每个点每天测定的4个数据通过算术平均来代表该林地的当日平均土壤呼吸、土壤温度和土壤湿度。数据处理和统计分析在SPSS18.0、office Excel 2003、OriginPro9.0与MATLAB R2012B等软件中进行,采用单因素方差分析法(LSD)比较土壤呼吸在不同氮处理、不同季节间的差异水平,采用回归分析方法描述土壤呼吸与温度(T)及湿度(W)的关系。
土壤呼吸与土壤温度之间的关系采用指数模型[16]:Rs=R0eβT;Q10值通过下式计算[17- 18]:Q10=e10β;采用二次项模型[19]:Rs=a+b1W+b2W2与幂函数模型[20]:Rs=aWb研究土壤呼吸与土壤湿度的相关性;利用线性模型[21]:Rs=a+bT+cW和指数幂函数模型[22]:Rs=aebTWc研究土壤温度和土壤湿度交互作用对土壤呼吸的影响。式中Rs为土壤呼吸μmol m-2s-1,T为地温,R0是温度为0℃时的土壤呼吸,β为温度反应系数,Q10为土壤呼吸温度敏感性指数,W为土壤湿度,a,b,c为待定参数。
2结果
2.1两种林分土壤呼吸、土壤温度和土壤湿度季节特征
实验期内两林分土壤呼吸和温湿度随季节均表现出先减后增的变化趋势,且差异显著(图1)。T0为空白对照,其值反映了两森林生态系统内土壤呼吸特征。柑橘林和马尾松林土壤呼吸最大值出现在夏季,分别为(4.947±0.642)、(3.930±0.378)μmol m-2s-1,相应的地表土壤温度最大值也出现在夏季,分别为(31.10±6.287)℃、(27.77±1.575)℃;而最小值则出现在冬季,此时两林分的土壤呼吸分别为(2.868±0.695)μmol m-2s-1和(2.634±0.413)μmol m-2s-1,温度则为(14.87±1.600)℃和(13.77±0.429)℃;柑橘林(43.902±3.985)%和马尾松林(33.259±4.985)%土壤湿度最大值均出现在春季,最小值出现在秋季(20.087±1.590)%、(15.866±1.250)%(图1,图2)。
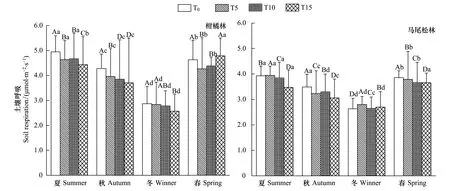
图1 重庆缙云山马尾松林和柑橘林在不同施氮处理下的土壤呼吸的季节动态变化Fig.1 Seasonal variation of soil respiration rate under the Masson pine forest and Citrus plantation with various nitrogen addition amount treatments at Mt. Jinyun, ChongqingP<0.05,大写字母表示同一季节不同处理间差异性水平,小写字母表示同一处理不同季节间差异性水平;T0:control check,对照组;T5:low nitrogen,低氮水平;T10:middle nitrogen,中氮水平;T15:high nitrogen,高氮水平
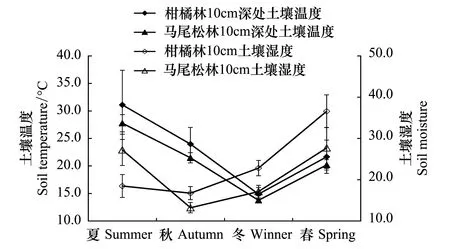
图2 重庆缙云山柑橘林和马尾松林下土壤的表层温度及湿度的季节动态变化Fig.2 Seasonal variation of soil temperature and soil moisture under the Masson pine forest and nearby Citrus plantation at Mt. Jinyun, Chongqing
2.2两种林分土壤呼吸对模拟氮沉降的响应
不同氮沉降处理的土壤呼吸具有一致的变化规律,表现为夏季最高,其次是春季和秋季,冬季最低,且各季度间差异显著(图1)。在春季,马尾松林土壤呼吸随着模拟的氮沉降量增加呈直线下降趋势,T5、T10和T15处理分别比对照T0降低了1.76%、4.86%及5.11%,但柑橘林则表现为先降低后升高的变化趋势,与对照T0相比,T5、T10和T15分别降低了7.87%、降低了5.36%、升高了3.38%;在夏季,马尾松林土壤呼吸随氮沉降量增加而表现为先升高后降低,T5、T10和T15处理分别比对照T0升高了0.44%、降低了2.11%和11.62%,但柑橘林土壤呼吸基本上呈降低趋势,T5、T10和T15处理后的土壤呼吸均比对照T0低,且分别降低6.34%、5.68%及10.30%;在秋季,较对照T0而言,两林分土壤呼吸在氮沉降处理后均降低,马尾松林T5、T10和T15处理分别降低了7.44%、5.50%、12.36%,而柑橘林T5、T10和T15处理则分别降低7.43%、10.04%、12.34%;在冬季,氮沉降处理后马尾松林T5、T10和T15处理土壤呼吸比对照T0高6.32%、0.43%和2.51%,而柑橘林T5、T10和T15处理土壤呼吸则比对照T0低0.96%、3.11%和10.52%。
2.3两种林分土壤呼吸与土壤温度的关系模型对模拟氮沉降的响应
各处理土壤呼吸与土壤温度进行回归分析(图3)结果可知,土壤呼吸与土壤温度存在极显著的指数关系(P<0.01),柑橘林T0、T5、T10和T15处理与10 cm深处土壤温度之间拟合方程的决定系数R2分别为0.613、0.358、0.358和0.377,解释了柑橘林土壤呼吸35%—62%的变异,Q10值分别为1.36、1.35、1.33、1.40;马尾松林拟合方程的决定系数R2分别为0.558、0.238、0.428和0.165,可解释马尾松林土壤呼吸16%—56%的变异,Q10值分别为1.32、1.25、1.35、1.19。

图3 重庆缙云山柑橘林和马尾松林在不同氮沉降量模拟条件下土壤10cm深处的呼吸与温度的关系Fig.3 Relationship between soil respiration and soil temperature at 10 cm depth with various nitrogen addition amount treatments under the Masson pine forest and Citrus plantation from Mt. Jinyun, Chongqing
2.4两种林分土壤呼吸与土壤湿度的关系模型对模拟氮沉降的响应
将不同处理的土壤呼吸与10 cm深处土壤湿度用二次模型和幂函数模型进行拟合,拟合度R2在0.052—0.233(表2),明显低于土壤呼吸与温度关系方程的R2值,且二次模型拟合效果优于幂函数模型。这些结果也说明,2种林分条件下的土壤湿度变化显著影响着土壤的呼吸。
表2重庆缙云山柑橘林和马尾松林下土壤表层在模拟不同氮沉降处理条件下的呼吸与湿度的关系方程
Table 2Correlation equations between soil respiration rate and soil moisture at 10cm depth across various nitrogen addition treatments under the Masson pine forest and Citrus plantation at Mt. Jinyun, Chongqing

林型Foresttype处理TreatmentRs=a+b1W+b2W2Rs=aWbab1b2PR2abPR2柑橘林T05.018-0.014-0.0010.0040.16611.110-0.3410.0020.147CitrusplantationT56.036-0.1610.0030.1060.0717.828-0.2480.0470.062T104.1160.019-0.0010.1370.0638.723-0.2930.0690.052T151.1570.156-0.0010.0140.1110.5040.6400.0000.172马尾松林T03.543-0.0460.0020.0250.1482.2020.1450.0510.078MassonpineforestT54.104-0.1350.0050.0040.2161.5090.2680.0090.136T102.8420.0010.0010.0020.2331.5700.2510.0020.186T151.9380.086-0.0010.0060.1991.4140.2700.0010.204
2.5两种林分土壤呼吸与土壤温度、土壤湿度的复合关系模型对模拟氮沉降的响应
分别利用线性模型和指数幂函数模型对土壤呼吸与土壤温度和土壤湿度的复合关系进行分析(表3)。两林分T0处理的指数幂函数模型拟合效果不如线性模型,其他处理的指数幂函数模型效果均优于线性模型。从指数幂函数模型拟合结果可知,土壤温度和湿度能够共同解释两林分土壤呼吸季节变化的56.6%—61.4%。不同氮沉降处理下马尾松和柑橘林土壤呼吸受土壤温度和土壤湿度影响程度不一样,两林分T0拟合度R2均高于氮沉降T5、T10和T15组,氮沉降处理下土壤温度和土壤湿度仅能解释土壤呼吸28.0%—47.8%的变异。
3讨论
土壤呼吸与植被类型、林下温湿度、pH、土壤质地及有机质含量等密切相关[23- 25]。不同植被的凋落物质量、数量和纤维素含量各异,这些均可影响土壤中微生物和酶等分解凋落物的强度和时间[26]。植被不同,其叶面积指数、根生物量等均有差异,叶面积大小可影响植被覆盖下土壤微型气候,Sims等[27]发现土壤CO2通量值和同时测量的叶面积指数值有显著的相关性。Althea等[28]在研究本宁堡松树林时发现土壤呼吸与距离最近的松树成负相关,与根生物量成正相关。本实验中柑橘林土壤呼吸显著高于马尾松林,原因可能是柑橘林郁闭度低、凋落物输入量大,柑橘树根细、量多且分布广,同时有机肥添加和采果、剪枝等人为活动促进了土壤养分积累与转化,加之林下良好的光、热、水等环境条件促进了土壤微生物和根系生长与代谢,增加了微生物活性与数量,加快了土壤有机质分解速度,因此产生CO2的速率加快。
表3重庆缙云山柑橘林和马尾松林林下模拟氮沉降各处理情况下土壤呼吸与10cm深度温度(T)及湿度(W)的复合关系方程
Table 3Combined relationships between soil respiration and soil temperature as well soil moisture at 10 cm depth across different nitrogen addition amount treatments under the Masson pine forest and Citrus plantation at Mt. Jinyun, Chongqing
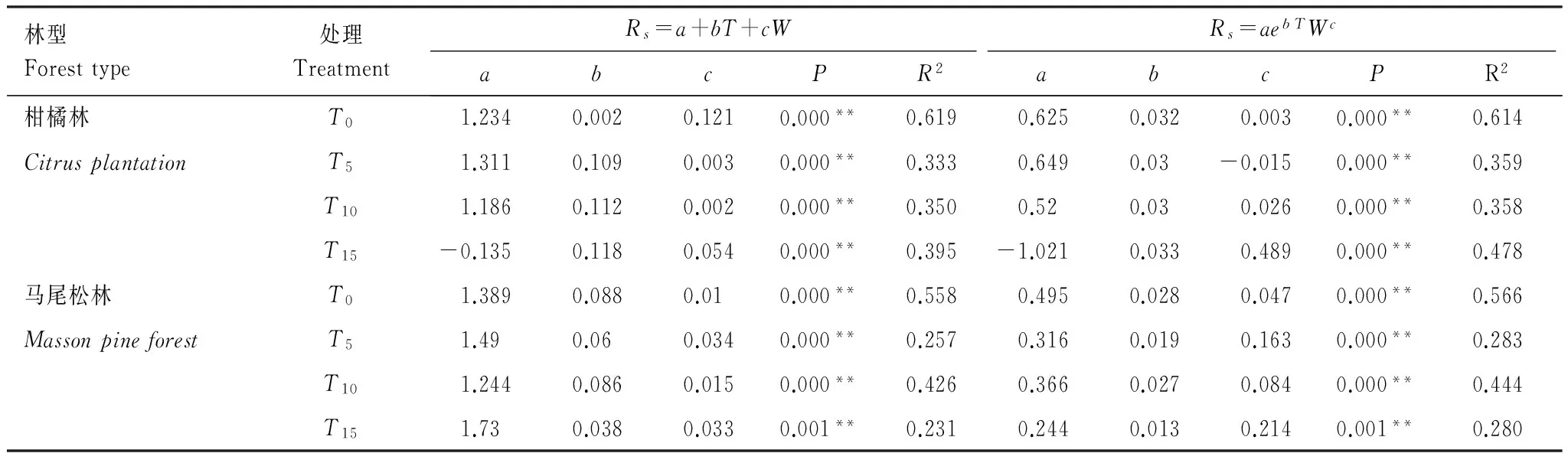
林型Foresttype处理TreatmentRs=a+bT+cWRs=aebTWcabcPR2abcPR2柑橘林T01.2340.0020.1210.000**0.6190.6250.0320.0030.000**0.614CitrusplantationT51.3110.1090.0030.000**0.3330.6490.03-0.0150.000**0.359T101.1860.1120.0020.000**0.3500.520.030.0260.000**0.358T15-0.1350.1180.0540.000**0.395-1.0210.0330.4890.000**0.478马尾松林T01.3890.0880.010.000**0.5580.4950.0280.0470.000**0.566MassonpineforestT51.490.060.0340.000**0.2570.3160.0190.1630.000**0.283T101.2440.0860.0150.000**0.4260.3660.0270.0840.000**0.444T151.730.0380.0330.001**0.2310.2440.0130.2140.001**0.280
**表示在0.01显著水平下的差异性;*表示在0.05显著水平下的差异性
土壤呼吸有明显的季节性变化,丁访军等[29]在研究喀斯特地区的灌木林、阔叶混交林和针阔混交林土壤呼吸时发现:土壤呼吸呈夏季>秋季>春季>冬季的变化规律。这与刘春霞等[30]研究结论一致。而本实验结果为夏季>春季>秋季>冬季,春季>秋季的结论与丁访军、刘春霞等研究不一致,原因可能是研究对象、处理空间和测定时间的异质性导致的。实验区夏季土壤呼吸最大,夏季光照充足、温度较高、降雨量也最丰富,良好环境条件有利于植物生长发育,促进根系呼吸,同时加快凋落物分解,增加微生物种群数量及其代谢活性并使土壤中聚集较高的土壤酶活性,进而使土壤微生物呼吸、土壤动物呼吸及土壤中氧化还原反应产生CO2的速度加快。
氮沉降对森林土壤呼吸的影响主要有3种:促进、抑制和不显著。总的来说,本实验除冬季氮沉降促进马尾松林土壤呼吸外,其余均表明氮沉降抑制土壤呼吸。这与国内外很多研究结果一致。比如,吴迪等[8]在研究长江中下游欧美杨(Populus dettoides)人工林时发现低氮、中氮和高氮处理使CO2排放量分别减少12.35%、14.45%和15.73%。刘益君等[9]发现,氮沉降显著抑制了湿地松林分的土壤呼吸且抑制作用第1年显著大于第2年,施氮处理后第1年低、中、高处理土壤呼吸分别比对照降低了31.3%、27.4%和24.5%,第2年则分别降低24.0%、16.5%和25.3%。孙素琪等[10]于2012年研究缙云山常绿阔叶林对模拟氮沉降的响应时发现低、中、高氮处理分别使土壤呼吸降低了20.88%,34.80%和42.23%。Sun等[11]在研究河北塞罕坝国家森林公园落叶松林土壤呼吸时发现低氮处理使土壤呼吸降低10.0%,高氮则降低12.5%,与本文结果基本一致但与冬季马尾松林处理后所得结果相异。也有研究表明氮沉降对土壤呼吸有促进作用,Tu等[12]在研究竹林生态系统时发现氮沉降显著增加了土壤呼吸。Hasselquist等[13]研究结果则表明氮沉降对土壤呼吸无影响。这可能跟植被、土壤性质、施氮处理方式、处理与测定时间间隔长度以及气候条件等不同有关。
氮沉降使土壤呼吸降低的原因可能是:①氮沉降增加了土壤铵态氮和硝态氮含量,高浓度氨氮含量降低微生物活性和数量,产生抑制物质或影响土壤酶系统,从而阻碍凋落物分解和养分释放,进而减慢土壤碳释放速度。②氮沉降加强土壤硝化作用,使森林土壤pH值降低,加速土壤酸化[31],降低植物根系生长和根生物量,致使根际分泌物减少,进而抑制植物根系呼吸[10]。③在氮素丰富的地区,当森林土壤氮含量达到饱和状态后,过量的氮沉降就会对土壤呼吸产生抑制的作用[32- 33]。
缙云山两林分土壤表层呼吸受土壤温度和湿度共同影响。图4是基于指数幂函数模型的土壤呼吸对温湿度协同变化的响应趋势面。该响应趋势面表明:第一,土壤呼吸随温度升高而加强;第二,低温下,土壤呼吸受土壤湿度影响较小,高温时,土壤呼吸则受温度和湿度协同控制;第三,土壤湿度对马尾松林土壤呼吸影响较明显,但对柑橘林土壤呼吸影响较小。Zeng等[34]在研究太行山洋槐林、黄荆林及铁杆蒿地时发现土壤呼吸主要受土壤温度驱动而与土壤湿度仅有微弱的关系。Chen等[35]研究北京奥林匹克公园时发现土壤呼吸年周期主要受温度控制,而夏季土壤呼吸主要受湿度控制,这均与本研究结论一致。说明土壤呼吸受土壤温度影响更大,而多数情况下土壤呼吸受土壤温度及湿度共同控制。

图4 柑橘林和马尾松林土壤呼吸的温湿度响应趋势面Fig.4 Response surface of soil respiration to soil temperature and soil moisture in Masson pine forest and Citrus plantation
4结论
(1)两林分土壤呼吸均具有明显的季节变化规律,其中夏季土壤呼吸最高,其次是春季和秋季,最低为冬季,且柑橘林土壤呼吸明显高于马尾松林,不同季度观测的土壤呼吸平均值之间差异显著(P<0.05)。
(2)总体而言,氮沉降抑制了两林分土壤表层呼吸,抑制程度随氮沉降增加而升高,只在土壤湿度低的冬季,氮沉降对马尾松林土壤呼吸有促进作用。
(3)土壤呼吸与土壤温度存在极显著的指数关系,与土壤呼吸有显著的二次模型拟合关系,但均受氮沉降量处理的影响。
[1]IPCC.ClimateChange2013:ThePhysicalScienceBasis:WorkingGroupIContributiontotheIPCCFifthAssessmentReport(AR5),FinalDraftUnderlyingScientific-TechnicalAssessment. [2015-06- 28].http://www.climatechange2013.org/images/report/WG1AR5_SPM_FINAL.pdf
[2]唐秀萍. 碳汇 拓展林业外部空间. 中国林业, 2005, (22): 11- 17.
[3]SchlesingerWH,AndrewsJA.Soilrespirationandtheglobalcarboncycle.Biogeochemistry, 2000, 48(1): 7- 20.
[4]SehimelDS.Terrestrialecosystemsandthecarboncycle.GlobalChangeBiology, 1995, 1(1): 77- 91.
[5]ChenST,ZouJW,HuZH,ChenHS,LuYY.Globalannualsoilrespirationinrelationtoclimate,soilpropertiesandvegetationcharacteristics:Summaryofavailabledata.AgriculturalandForestMeteorology, 2014, 198- 199: 335- 346.
[6]魏书精, 罗碧珍, 孙龙, 魏书威, 刘芳芳, 胡海清. 森林生态系统土壤呼吸时空异质性及影响因子研究进展. 生态环境学报, 2013, 22(4): 689- 704.
[7]LuCQ,TianHQ,LiuML,RenW,XuXF,ChenGS,ZhangC.EffectofnitrogendepositiononChina′sterrestrialcarbonuptakeinthecontextofmultifactorenvironmentalchanges.EcologicalApplications, 2012, 22(1): 53- 75.
[8]吴迪, 张蕊, 高升华, 付晓, 邓红兵, 邵国凡, 张旭东. 模拟氮沉降对长江中下游滩地杨树林土壤呼吸各组分的影响. 生态学报, 2015, 35(3): 717- 724.
[9]闫文德, 徐旺明, 郑威, 王光军, 张徐源, 梁小翠, 高超. 施氮对湿地松(Pinus elliottii)林土壤呼吸及其相关因子的消减效应. 生态学报, 2016, 36(2),doi:10.5846/stxb201311192767.
[10]孙素琪, 王玉杰, 王云琦, 张会兰, 李云霞, 于雷, 胡波, 刘婕. 缙云山常绿阔叶林土壤呼吸对模拟氮沉降的响应. 林业科学, 2014, 50(1): 1- 8.
[11]SunZZ,LiuLL,MaYC,YinGD,ZhaoC,ZhangY,PiaoSL.Theeffectofnitrogenadditiononsoilrespirationfromanitrogen-limitedforestsoil.AgriculturalandForestMeteorology, 2014, 197: 103- 110.
[12]TuLH,HuTX,ZhangJ,LiXW,HuHL,LiuL,XiaoYL.Nitrogenadditionstimulatesdifferentcomponentsofsoilrespirationinasubtropicalbambooecosystem.SoilBiologyandBiochemistry, 2013, 58: 255- 264.
[13]HasselquistNJ,MetcalfeDB,HögbergP.ContrastingeffectsoflowandhighnitrogenadditionsonsoilCO2fluxcomponentsandectomycorrhizalfungalsporocarpproductioninaborealforest.GlobalChangeBiology, 2012, 18(12): 3596- 3605.
[14]GundersenP,EmmettBA,KjønaasOJ,KoopmansCJ,TietemaA.Impactofnitrogendepositiononnitrogencyclinginforests:asynthesisofNITREXdata.ForestEcologyandManagement, 1998, 101(1/3): 37- 55.
[15]FennME,PothMA,AberJD,BaronJS,BormannBT,JohnsonDW,LemlyAD,McNultySG,RyanDF,StottlemyerR.NitrogenexcessinNorthAmericanecosystems:predisposingfactors,ecosystemresponses,andmanagementstrategies.EcologicalApplications, 1998, 8(3): 706- 733.
[16]FangC,MoncriefJB.ThedependenceofsoilC02effluxontemperature.SoilBiologyandBiochemistry, 2001, 33(2): 155- 165.
[17]BowdenRD,NadelhofferKJ,BooneRD,MelilloJM,GarrisonJB.Contributionsofabovegroundlitter,belowgroundlitter,androotrespirationtototalsoilrespirationinatemperatemixedhardwoodforest.CanadianJournalofForestResearch, 1993, 23(7): 1402- 1407.
[18]ReyA,PegoraroE,TedeschiV,DeParriI,JarvisPG,ValentiniR.AnnualvariationinsoilrespirationanditscomponentsinacoppiceoakforestincentralItaly.GlobalChangeBiology, 2002, 8(9): 851- 866.
[19]SottaED,MeirP,MalhiY,NobreAD,HodnettM,GraceJ.SoilCO2effluxinatropicalforestinthecentralAmazon.GlobalChangeBiology, 2004, 10(5): 601- 617.
[20]姜艳, 王兵, 汪玉如, 杨清培. 亚热带林分土壤呼吸及其与土壤温湿度关系的模型模拟. 应用生态学报, 2010, 21(7): 1641- 1648.
[21]WildungRE,GarlandTR,BuschbomRL.Theinterdependenteffectsofsoiltemperatureandwatercontentonsoilrespirationrateandplantrootdecompositioninaridgrasslandsoils.SoilBiologyandBiochemistry, 1975, 7(6): 373- 378.
[22]李化山, 汪金松, 刘星, 蒋思思, 张春雨, 赵秀海. 模拟氮沉降对太岳山油松林土壤呼吸的影响及其持续效应. 环境科学学报, 2014, 34(1): 238- 249.
[23]SongXZ,PengCH,ZhaoZY,ZhangZT,GuoBH,WangWF,JiangH,ZhuQ.QuantificationofsoilrespirationinforestecosystemsacrossChina.AtmosphericEnvironment, 2014, 94: 546- 551.
[24]ShabagaJA,BasilikoN,CaspersenJP,JonesTA.Seasonalcontrolsonpatternsofsoilrespirationandtemperaturesensitivityinanorthernmixeddeciduousforestfollowingpartial-harvesting.ForestEcologyandManagement, 2015, 348: 208- 219.
[25]李凯, 江洪, 由美娜, 曾波. 模拟氮沉降对石栎和苦槠幼苗土壤呼吸的影响. 生态学报, 2011, 31(1): 82- 89.
[26]张东秋, 石培礼, 张宪洲. 土壤呼吸主要影响因素的研究进展. 地球科学进展, 2005, 20(7): 778- 785.
[27]SimsPL,BradfordJA.Carbondioxidefluxesinasouthernplainsprairie.AgriculturalandForestMeteorology, 2001, 109(2): 117- 134.
[28]ArchMillerAA,SamuelsonLJ.Intra-annualvariationofsoilrespirationacrossfourheterogeneouslongleafpineforestsinthesoutheasternUnitedStates.ForestEcologyandManagement, 2015,doi:10.1016/j.foreco.2015.05.016.
[29]丁访军, 高艳平, 吴鹏, 崔迎春, 胡蕖. 喀斯特地区3种林型土壤呼吸及其影响因子. 水土保持学报, 2010, 24(3): 217- 221, 237- 237.
[30]刘春霞, 王玉杰, 王云琦, 冯华敏, 胡波. 重庆缙云山3种林型土壤呼吸及其影响因子. 土壤通报, 2013, 44(3): 587- 593.
[31]肖辉林. 大气氮沉降对森林土壤酸化的影响. 林业科学, 2001, 37(4): 111- 116.
[32]FranklinO,HögbergP,EkbladA, ÅgrenGI.PineforestfloorcarbonaccumulationinresponsetoNandPKadditions:bomb14Cmodellingandrespirationstudies.Ecosystems, 2003, 6(7): 644- 658.
[33]MaierCA,KressLW.SoilCO2evolutionandrootrespirationin11year-oldloblollypine(Pinus taedaL.)plantationsasaffectedbymoistureandnutrientavailability.CanadianJournalofForestResearch, 2000, 30(3): 347- 359.
[34]ZengXH,ZhangWJ,ShenHT,CaoJS,ZhaoX.SoilrespirationresponseindifferentvegetationtypesatMountTaihang,China.Catena, 2014, 116: 78- 85.
[35]ChenWJ,JiaX,ZhaTS,WuB,ZhangYQ,LiCY,WangXP,HeGM,YuHQ,ChenGP.SoilrespirationinamixedurbanforestinChinainrelationtosoiltemperatureandwatercontent.EuropeanJournalofSoilBiology, 2013, 54: 63- 68.
基金项目:国家自然科学基金项目(41401337)
收稿日期:2015- 06- 30; 2015- 06- 30;
修订日期:2015- 11- 27 2015- 10- 29
*通讯作者
Corresponding author.E-mail: suyzh@lzb.ac.cn .E-mail: hebinghui@swu.edu.cn
DOI:10.5846/stxb201506301362 10.5846/stxb201506301367
Soil stoichiometry characteristics ofHaloxylonammodendronwith different plantation age in the desert-oasis ecotone, north China
ZHANG Ke1,2, SU Yongzhong1,*, WANG Ting1,2, LIU Tingna1
1KeyLaboratoryofEco-HydrologyinInlandRiverBasin,LinzeInlandRiverBasinResearchStation,ColdandAridRegionsEnvironmentalandEngineeringResearchInstitute,ChineseAcademyofSciences,Lanzhou730000,China2UniversityofChineseAcademyofSciences,Beijing100049,China
Abstract:Vegetation construction is one of the most important measures of ecological restoration in arid area and also one of the most effective methods of desertification control. Haloxylon ammodendron, one important sand-fixation plant in desert region of China with remarkable tolerate to drought, hot environmental stress, and can grow rapidly after planting on sand lands, was widely used for restoring vegetation on the peripheries of the oasis to fix the sand dunes in Linze Station since 1975. To clarify the effect of H. ammodendron on soil nutrient stoichiometric characteristics and explore the indicative significance of nutrient stoichiometric ratios on the recovery of soil, this study conducted the research on the artificial forest of H. ammodendron in an desert-oasis ecotone by analyzing soil organic carbon (SOC), total nitrogen (TN), total phosphorus (TP), Available P and CaCO3 in an age sequence of 0-, 2- , 5- , 9- , 13- , 16- , 31- , and 39- year-old H. ammodendron plantations. Results showed that (1) distribution of SOC, TN and TP presented significant layered phenomenon along with the increase of plantation age, even though this feature in TP was relatively backward. In different soil depth, SOC and TN increased significantly with the increase of plantation age, but TP has no significant variation. (2) C∶P and N∶P also displayed obvious stratification trait and C∶P and N∶P in different layer also increased with the increase of plantation age. However, C∶N presented at a relatively constant variation. (3) The lower content of Available P expressed significant stratification feature after 2-year-old plantation and Available P and CaCO3 in 0—5 cm increased with the increase of plantation age. Nevertheless, CaCO3∶Available P also has no remarkable changes. (4) With the increase of plantation age, soil erodible fraction was highly significant decreased and has remarkably negative correlated with soil nutrients. These results exhibited significant increase of SOC and TN with the increase of establishment time. C∶N exhibited a stable trend, compared with C∶P and N∶P, indicating the formation of nitrogen need a relative fixation ratio of carbon and also stating C∶N that less susceptible to the effects of climate factors is quite stable. The small variation of TP mainly derived from the weathering of rock was strongly restricted by time. Moreover, the lower content of Available P and the narrow change of CaCO3∶ Available P manifested that the growth of H. ammodendron may limited by soil P and the limited Available P was more fixed by CaCO3. In addition, the negative relationship between soil nutrients and soil erodible fraction further showed that the establishment of H. ammodendron has improved soil texture and enhance soil nutrients contents. Our findings aimed at soil environment has enriched the study on H. ammodendron. However, further study is needed to clarify the response of soil nutrient stoichiometric characteristics to H. ammodendron and herbaceous, so as to provide more knowledge for the restoration effect of H. ammodendron on arid environmental and maintain sustainable development under the limited water condition.
Key Words:Haloxylon ammodendron; plantation age; soil stoichiometry characteristics; desert-oasis ecotone
Seasonal responses of soil respiration to simulated nitrogen deposition in a citrus plantation and masson pine forest in Mt. Jinyun, Chongqing, China
ZENG Qingping1, HE Binghui1,*, MAO Qiaozhi1, QIN Huajun2, LI Yuan3, HUANG Qi4
1KeyLaboratoryofThreeGorgesRegionEco-Environments,MinistryofEducation,CollegeofResourcesandEnvironment,SouthwestUniversity,Chongqing400715,China2GuizhouInstituteofBiology,Guiyang550009,China3ForestryAdministrationofFuling,Chongqing408000,China4HydroChinaGuiyangEngineeringCorporationLimited,Guiyang550009,China
张珂,苏永中,王婷,刘婷娜.荒漠绿洲区不同种植年限人工梭梭林土壤化学计量特征.生态学报,2016,36(11):3235- 3243.
Zhang K, Su Y Z, Wang T, Liu T N.Soil stoichiometry characteristics ofHaloxylonammodendronwith different plantation age in the desert-oasis ecotone, north China.Acta Ecologica Sinica,2016,36(11):3235- 3243.
