羌塘高寒草地物种多样性与生态系统多功能关系格局
2016-08-09熊定鹏赵广帅武建双石培礼张宪洲
熊定鹏,赵广帅,武建双,石培礼,*,张宪洲
1 中国科学院地理科学与资源研究所生态系统网络观测与模拟重点实验室, 北京 100101 2 中国科学院大学, 北京 100049
羌塘高寒草地物种多样性与生态系统多功能关系格局
熊定鹏1,2,赵广帅1,2,武建双1,石培礼1,*,张宪洲1
1 中国科学院地理科学与资源研究所生态系统网络观测与模拟重点实验室, 北京100101 2 中国科学院大学, 北京100049
摘要:传统的生物多样性-生态系统功能研究大多侧重于单一生态系统功能与物种多样性的关系,忽略了生态系统的重要价值在于其能够同时提供多种功能或服务,即生态系统的多功能性。基于藏北羌塘高寒草地样带调查数据,选取植被地上生物量、地下生物量、土壤全氮、硝态氮及铵态氮含量、土壤全磷含量、土壤有机碳储量等7个与植物生长、养分循环、土壤有机碳蓄积相关的参数来表征生态系统多功能性。采用上述参数转换为Z分数后的平均值计算多功能性指数(M)。分析了不同生物多样性指数与生态系统多功能指数的关系以及年降水量和年均温度对物种多样性和生态系统多功能性指数的影响。结果表明,物种丰富度指数与生态系统多功能性之间呈极显著的正相关关系,Shannon-wiener和Simpson物种多样性指数也与多功能性指数间呈显著的正相关,但多功能性指数与Pielou均匀度指数没有表现出明显的相关关系。物种丰富度与表征植物生长、养分循环以及土壤有机碳蓄积的生态系统功能指数间也均呈极显著的正相关关系。降水格局显著影响羌塘高原物种丰富度和生态系统多功能指数,二者均随年降雨量的增加而显著增加,但物种多样性指数并未与年降水量呈现显著相关关系。研究强调了群落物种丰富度即群落物种数量对维持生态系统多功能性的重要意义,这意味着由于人类活动导致的物种丧失可能会给藏北高寒草地生态系统多功能和生态服务带来更为严重的后果。就退化草地恢复或草地可持续管理而言,在藏北羌塘地区,本地植物种的物种丰富度恢复和维持应作为重要目标之一。
关键词:羌塘高原;高寒草地;物种多样性;物种丰富度;生态系统多功能性
在全球范围内,由于人类活动导致的物种灭绝和生态系统结构简化的趋势不断加剧,生态系统中物种丧失可能带来的潜在生态后果备受关注。近20年以来,生物多样性与生态系统过程(功能)的关系成为生态学领域内的核心科学问题。大量相关实验研究在全球范围内广泛开展,多数实验结果表明生物多样性与单一的生态系统功能(例如:生产力,氮素累积)之间存在因果关系[1- 3],多样性导致更高的群落生产力、更高的系统稳定性和更高的抗入侵能力[1,4-5]。对生物多样性-生态系统功能关系实验结果的整合分析也表明,物种多样性与生态系统功能之间呈正相关关系,随物种多样性的增加这种关系通常会趋于饱和,但相关的机理性解释仍存在较大争议[6-8]。然而,这些生物多样性-生态系统功能关系研究仅考虑了单一生态系统功能与物种多样性的关系,却忽略了生态系统最为重要价值在于其能够同时维系或提供诸如净初级生产力、碳氮循环、养分维持等多种不同的功能或服务[9-10],即生态系统的多功能性(multifunctionality)[11]。
近年来,一些学者认为只有将各类的生态系统系统功能作为整体考虑,才能更利于全面地理解生态系统功能与生物多样性的关系。此外,从生态系统多功能性角度出发,也为生态系统管理提供了一个全新的、综合性的视角。因为生态系统功能间存在着权衡(trade-off)关系,例如,植物生产力与抗胁迫能力间通常表现为负相关,即不太可能同时最大化此两种生态系统功能[12],拥有较高的某种生态功能的物种,其性状通常限制了其他功能水平。因此仅考虑某一生态系统功能的管理策略可能会削弱生态系统维持或提供其他功能的能力。在此背景下,生物多样性-生态系统多功能性关系研究逐渐成为一个新的热点议题[9,12]。例如,Hector[9]基于欧洲草地BIODEPTH实验数据,选择了7种生态系统过程参数包括地上、地下生产力、地上植被碳库、土壤无机氮含量、光合有效辐射传输比率以及木质素和纤维素降解速率,研究了物种多样性与生态系统多功能性的关系。Zavaleta[12]等根据美国Cedar Creek长期实验研究数据,综合考虑了地上、地下生产力、土壤无机氮含量、群落抗入侵能力、昆虫物种多样性、土壤碳库和植物氮素含量8种生态系统功能,分析了不同时间尺度上物种多样性与生态系统多功能性的关系。Maestre[10]等基于全球干旱地区的实验结果研究生态系统多功能性与物种丰富度的关系,其选取了与生态系统碳、氮、磷循环相关的14种生态系统功能来表征生态系统多功能。以上生物多样性-生态系统多功能性研究都认为维持或提供生态系统多功能性需要更多的物种数或功能群组合[9-10,12-13],可能原因是不同物种(或功能型)对不同的生态系统功能(过程)都具有重要影响。然而,目前国内对生态系统的多功能研究还很少,只有对农林生态系统多功能的概念框架进行了介绍[14-16],而对多功能性与生物多样性的关系研究尤其缺乏。毫无疑问,这会局限了人们对生态系统多功能性的认识,不利于生态系统管理和生态服务的发挥。
藏北高原地处青藏高原西北腹地,是西藏重要畜牧业生产基地,也是生态环境极为脆弱的典型区域[17]。草地是藏北地区最主要的生态系统类型,各类天然草地面积达4.8×105km2, 占藏北高原土地面积的81%[18]。高寒草地生态系统为藏北地区牧民提供了净初级物质生产、碳蓄积与碳汇、调节气候、涵养水源、水土保持、防风固沙和维持生物多样性等生态服务功能,是区域牧业经济发展的基础,也是青藏高原生态安全屏障的重要组成部分[19-21]。该区域以往生物多样性-生态系统功能关系研究多集中于样点尺度,且仅针对单一的生态系统功能,基于样带尺度的研究也仅限于分析地上生产力与生物多样性的关系格局[22]。尚缺乏藏北天然草地生态系统多功能性和生物多样性关系的定量研究。据此,本研究采用样带分析方法,基于样地调查数据,综合7种与植被生产力、养分循环、土壤有机碳蓄积相关的生态系统参数,分析了藏北高寒草地物种多样性和功能多样性与生态系统多功能性之间的关系。研究结果将加深人们对高寒草地生态系统多功能性与生物多样性关系的认识,为草地生物多样性保护以及可持续利用管理提供有益科学参考。
1材料与方法
1.1研究区概况
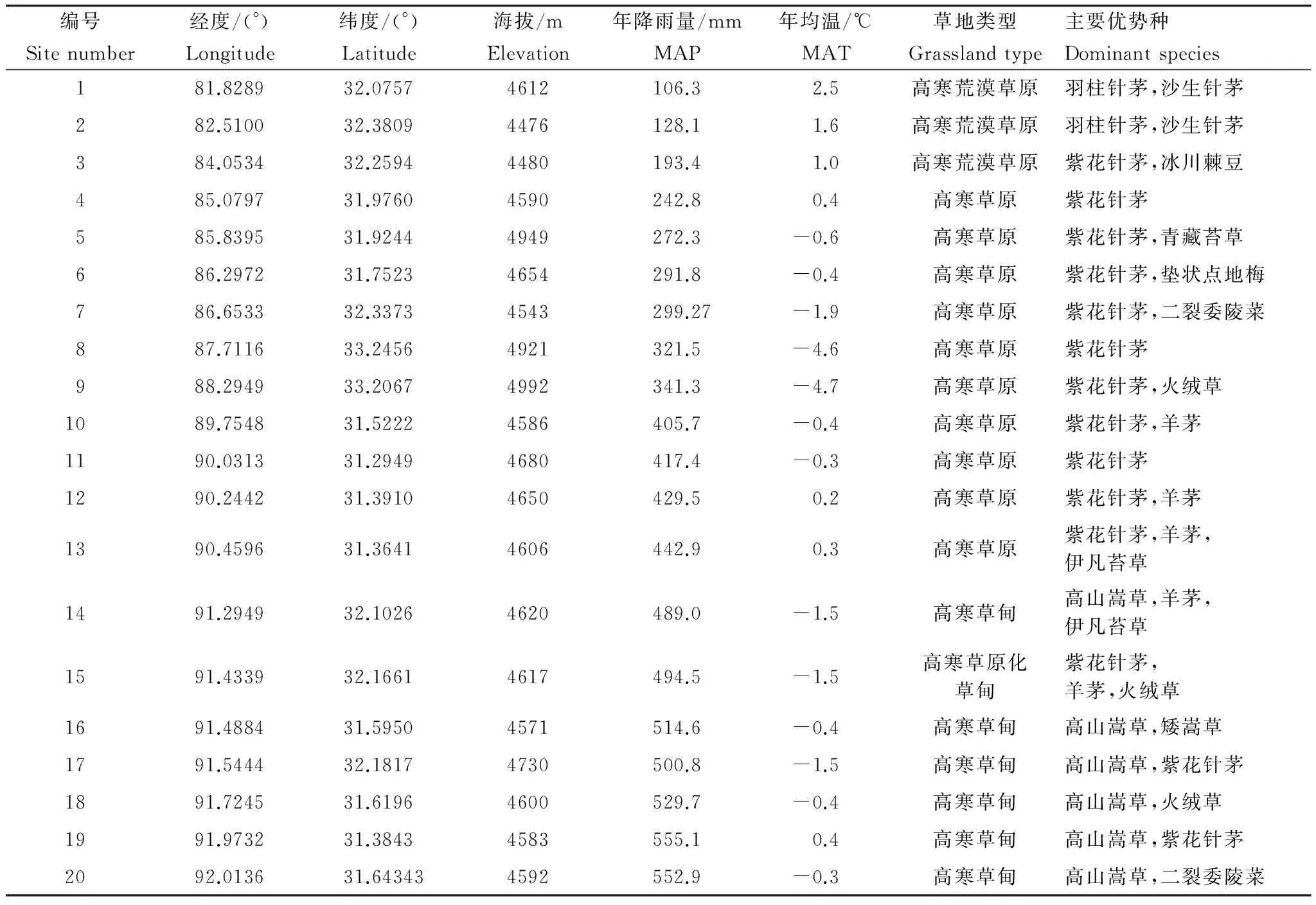
藏北高原,又称“羌塘”,意为北方高平地,位于西藏自治区北部。整个羌塘地区,东迄91°E左右的内外流水系之分水岭,西至国界并与克什米尔地区相邻接,南达冈底斯-念青唐古拉山脉,北到昆仑山脉,面积约为5.95×105km2[18]。研究区属亚寒带气候,气候寒冷干燥,海拔在4600—5100m之间,大部分地区年平均气温低于0℃, 最暖月均温不及14℃, 最冷月均温-10℃以下。受西南季风运动路径及地形因素影响,降水自东向西呈递减趋势,其中65%—85%的年降水量集中在植物生长旺盛的6—8月。研究区内部主要包括高寒草甸,高寒草原,高寒荒漠草原等地带性高寒草地生态系统类型[23]。土壤类型主要包括栗钙土、寒冻钙土、雏漠土及寒漠土等。
1.2野外取样和室内分析
2011年7月下旬—8月初(植物生长旺盛期),在藏北羌塘高原东西样带上选取了20个围栏样地作为调查样点(表1),研究样地均为西藏自治区退牧还草工程划定的禁牧草地,于2004或2005年起实施全年禁牧管理。在每个样地选取一块地势相对平坦,植物生长均匀的区域(100m×100m)进行取样调查,在该区域对角线上设置一条 100m 样线,沿样线设置5个0.5m×0.5m的样方,相当于5次重复。在各样方中,分别记录样方内植物种类、分物种盖度、平均高度和群落总盖度,采用齐地刈割法收获分种地上生物量;同时在每个样点随机抛0.1m2样圈30次,记录所出现的物种和次数,用以计算样地的物种丰富度和各物种的相对频度。每个样方,地上部分刈割后,用根钻(直径=7cm)取4钻(0—20cm深度)带根系的土样,混合后装入布袋中,样品带回住处后立即冲洗漂净并将根系分离;随后用直径4cm的土钻获取5钻(0—20cm深度)土壤样品,土壤样品混合后一部分装入塑封袋中放入冰箱冷冻,一部分装入布袋待测。此外,在样线旁挖掘一个容重坑,用环刀法获取土壤容重样品,用以测定土壤容重及砾石比。
1.3数据处理1.3.1物种重要值
重要值(IV)是群落物种的重要定量指标, 本研究采用草地调查样方内物种相对盖度(Cr),相对高度(Hr)和样圈法统计的物种相对频度(Fr)计算重要值[23],公式如下:
IV=(Cr+Hr+Fr)/3 (1)
1.3.2物种多样性
本研究采用物种丰富度指数(Species richness,S),Shannon-wiener多样性指数(H′),Simpson优势度指数(P),Pielou均匀度指数(E)来表征物种多样性[26]。
(2)
(3)
(4)
式中,S为群落物种数目,Pi为第i种植物的相对重要值。
1.3.3生态系统多功能性指数
选择与植物生长(地上生物量、地下生物量)、养分循环(土壤全氮含量、土壤氨态氮含量、土壤硝态氮含量和土壤全磷含量)和土壤有机碳蓄积(土壤有机碳密度)相关的7种重要生态系统参数来表征生态系统多功能性(M)。为量化生态系统多功能性,首先计算20个调查样地7种生态系统参数的Z分数,单样本Kolmogorov-Smirnov检验结果表明各参数均服从正态分布(P> 0.05)(表2),因此直接使用原始数据进行计算。Z分数计算公式如下:
Zij=(xij-μj)/σj
(5)
式中,Zij为样地i第j种生态系统参数的Z分数,i的范围在1—7之间,j的范围在1—20之间;xij为样地i第j种生态系统参数的数值;μj为第j种生态系统参数在20个样地间的平均值;σj为第j种生态系统参数在20个样地间的标准差。
对每一样地7种不同生态系统参数的Z分数求平均值,即为各样地的多功能多样性指数(M)。计算公式如下:
(6)
式中,Mi为样地i的生态系统多功能性指数。该方法的优点是,其计算得到多功能性指数具有较好的统计学特性,且不会改变原始数据的变异程度,计算简便,能够直观的表征植物群落同时提供多种生态系统功能或服务的能力[10]。
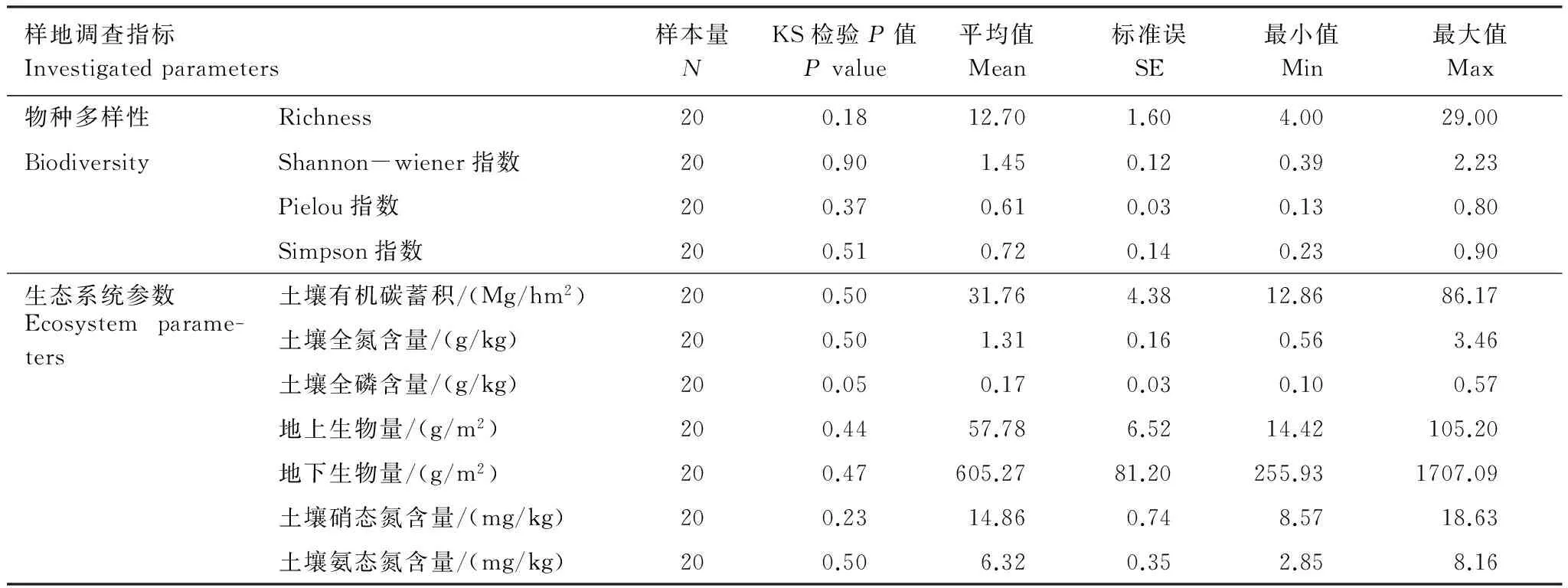
表2 羌塘样带各样地调查指标描述性统计
1.4数据分析
通过一元线性回归方法分析物种多样性和功能多样性指数与生态系统多功能性指数间的关系,此外,将7个变量划分为与植物生长、养分循环、土壤有机碳蓄积相关的3种重要生态系统功能,并分别与不同生物多样性指数进行线性回归。同时分析了年降水量、年均温度与物种多样性和生态系统功能多样性指数间的线性回归关系。以上分析采用最小二乘法拟合回归方程。所有数据分析在SPSS 20.0中进行。
2结果与分析
线性回归分析结果表明(表3,图1),藏北高寒草地生态系统多功能性指数随物种丰富度的增加而呈极为显著的增加趋势,物种丰富度解释了约55%的草地生态系统多功能性指数的变化。Shannon-wiener多样性指数和Simpson优势度指数也与藏北高寒草地生态系统多功能性指数间呈显著的正相关关系(P<0.05),Pielou均匀度指数与多功能性指数间拟合结果不显著(表3,图1)。
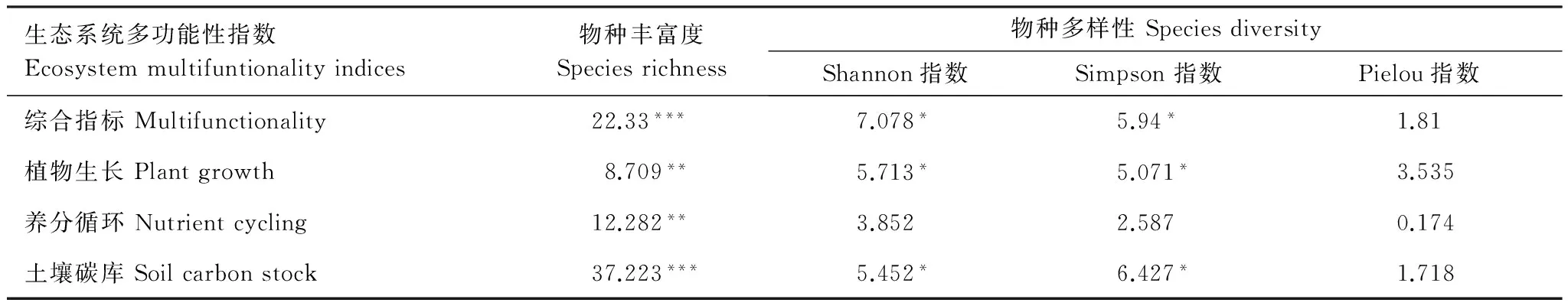
表3 生物多样性与生态系统多功能性线性回归的F值和显著性
*P<0.05, **P<0.01, ***P<0.001
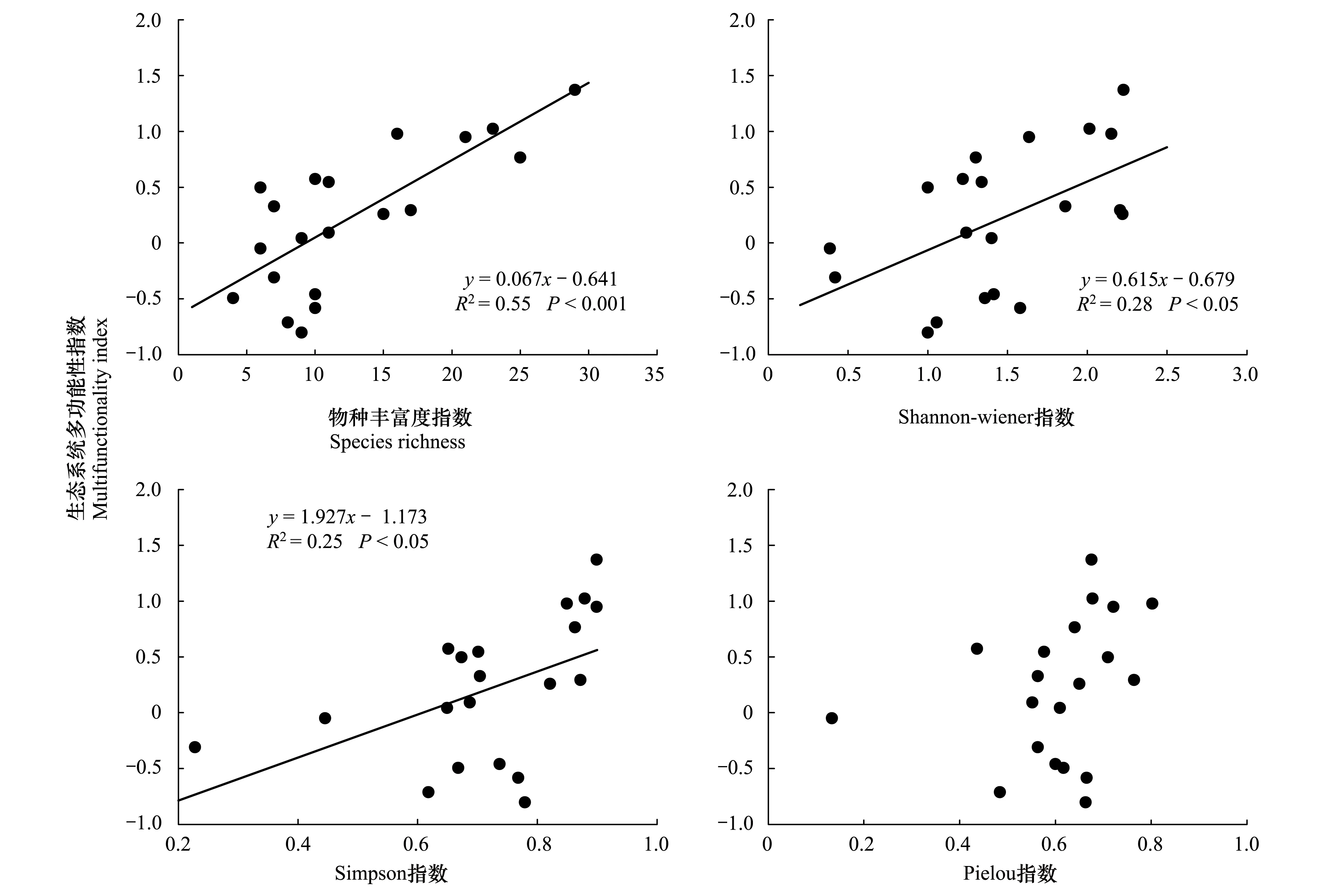
图1 高寒草地物种多样性与生态系统多功能性的关系Fig.1 Relationship between species diversity and ecosystem multifunctionality in alpine grasslands
物种丰富度和表征植物生长、养分循环以及土壤有机碳蓄积3种重要的生态系统功能指数间均呈极显著的正相关关系,且线性拟合的P值均小于0.01,在统计上达到了99%的置信度水平(表3,图2)。与植物生长和土壤有机碳蓄积相关的生态系统功能指数呈现随Shannon-wiener指数和Simpson指数增加而显著增加的趋势,但与养分循环相关的生态系统功能指数与两种多样性指数间无明显相关关系(表3,图2)。Pielou均匀度指数与3种生态系统功能指数间均未表现出明显的相关关系。两种功能多样性指数与3种生态系统功能指数间的线性拟合结果均不显著(表3)。
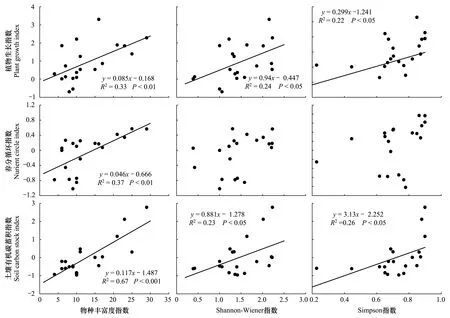
图2 高寒草地物种多样性与不同生态系统功能的关系Fig.2 Relationship between species diversity and different ecosystem functions in the alpine grasslands
羌塘高原草地群落的物种丰富度随年降雨量的增加显著增加(R2=0.27,P<0.05),但年降雨量与Shannon-wienner指数、Simpson指数和Pielou指数间没有呈现出明显的相关关系。年降水量显著的影响生态系统多功能性及其相关组分,其与多功能性综合指数和表征植物生长、土壤养分循环相关的功能性指数间均呈极其显著的正相关关系(P<0.001),与土壤有机碳蓄积指数间呈显著的正相关关系(图3)。而物种多样性指数和多功能性指数与温度之间均没有相关关系。可见,藏北羌塘高原降水梯度上多功能性及其组分指数随年降水量增加而显著增加的格局主要是降水因子决定的。
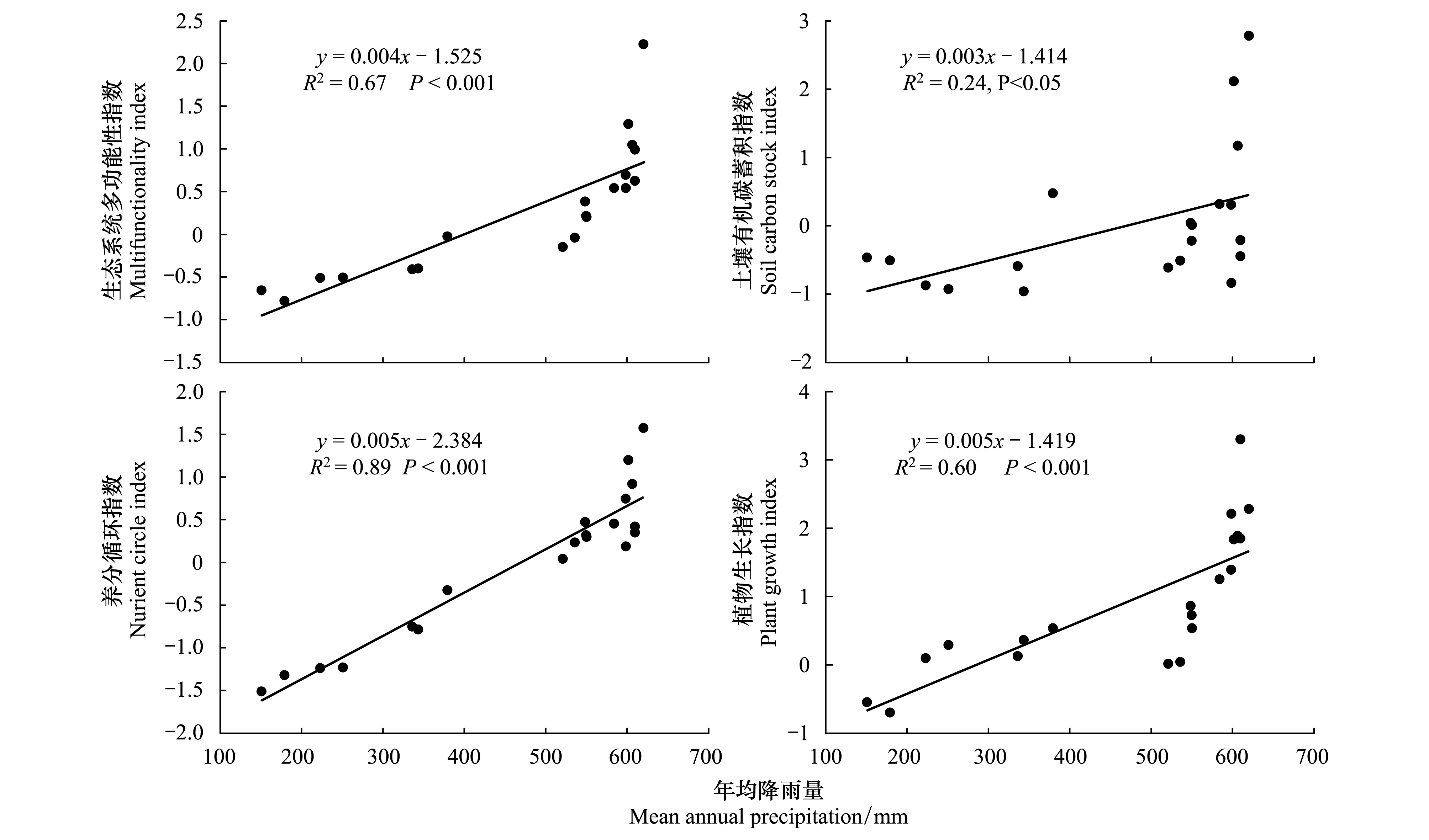
图3 高寒草地物种多样性与年降水量的关系Fig.3 Relationship between ecosystem multifunctionality and mean annual precipitation in the alpine grasslands
3讨论
在藏北羌塘高寒草地,不同物种多样性指数与草地生态系统多功能性指数间关系格局存在较大差异。本研究发现物种丰富度指数与多功能性指数间线性回归关系最显著,而Pielou均匀度指数与生态系统多能性指数关系不显著。物种丰富度指数代表了植物群落内物种数目的多少,均匀度指数则体现了各物种个体数目分配的均匀程度。因此,这一结果说明,藏北羌塘高寒草地生态系统多功能性主要受群落物种数量而非物种分布的均匀程度影响。在羌塘高原,草地生态系统同时维持和提供多种生态系统功能和服务需要较高的植物种数量来支撑。本研究结果与诸多基于区域或全球尺度的研究相一致,例如,Hector[9]等基于欧洲BIODEPTH草地生物多样性-生态系统功能实验数据的研究结果,Zavaleta[12]等基于全球干旱地区实验数据的研究以及Lefcheck[27]等在全球尺度上跨越不同营养级和生境的整合分析研究。这些研究都表明物种丰富度的增加对生态系统多功能性具有正效应。Shannon-wienner和Simpson多样性指数指数通过对群落的丰富度和均匀度进行综合评价来表征群落的复杂程度,本研究发现两种指数也与生态系统多功能性指数间成显著的正相关关系(P<0.05)。这种正相关关系表明,生态系统的多功能性主要受物种丰富度的影响。
本研究中,植物初级生产力、土壤碳蓄积,以及养分循环这3类重要的生态系统功能指数均随物种多样性指数特别是物种丰富度的增加而显著增加。研究结果与大多数的物种多样性-生产力试验结果相一致,即物种丰富的群落具有更高的群落生产力[1,3]。对土壤有机碳固持而言,物种丰富度的增加通常通过提高地上、地下生物量生产力[28]和根系凋落物碳库来增加土壤碳累积[29],另一方面,物种多样性能够通过改变凋落物的构成和多样性以及土壤的生物/非生物属性来影响土壤有机质分解[29],从而提高土壤有机碳蓄积。有研究表明,植物多样性越高,会增加根际微生物碳源的输入,从而导致微生物活性增强和凋落物分解,有利于碳储量增加[30]。物种丰富度与土壤养分循环指数间的正相关关系可能是由于较高的物种丰富度增加了土壤潜在氮矿化效率[31],同时较高的土壤有机碳蓄积有利于土壤中氨离子的交换,因此较高的土壤储量通常与较高土壤N矿化效率相联系[31-32]。这表明本研究中3种重要的生态系统功能/过程间存在一定的协同效应,维持较高的物种多样性水平不但能够提高藏北羌塘地区高寒草地生态系统多功能性,而且可以同时最大化这些重要的生态系统功能和服务。Wu[22]等人关于藏北羌塘高原草地地上生物量-物种丰富度关系的研究揭示了与以往诸多研究相一致的经典的单峰曲线格局。这一结果意味着:仅针对维持地上生产力水平而言,草地群落中某些植物种可能在功能上表现冗余,也就是对群落生产力的贡献不大。然而,从生态系统多功能性角度出发,本研究发现羌塘高寒草地生态系统多功能性随物种丰富度的增加呈显著线性增加趋势。这说明在羌塘高原,高寒草地生态系统维持多种功能或服务需要更多物种的参与,因为不同物种可能会在对维系不同种类的生态系统功能都产生贡献[9,13,33]。因次,孤立的考虑某一生态系统功能的研究可能会低估生物多样性的贡献[9,27]。由于不同生态系统功能之间可能存在的权衡关系,使用生态系统多功能性这一综合性概念有助于从整体上理解和认识生物多样性与生态系统功能间的关系格局[34]。
气候因子对于草地植物群落结构和物种丰富度的空间分布格局的形成具有重要意义[35],藏北羌塘草地样带横跨十个经度,年降水量从东至西由550mm递减至106mm,而在纬度上跨度很小,温度的影响很小。因此水分梯度变化可能是本研究中影响物种多样性变化的重要因素。研究结果表明,物种丰富度与年降水量间呈显著的正相关关系,这与杨元合等[36]对整个青藏高原高寒草地样带和胡云峰等[37]对内蒙古东北-西南草地样带多样性-降水量关系的研究结果相符。其他与均匀度紧密相关的多样性指数均与年降雨量间无明显的相关关系,表明均匀度更大程度上受到群落结构自身的影响,而对降水梯度变化不太敏感[37]。本研究中生态系统多功能性指数以及植被生产力、土壤碳储量和养分循环相关的参数均与年降水量间呈极显著的正相关关系,表明水分条件是决定羌塘高寒草原植物生长、养分循环和碳蓄积的主要限制因子。
藏北羌塘地区气候条件恶劣,环境脆弱,其草地对气候变化和人类活动的干扰响应极为敏感,生态系统脆弱。因此,藏北高原高寒草地由于过度放牧利用和气候变化的影响,威胁到群落的多样性,从而对生态系统多功能性造成显著地影响[38]。本研究结果强调了物种丰富度,即群落的物种数目对于维持藏北高寒草地生态系统多功能性的重要意义。较高的物种多样性水平有利于提高植物生产力、土壤有机碳固定以及土壤养分循环。相应的,物种丧失可能会给此区域带来更加严重的生态后果,威胁当地牧民的赖以生存的物质经济基础,以及野生动物的生存。因此,在退化草地恢复过程中,生物多样性水平特别是本地植物种丰富度的恢复应当作为一个重要目标。就草地可持续管理而言,应多从生态系统多功能性角度出发,而不是单纯考虑某一生态系统功能或服务的实现。
4结论
本研究表明,物种丰富度水平对于维持羌塘高寒草地生态系统多功能性具有重要意义,物种丰富度可以同时最大化藏北高寒草地的主要生态系统功能和服务。孤立的考虑某一生态系统功能的研究可能会低估生物多样性的贡献。由于人类活动导致的物种丧失可能会给藏北羌塘草原带来更加严重的生态后果,生物多样性水平(特别是本地物种丰富度)的恢复和维持应当作为该地区退化草地恢复的重要目标之一。
参考文献(References):
[1]Hector A, Schmid B, Beierkuhnlein C, CaldeiraM C, Diemer M, Dimitrakopoulos P G, Finn J A, Freitas H, Giller P S, Good J, Harris R, Högberg P, Huss-Danell K, Joshi J, Jumpponen A, Körner C, LeadleyP W, Loreau M, Minns A, Mulder C P H, O′Donovan G, Otway S J, Pereira J S, Prinz A, Read D J, Scherer-Lorenzen M, Schulze E D, Siamantziouras A S D, Spehn E M, Terry A C, Troumbis A Y, Woodward F I, Yachi S, Lawton J H. Plant diversity and productivity experiments in European grasslands. Science, 1999, 286(5442): 1123- 1127.
[2]Hooper D U, Chapin F S, Ewel J J, Hector A, InchaustiP, Lavorel S, Lawton J H, Lodge D M, Loreau M, Naeem S, Schmid B, Setälä H, SymstadA J, Vandermeer J, Wardle D A. Effects of biodiversity on ecosystem functioning: A consensus of current knowledge. Ecological Monographs, 2005, 75(1): 3- 35.
[3]Flombaum P, Sala O E. Higher effect of plant species diversity on productivity in natural than artificial ecosystems. Proceedings of the National Academy of Sciences of the United States of America, 2008, 105(16): 6087- 6090.
[4]张全国, 张大勇. 生物多样性与生态系统功能: 最新的进展与动向. 生物多样性, 2003, 11(5): 351- 363.
[5]Kennedy T, Naeem S, Howe K M, Knops J M H, Tilman D, Reich P. Biodiversity as a barrier to ecologicalinvasion. Nature, 2002, 417(6889): 636- 638.
[6]Srivastava D S, Velland M. Biodiversity-ecosystem function research: is it relevant toconservation? Annual Review of Ecology, Evolution, and Systematics, 2005, 36(1): 267- 294.
[7]Balvanera P, Pfisterer A B, Buchmann N, He JS, Nakashizuka T, Raffaelli D, Schmid B. Quantifying the evidence for biodiversity effects on ecosystem functioning and services. Ecology Letters, 2006, 9(10): 1146- 1156.
[8]Cardinale B J, Srivastava D S, Duffy J E,Wright J P, Downing A L, Sankaran M, Jouseau C. Effects of biodiversity on the functioning of trophic groups and ecosystems. Nature, 2006, 443(7114): 989- 992.
[9]Hector A, Bagchi R. Biodiversity and ecosystem multifunctionality. Nature, 2007, 448(7150): 188- 190.
[10]Maestre F T, Quero J L, Gotelli N J, Escudero A, Ochoa V, Delgado-Baquerizo M, Garcia-Gomez M, Bowker M A, Soliveres S, Escolar C, Garcia-Palacios P, Berdugo M, Valencia E, Gozalo B, Gallardo A, Aguilera L, Arredondo T, Blones J, Boeken B, Bran D, Conceicao A A, Cabrera O, Chaieb M, Derak M, Eldridge D J, Espinosa C I, Florentino A, Gaitan J, Gatica M G, Ghiloufi W, Gomez-Gonzalez S, Gutierrez J R, Hernandez R M, Huang X, Huber-Sannwald E, Jankju M, Miriti M, Monerris J, Mau R L, Morici E, Naseri K, Ospina A, Polo V, Prina A, Pucheta E, Ramirez-Collantes D A, Romao R, Tighe M, Torres-Diaz C, Val J, Veiga J P, Wang D, Zaady E. Plant species richness and ecosystem multifunctionality in global drylands. Science, 2012, 335(6065): 214- 218.
[11]Byrnes J E K, Gamfeldt L, Isbell F, Lefcheck J S, Griffin J N, Hector A, Cardinale B J, Hooper D U, Dee L E, Duffy J E, Freckleton R. Investigating the relationship between biodiversity and ecosystem multifunctionality: Challenges and solutions. Methods in Ecology and Evolution, 2014, 5(2): 111- 124.
[12]Zavaleta E S, Pasari J R, Hulvey K B, Tilman G D. Sustaining multiple ecosystem functions in grassland communities requires higher biodiversity. Proceedings of the National Academy of Sciences of the United States of America, 2010, 107(4): 1443- 1446.
[13]Gamfeldt L, Hillebrand H, Jonsson PR. Multiple functions increase the importance of biodiversity for overall ecosystem functioning. Ecology, 2008, 89(5): 1223- 1231.
[14]江娟. 北京市延庆县森林资源及其多功能效益评价[D]. 北京: 中国林业科学研究院, 2010.
[15]彭建, 刘志聪, 刘焱序. 农业多功能性评价研究进展. 中国农业资源与区划, 2014, 35(6): 1- 8.
[16]吴晶晶. 林农复合经营系统多功能性分析及评价研究[D]. 南京: 南京林业大学, 2013.
[17]王景升, 张宪洲, 赵玉萍, 沈振西, 石培礼, 余成群. 藏北羌塘高原气候变化的时空格局. 资源科学, 2008, 30(12): 1852- 1859.
[18]李明森. 藏北高原草地资源合理利用. 自然资源学报, 2000, 15(4): 335- 339.
[19]刘兴元, 龙瑞军, 尚占环. 草地生态系统服务功能及其价值评估方法研究. 草业学报, 2011, 20(1): 167- 174.
[20]宋春桥, 游松财, 柯灵红, 刘高焕, 钟新科. 藏北高原地表覆盖时空动态及其对气候变化的响应. 应用生态学报, 2011, 22(8): 2091- 2097.
[21]孙鸿烈, 郑度, 姚檀栋, 张镱锂. 青藏高原国家生态安全屏障保护与建设. 地理学报, 2012, 67(1): 3- 12.
[22]Wu J S, Shen Z X, Zhang X Z. Precipitation and species composition primarily determinethe diversity-productivity relationship of alpine grasslandson the Northern Tibetan Plateau. Alpine Botany, 2014, 124(1): 13- 25.
[23]武建双, 李晓佳, 沈振西, 张宪洲, 石培礼, 余成群, 王景升, 周宇庭. 藏北高寒草地样带物种多样性沿降水梯度的分布格局. 草业学报, 2012, 21(3): 17- 25.
[24]中国科学院南京土壤研究所. 土壤理化分析. 上海: 上海科技出版社, 1978.
[25]陈杰. 贵州省植烟土壤砾石含量分布. 安徽农业科学,2007, 35(32): 10334- 10335.
[26]Tilman D, Elhaddi A. Drought and biodiversity in grasslands. Oecologia, 1992, 89(2): 257- 264.
[27]Lefcheck J S, Byrnes J E K, Isbell F, Gamfeldt L, Griffin J N, Eisenhauer N, Hensel M J S, Hector A, CardinaleB J, Duffy J E. Biodiversity enhances ecosystem multifunctionality across trophic levels and habitats. Nature Communication, 2015, 6: 6936.
[28]FornaraDA, Tilman D. Plant functional composition influences rates of soilcarbon and nitrogen accumulation. Journal of Ecology, 2008, 96(2): 314- 322.
[29]Cong W F, Van Ruijven J, Van Der Werf W, De Deyn G B, Mommer L, Berendse F, Hoffland E. Plant species richness leaves a legacy of enhanced root litter-induceddecomposition in soil. Soil Biology and Biochemistry, 2015, 80: 341- 348.
[30]Lange M, Eisenhauer N, Sierra CA, Bessler H, Engels C, Griffiths RI, Mellado-VazquezPG, Malik AA, Roy J, Scheu S, Steinbeiss S,Thomson B C,Trumbore S E, Gleixner G. Plant diversity increases soil microbial activity and soil carbon storage. Nature Communications, 2014, 6: 6707.
[31]Mueller K, Hobbie S E, Tilman D, Reich P B. Effects of plant diversity, N fertilization, and elevatedcarbon dioxide on grassland soil N cycling in along-term experiment. Global Change Biology, 2013, 19(4): 1249- 1261.
[32]Cong W F, Van Ruijven J, Mommer L, De Deyn G B, Berendse F, Hoffland E, Lavorel S. Plant species richness promotes soil carbon andnitrogen stocks in grasslands without legumes. Journal of Ecology, 2014, 102(5): 1163- 1170.
[33]Isbell F, Calcagno V, Hector A, Connolly J, Harpole W S, Reich P B, Scherer-Lorenzen M, Schmid B, Tilman D, Van RuijvenJ, Weigelt A, Wilsey B J, Zavaleta E S, Loreau M.High plant diversity is needed to maintain ecosystem services. Nature, 2011, 477(7363): 199- 202.
[34]Byrnes J, Lefcheck J S, Gamfeldt L, Griffin JN, Isbell F, Hector A. Multifunctionality does not imply that all functions are positively correlated. Proceedings of the National Academy of Sciences of the United States of America, 2014, 111(51): E5490.
[35]Cleland EE, CollinsSL, Dickson TL, FarrerEC, GrossKL, Gherardi L A, Hallett L M, Hobbs R J, Hsu J S, Turnbull L, Suding K N. Sensitivity of grassland plant community compositionto spatial vs. temporal variation in precipitation. Ecology, 2013, 94(8): 1687- 1696.
[36]杨元合, 饶胜, 胡会峰, 陈安平, 吉成均, 朱彪, 左闻韵, 李轩然, 沈海花, 王志恒, 唐艳鸿, 方精云. 青藏高原高寒草地植物物种丰富度及其与环境因子和生物量的关系. 生物多样性, 2004, 12(1): 200- 205.
[37]胡云锋, 艳燕, 阿拉腾图雅, 毕力格吉夫. 内蒙古东北-西南草地样带植物多样性变化. 资源科学, 2012, 34(6): 1024- 1031.
[38]Gao Q Z, Li Y, Wan Y F, Jiangcun W Z, Qin X B, Wang B S. Significant achievements in protection and restoration of alpine grassland ecosystem in northern Tibet, China. Restoration Ecology, 2009, 17(3): 320- 323.
基金项目:国家自然科学基金项目(41271067); 中国科学院战略性先导专项课题(XDA05060700)
收稿日期:2015- 06- 30;
修订日期:2015- 11- 13
*通讯作者
Corresponding author.E-mail: shipl@igsnrr.ac.cn
DOI:10.5846/stxb201506301361
The relationship between species diversity and ecosystem multifunctionality in alpine grasslands on the Tibetan Changtang Plateau
XIONG Dingpeng1,2, ZHAO Guangshuai1,2, WU Jianshuang1, SHI Peili1,*,ZHANG Xianzhou1
1KeyLaboratoryofEcosystemNetworkObservationandModeling,InstituteofGeographicSciencesandNaturalResourcesResearch,Chinese,Beijing100101,China2UniversityofChineseAcademyofSciences,Beijing100049,China
Abstract:The classic species diversity-ecosystem function relationship studies attached more importance on single function and its correlation with diversity despite the fact that most ecosystems are managed or valued for their ability to maintain multiple goods and services simultaneously, so called multifunctionality. Therefore, multifunctionality is an important perspective for biodiversity-function research in point of view of comprehensive function analysis. We conducted an ecosystem multifunctionality vs. biodiversity study on the Tibetan Changtang Plateau, in terms of seven indicators of ecosystem functioning related to plant growth (aboveground biomass, belowground biomass), nutrient cycling (soil total nitrogen content, soil and content, soil phosphorus content), and soil carbon storage (soil organic carbon density). Z scores (standardized deviates) of the seven indicators were assessed to represent multifunctionality. The multifunctionality index (M) was calculated as the average Z-score for all indicators measured in each site. Linear regression was used to analyze the relationship between multifunctionality and species diversity indices (Species richness, Shannon-wiener index, Simpson dominance index, Pielou evenness index). We also tested whether ecosystem multifunctionality and species diversity were associated with climatic factors (mean annual temperature and mean annual precipitation) by using linear regression. It was indicated that ecosystem multifunctionality indices were correlated most significantly with species richness, notable positive with Shannon-wiener and Simpson index, but not with Pielou evenness index. Separate analyses of functions related to the plant growth, soil carbon storage and nutrient cycling also yield significantly positive relationships with species richness. Precipitation plays a positive role in affecting species diversity and ecosystem multifunctionality indices, as the species diversity and multiple functions increase linearly with mean annual precipitation. The present study highlights the importance of species richness in maintaining multiple ecosystem functions, goods and services in Changtang Plateau. Because each species contribute to different functions, studies with focus on single function (i.e. net production) might underestimate levels of species richness required to maintain multiple ecosystem goods and services. The alpine grassland ecosystem is vulnerable to climate change and human induced biodiversity loss. It is therefore implied that species loss may cause severe consequence for ecosystem functioning, which would threaten ecosystem health and human well-being in this region. We suggest that the species richness of native species must be maintained and sustained in order to meet the goal of degraded grassland restoration and sustainable management on the Changtang Plateau.
Key Words:Tibetan Changtang Plateau; alpine grasslands; species diversity; species richness; ecosystem multifunctionality
熊定鹏,赵广帅,武建双,石培礼,张宪洲.羌塘高寒草地物种多样性与生态系统多功能关系格局.生态学报,2016,36(11):3362- 3371.
Xiong D P, Zhao G S, Wu J S, Shi P L,Zhang X Z.The relationship between species diversity and ecosystem multifunctionality in alpine grasslands on the Tibetan Changtang Plateau.Acta Ecologica Sinica,2016,36(11):3362- 3371.
