小麦CBL结合蛋白激酶基因TaCIPK16的克隆及特征分析
2016-08-06MYOTHWIN薛庆贺康振生
MYO THWIN,刘 芃,马 微,薛庆贺,郭 军,康振生
(西北农林科技大学 植物保护学院,旱区作物逆境生物学国家重点实验室,陕西杨陵 712100)
小麦CBL结合蛋白激酶基因TaCIPK16的克隆及特征分析
MYO THWIN,刘芃,马微,薛庆贺,郭军,康振生*
(西北农林科技大学 植物保护学院,旱区作物逆境生物学国家重点实验室,陕西杨陵 712100)
摘要:类钙调磷酸酶亚基B蛋白(calcineurin B-1ike protein,CBL)作为一类钙离子结合蛋白,通过与一类蛋白激酶(CBL-interacting protein kinase,ClPK)结合,从而在钙信号依赖的生理生化过程中发挥作用。该研究在条锈菌诱导的小麦叶片中克隆获得CIPK家族中1个基因TaCIPK16,并利用qRT-PCR技术、酵母双杂交技术及亚细胞定位技术分析了其功能特性。序列分析表明,TaCIPK16编码447个氨基酸,包含保守的激酶催化结构域及调控结构域,与水稻、拟南芥CIPK蛋白具有高度相似性。酵母双杂交分析验证显示,TaCIPK16与TaCBL4和TaCBL9存在强烈互作。定量分析表明,TaCIPK16受到条锈菌的诱导表达,在小麦与条锈菌互作过程中呈显著差异表达趋势。综上结果,TaCIPK16可能作为正调控因子参与了小麦对条锈菌的抗病防卫反应。
关键词:小麦,CBL,蛋白激酶,TaCIPK16,条锈菌
在适应复杂多变自然环境的进化中,植物形成了完善的信号通路。Ca2+是信号传导中重要的第二信使,在植物的生理和生长过程中起着重要的作用。植物在响应外界的信号和胁迫时,细胞中的Ca2+水平会发生相应的改变[1]。植物中存在3种钙离子结合蛋白,分别是钙调素CaM (Calmodulin)及其相关蛋白、钙依赖性蛋白激酶(Ca2+-dependent protein kinases, CDPK)、钙调磷酸酶B类蛋白(calcineurin B-like protein, CBL)[2]。其中CBL本身没有激酶活性,必须和靶蛋白CIPK结合形成复合体才能发挥作用[3]。CIPK是一个植物特有的丝氨酸-苏氨酸蛋白激酶家族,属于第3类的SnRK3 激酶(SNF1-related protein kinase3, SnRK3)[4]。对模式植物拟南芥CIPK蛋白的结构分析表明,所有的CIPK蛋白含有N-端的激酶蛋白域和C-端的调节域,后者为CBL特异接合的NAF结构域[5]和能与PP2C相互作用的PPI结构域[6]。
前期研究表明CIPK家族基因参与响应高盐、渗透或者干旱胁迫,冷胁迫,以及低K+、硝酸盐、低氧等其它胁迫[7]。在拟南芥中,AtCIPK23 可以与 AtCBL1互作,磷酸激活 K+通道 (AKT1)促进 K+的吸收[8]。当拟南芥遭受高盐胁迫时, SOS3(CBL4)及CBL10结合Ca2+, 随后与SOS2(CIPK24)蛋白激酶结合形成蛋白复合体, 分别在地下及地上部分调控相应靶蛋白的表达, 从而使植物能抵御或减轻高盐胁迫带来的伤害[9]。在水稻中,通过RNAi技术沉默OsCIPK23能够导致水稻抗旱性的减弱[10]。除此之外,CBL-CIPK信号系统广泛参与植物生理和发育的过程。水稻中OsCIPK15与水稻的缺氧耐性有关,可能调节SnRK1A整合缺氧响应和糖信号反应[11]。在CIPK6功能缺失的突变体cipk6中,向基式与向顶式的生长素运输均明显减弱,造成植株表现为子叶融合、下胚轴膨大、侧根发生延迟等生长缺陷[12]。
除此之外,关于CIPK蛋白激酶在植物响应生物胁迫过程中的作用也有了一定的研究。众所周知,活性氧(reactive oxygen species, ROS)在植物抗病反应中起到至关重要的作用,它能够引起细胞坏死,从而限制病原菌的进一步扩展,并且可作为一个信号分子诱导抗病相关基因的表达[13]。在拟南芥中,AtCIPK26能够和一个NADPH氧化酶AtRbohF互作并负调控其产生ROS[14]。在水稻中,OsCIPK14/15能都受到病原相关分子模式(microbe-associated molecular patterns,MAMP)的诱导,若OsCIPK14/15的表达受到抑制,MAMP诱导产生的ROS也受到抑制,从而说明OsCIPK14/15参与了PAMP诱导的抗性(PAMP-triggered immunity, PTI)[15]。在番茄中,CBL10和CIPK6能够正调控细胞免疫反应和细胞程序性死亡。在烟草中,CIPK6与CBL10的互作参与ETI(effector-triggered immunity)过程中ROS的产生[16]。有研究表明,水杨酸信号通路也受到了CIPK的调节。在拟南芥中,NPR1能够被CIPK11/PKS5磷酸化,从而诱导WKY38和WKY62的表达[17]。
目前,小麦中关于CIPK蛋白激酶在生物胁迫中的功能知之甚少。研究小麦CIPK蛋白激酶在小麦-条锈菌互作体系的功能对阐释其作用机理具有重要的意义。因此,本研究分离获得了一个蛋白激酶基因TaCIPK16,并利用qRT-PCR技术、酵母双杂交技术及亚细胞定位技术分析了其功能特性,为进一步揭示其在小麦-条锈菌互作过程中的分子机理奠定了基础。
1材料与方法
1.1供试材料及试验试剂
本实验所用小麦条锈菌生理小种为‘条中23号’(CYR23)和‘条中31号’(CYR31),小麦品种为‘铭贤169’和‘水源11’。小麦培养、条锈菌繁殖及接种方法参考Kang等[18]的方法。Taq DNA 聚合酶(Fermentas,美国),dNTP、pMD18-T(Takara,日本),琼脂糖凝胶 DNA 回收试剂盒(百泰克,中国),Top10感受态细胞(Wolsen,美国),质粒提取盒(BioMiga,美国),限制性内切酶(Fermentas,美国),T4 DNA连接酶(Fermentas),BIOZOL(BioFlux,日本),RNase inhibitor(Promega,美国),M-MLV反转录试剂盒(Fermentas,美国),SY BR Green 和 Rox reference 染料(Invitrogen,美国)。
1.2方法
1.2.1TaCIPK16基因的克隆在TAIR v10 (http://www.arabidopsis.org/)和RGAP v7 (http://rice.plantbiology.msu.edu/)数据库中得到拟南芥和水稻的CIPK序列,在中国春基因组数据库中(http://wheat-urgi.versailles.inra.fr/Seq-Repository/Genes-annotations)得到小麦cDNA数据库,并以水稻和拟南芥的CIPK序列为种子序列,参照NCBI“BLAST+ user manual”中的方法,对下载的小麦cDNA数据库进行本地BLAST检索,得到小麦候选的CIPK序列,并通过MEGA5.0构建进化树明确与OsCIPK16亲缘关系最近的序列,即为候选的TaCIPK16。根据序列分析得到的候选基因序列设计引物TaCIPK16-F/R(表 1)。提取水源11叶片总 RNA 并反转录成 cDNA,具体方法参照 Ferments ReverAid First Strand cDNA synthesis 试剂盒操作说明。以 cDNA 为模板,PCR 扩增程序为95 ℃预变性 5 min,95 ℃ 变性30 s,55 ℃ 退火30 s,72 ℃延伸 1 min,共 35 个循环;72 ℃ 延伸10 min。PCR 产物纯化后连接到pGEM®-T Easy载体,经大肠杆菌转化后提取质粒并送测序。
1.2.2序列分析将实验克隆测序的结果和通过预测得到的候选基因序列进行相似性比对。使用ExPAsy网站的在线软件Compute pI/Mw tool(http://web.expasy.org/compute_pi/)进行计算编码蛋白的等电点和分子量;利用DNASTAR软件,BLAST (http://www.ncbi.nlm.nih.gov/blast/)和ORF Finder (http://www.ncbi.nlm.nih.gov/gorf/gorf.html)进行cDNA序列的分析。利用ClustalW2 (http://www.ebi.ac.uk/Tools/msa/muscle/) 和 DNAMAN7.0 软件(version 7.0; Lynnon Biosoft, USA)进行序列比对分析。使用 MEGA5 software (version 5.0)软件以邻接法(Neighbor-Joining method)进行目的蛋白与其同源蛋白的多重序列比对及进化树分析。使用InterProScan (http://www.ebi.ac.uk/Tools/pfa/iprscan/) 和 PROSITE Scan (http://prosite.expasy.org/scanprosite/) 来预测蛋白的保守蛋白结构域。
1.2.3酵母双杂交分析设计并合成TaCBL4(GenBank登录号为KU736850)和TaCBL9(GenBank登录号为KU736852)酵母双杂交克隆引物TaCBL4-BD-F/R、TaCBL9-BD-F/R(表 1),并克隆至pGBKT7载体上,设计TaCIPK16酵母双杂交克隆引物TaCIPK16-AD-F/R(表 1),并克隆至pGADT7载体上。按照酵母双杂交手册(Clontech, Japan),将TaCBL4与TaCIPK16或者TaCBL9与TaCIPK16共转至酵母菌株AH109中,并分别在缺陷性培养基SD-Leucine-Tryptophan (-L-T)和SD-Ade-nine-Histidine-Leucine-Tryptophan(含20 μg·mL-1的X-α-Gal)(-L-T-H-A+ X-α-gal)上进行培养。培养4 d后,利用血球计数板将酵母细胞稀释到每毫升水含有107、106、105和104个细胞,点至-L-T-H-A+ X-α-gal培养基上培养,24 h后照相。
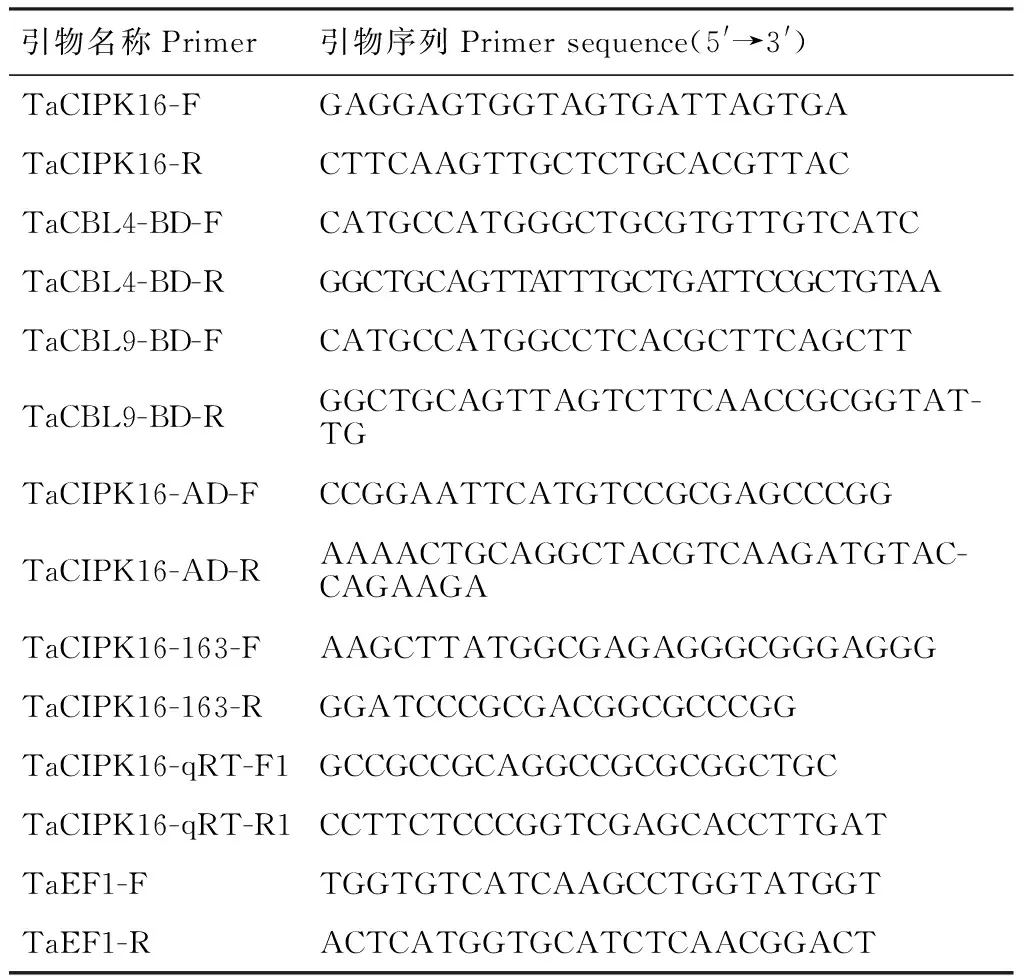
表1 引物表
1.2.4亚细胞定位依据pCaMV35S:GFP中多克隆位点信息,设计并合成TaCIPK16亚细胞定位克隆引物TaCIPK16-163-F/R(表1),并将TaCIPK16克隆至pCaMV35S:GFP中,参照Li 等[19]的方法,制备小麦原生质体,并参照该方法将融合载体pCaMV35S:TaCIPK16∶GFP及空载体pCaMV35S∶GFP转至小麦原生质体中,培养18 h后,通过共聚焦显微镜进行观察并照相。
1.2.5qRT-PCR分析根据克隆出来的TaCIPK16 基因和内参基因小麦延伸因子TaEF-1α(GenBank登录号Q03033)分别设计特异定量引物TaCIPK16-qRT-F1/R1及TaEF1-F/R(表 1)。采集接种条锈菌后 0、12、18、24、72和120 h 时间点的叶片提取 RNA,反转录为 cDNA,以此 cDNA为模板,qRT-PCR 反应程序为95 ℃预变性5 min,95 ℃变性,10 s,55 ℃ 退火 20 s,72 ℃延伸 40 s,共40个循环。qRT-PCR的反应体系及程序参照Feng等的方法[20],最后采用2-ΔΔCt法计算目标基因对内参基因的相对表达量。
2结果与分析
2.1小麦TaCIPK16基因的克隆及序列分析
通过RT-PCR技术克隆得到的cDNA序列具有1 606个核苷酸(图1),编码1个447氨基酸的蛋白,蛋白分子量为 49.521 kD ,等电点为9.09。使用InterProScan软件分析表明,其N端包含一个蛋白激酶结构域,C端含有一个调控结构域, 在调节域中包含能够与CBL 特异结合的NAF结构域及与PP2C相互作用的PPI结构域(图 2)。利用拟南芥与水稻中所有的CIPK序列氨基酸序构建进化树发现该蛋白与水稻的OsCIPK16(LOC_Os09g25090)及拟南芥中的AtCIPK5(AAF86504)、AtCIPK16(AAK50348)及AtCIPK25(AF44226)同属一个分支(图 3)。5条序列比对分析表明,该蛋白序列与OsCIPK16相似度达到88.04%,与拟南芥(Arabidopsisthaliana)中AtCIPK16相似度达到76.43%,故命名为TaCIPK16。

1~2. PCR扩增产物;M. Marker图1 TaCIPK16基因cDNA的PCR扩增片段1-2. PCR product; M. MarkerFig. 1 Electrophoresis of TaCIPK16 cDNA fragment

At. 拟南芥CIPK序列;Os. 水稻CIPK序列;箭头指出本实验克隆的TaCIPK16序列;节点上的数值表示Bootstrap验证中基于1 000次重复该节点的可信度图3 TaCIPK16与水稻、拟南芥中CIPK家族的进化树分析At, CIPK sequences of Arabidopsis download from TAIR v10; Os, CIPK sequences of rice download from RGAP v7; The arrow indicates TaCIPK16; The numbers at the nodes represents the reliability based on 1 000 replicatesFig. 3 Phylogenetic analysis of TaCIPK16 with CIPK family from rice and Arabidopsis

虚线表示N端蛋白激酶结构域;黑线表示C端调控结构域;黑框表示NAF结构域;虚线框表示PPI结构域图2 TaCIPK16与水稻、拟南芥同源基因的多序列比对The conserved protein kinase domain and regulatory domain are marked with black full line and dash line, respectively. The conserved NAF and PPI motifs are marked with black box and dash boxFig. 2 Multi-sequence alignment of TaCIPK16 with homologous proteins in rice and Arabidopsis
2.2酵母双杂分析
为了验证 TaCIPK16和TaCBL4或TaCBL9是否有相互作用,将TaCIPK16与TaCBL4或者TaCIPK16与TaCBL9共同转至酵母中。在筛选培养基SD-Leucine-Tryptophan上酵母能够正常生长,说明2种融合质粒成功转入酵母中(图 4,A)。在筛选培养基SD-Adenine-Histidine-Leucine-Tryptophan(含20μg·mL-1X-α-Gal)酵母能够生长并且成蓝色,与阳性对照相似,说明TaCIPK16与TaCBL4和TaCBL9均存在互作(图 4,B)。将酵母培养液稀释到每毫升含有104个酵母细胞时,仍能观察到酵母的生长,表明TaCIPK16与TaCBL4和TaCBL9具有强烈的互作。

含有重组质粒的AH109酵母菌株在选择培养基SD-Leucine-Tryptophan (-L-T)和SD-Adenine-Histidine-Leucine-Tryptophan(加20 μg/mL的X-α-gal)培养;A、B.阳性对照,Takala酵母试剂盒中的SV40 large T-antigen和murine p53互作结果:C、D.TaCIPK16和TaCBL4的互作结果;E、F.TaCIPK16和TaCBL9的互作结果;G、H.阴性对照,Takala酵母试剂盒中SV40 large T-antigen 与human lamin的互作结果图4 TaCIPK16与TaCBL4、TaCBL9的互作分析Cells of yeast strain AH109 harboring the indicated plasmid combinations were grown on selective media SD-Leucine-Tryptophan (-L-T) and SD-Adenine-Histidine-Leucine-Tryptophan (containing 20 μg/mL X-α-gal); A, B. Positive control, the interaction between SV40 large T-antigen and murine p53;C, D. The interaction of TaCIPK16 and TaCBL4: E,F. The interaction between TaCIPK16 and TaCBL9; G,H. Negative control, the interaction between SV40 large T-antigen and human lamin C.Fig. 4 Analysis of interactions between TaCIPK16 and TaCBL4/TaCBL9

A~D.阳性对照,pCaMV35S:GFP转化的小麦原生质细胞;E~H.pCaMV35S:TaCIPK16:GFP转化的小麦原生质细胞;A、E.受激发的GFP荧光和叶绿体自发荧光组合结果;B、F.叶绿体自发荧光;C、G.受激发的GFP荧光;D、H.白光下的原生质细胞图5 小麦原生质体中TaCIPK16亚细胞定位分析(Bar= 5 μm)A~D. Positive control, pCaMV35S:GFP; E~H. pCaMV35S:TaCIPK16:GFP; A, E. overlay; B, F. Chloroplast; C, G. GFP; D, H. Bright field.Fig. 5 Subcellular localization of TaCIPK16 in wheat protoplasts

图6 TaCIPK16基因在小麦-条锈菌互作过程中的转录表达特征Fig. 6 Expression pattern of TaCIPK16 during the compatible and incompatible interaction between wheat and Pst
2.3亚细胞定位分析
为验证TaCIPK16在小麦细胞中的分布情况,将融合载体pCaMV35S∶TaCIPK16∶GFP以及空载体pCaMV35S∶GFP转移至小麦原生质体中。结果表明,TaCIPK16在小麦细胞中分布与阳性对照GFP的情况相同,在整个小麦细胞中均有分布(图 5)。
2.4小麦TaCIPK16基因的实时定量RT-PCR分析
实时定量RT-PCR分析结果表明,TaCIPK16受到条锈菌诱导表达。在非亲和互作(CYR23)中,TaCIPK16转录水平在接种后6~36 h低于0 h表达水平,呈下调表达;当到48 h表达量明显上调,并达到峰值(相对表达量为10.8),在以后的时间点,表达量恢复到初始水平(图 6)。在亲和互作(CYR31)中,TaCIPK16的相对表达趋势与在非亲和中的表达趋势相同,但在接菌48 h后轻微的上调表达量(相对表达量为2)显著低于在非亲和互作中表达量(图 6)。由此推断,TaCIPK16可能参与了小麦对条锈菌的抗病防卫反应。
3讨论
在已有的研究中,植物CIPK基因家族是一个较大的家族,家族内不同的CIPK成员的功能存在特异性,可以响应一种或者多种生物或非生物胁迫。本研究克隆了1个小麦中编码CIPK蛋白激酶的基因,其与拟南芥和水稻中的AtCIPK16和OsCIPK16具有高度的相似性,尤其是N-端的催化结构域和C端的调节结构域,推测TaCIPK16能够感知Ca2+信号,并且功能与水稻和拟南芥中的CIPK16蛋白功能高度保守。
CBL蛋白需要与CIPK蛋白激酶结合形成蛋白复合体,能激活细胞内一系列的生理生化的变化。以前研究表明,不同的CBL蛋白与不同的CIPK蛋白结合能够产生不同的信号通路,并且在不同的信号通路之间,CBL-CIPK蛋白复合体也起到桥梁的作用[21]。因此,明确CIPK与CBL的互作网络是研究CIPK蛋白作用机理的基础。本研究中通过酵母双杂技术,明确TaCIPK16与TaCBL4和TaCBL9有强烈的互作关系。由此推断,TaCIPK16能够响应多种环境对植物的刺激,并且能与小麦CBL形成不同蛋白复合体发挥其特异功能。
分析CIPK在细胞中的分布情况有助于理解CBL-CIPK蛋白复合体的功能及其特异的钙离子信号通路。在拟南芥中CIPK家族中的大部分基因都分布在细胞核与细胞质中[22]。在玉米中,ZmCIPK16主要定位在细胞核和细胞膜上,在细胞质中含量相对较少[2]。在本实验中,TaCIPK16也分布于整个细胞中,推断TaCIPK16与ZmCIPK16功能具有保守性。同时,细胞中蛋白的定位情况和其功能关系密切相关,因此推测在细胞中TaCBL4-TaCIPK16与TaCBL9-TaCIPK16蛋白复合体能够激活多种蛋白所引起的信号通路。
TaCIPK16基因在条锈菌诱导下呈上调表达趋势,说明其参与了条锈菌诱导的小麦抗病防卫反应,但亲和反应和非亲和反应中的表达量差异说明TaCIPK16很可能参与了小麦R基因介导的抗病信号通路,在小麦-条锈菌组成的非亲和互作过程中起重要作用。根据Wang等[23]报道,在非亲和互作反应中,接种条锈菌前期12~24 h,是小麦细胞活性氧爆发阶段并在24 h时达到峰值,而接菌24 h之后活性氧的含量才开始降低,到接菌后48 h,活性氧的含量达到最低。而本实验中在接菌后12~36 hTaCIPK16的表达水平受到抑制,而在接菌后48 h TaCIPK16受到诱导且表达水平到最大。有研究表明,小麦TaCIPK29参与调控ROS信号通路[24]。因此推测TaCIPK16可能在小麦-条锈菌非亲和互作过程中调控ROS信号通路以此介导小麦的抗病防卫反应。
参考文献:
[1]ZHU JK. Salt and drought stress signal transduction in plants[J].AnnualReviewofPlantBiology. 2002, 53: 247.
[2]ZHAO J, SUN Z, ZHENG J,etal. Cloning and characterization of a novel CBL-interacting protein kinase from maize[J].PlantMolecularBiology, 2009, 69(6): 661-674.
[3]LUAN S, KUDLA J, RODRIGUEZ-CONCEPCION M,etal. Calmodulins and calcineurin B-like proteins calcium sensors for specific signal response coupling in plants[J].ThePlantCell, 2002, 14: S389-S400.
[4]KOLUKISAOGLUÜ, WEINL S, BLAZEVIC D,etal. Calcium sensors and their interacting protein kinases: genomics of theArabidopsisand rice CBL-CIPK signaling networks[J].PlantPhysiology, 2004, 134(1): 43-58.
[5]ALBRECHT V, RITZ O, LINDER S,etal. The NAF domain defines a novel protein-protein interaction module conserved in Ca2+regulated kinases[J].TheEMBOJournal, 2001, 20(5): 1 051-1 063.
[6]LEE S C, LAN W Z, KIM B G,etal. A protein phosphorylation/dephosphorylation network regulates a plant potassium channel[J].ProceedingsoftheNationalAcademyofSciences, 2007, 104(40): 15 959-15 964.
[7]LI R, ZHANG J, WEI J,etal. Functions and mechanisms of the CBL-CIPK signaling system in plant response to abiotic stress[J].ProgressinNaturalScience, 2009, 19(6): 667-676.
[8]LI L, KIM B G, CHEONG Y H,etal. A Ca2+signaling pathway regulates a K+channel for low-K response inArabidopsis[J].ProceedingsoftheNationalAcademyofSciences, 2006, 103(33): 12 625-12 630.
[9]XU J, LI H D, CHEN L Q,etal. A protein kinase, interacting with two calcineurin B-like proteins, regulates K+transporter AKT1 inArabidopsis[J].Cell, 2006, 125(7): 1 347-1 360.
[10]YANG W, KONG Z, OMO-IKERODAH E,etal. Calcineurin B-like interacting protein kinase OsCIPK23 functions in pollination and drought stress responses in rice (OryzasativaL.)[J].JournalofGeneticsandGenomics, 2008, 35(9): 531-543.
[11]LEE K W, CHEN P W, LU C A,etal. Coordinated responses to oxygen and sugar deficiency allow rice seedlings to tolerate flooding[J].ScienceSignaling, 2009, 2(91): ra61.
[12]TRIPATHI V, PARASURAMAN B, LAXMI A,etal. CIPK6, a CBL-interacting protein kinase is required for development and salt tolerance in plants[J].ThePlantJournal, 2009, 58(5): 778-790.
[13]MITTLER R, VANDERAUWERA S, SUZUKI N,etal. ROS signaling: the new wave?[J].TrendsinPlantScience, 2011, 16(6): 300-309.
[14]KIMURA S, KAWARAZAKI T, NIBORI H,etal. The CBL-interacting protein kinase CIPK26 is a novel interactor ofArabidopsisNADPH oxidase AtRbohF that negatively modulates its ROS-producing activity in a heterologous expression system[J].JournalofBiochemistry, 2013, 153(2): 191-195.
[15]KURUSU T, HAMADA J, NOKAJIMA H,etal. Regulation of microbe-associated molecular pattern-induced hypersensitive cell death, phytoalexin production, and defense gene expression by calcineurin B-like protein-interacting protein kinases, OsCIPK14/15, in rice cultured cells[J].PlantPhysiology, 2010, 153(2): 678-692.
[16]DE LA TORRE F, GUTIÉRREZ-BELTRN E, PAREJA-JAIME Y,etal. The tomato calcium sensor Cbl10 and its interacting protein kinase Cipk6 define a signaling pathway in plant immunity[J].ThePlantCell, 2013, 25(7): 2 748-2 764.
[17]XIE C, ZHOU X, DENG X,etal. PKS5, a SNF1-related kinase, interacts with and phosphorylates NPR1, and modulates expression of WRKY38 and WRKY62[J].JournalofGeneticsandGenomics, 2010, 37(6): 359-369.
[18]KANG Z S, LI Z Q. Discovery of a normal T. type new pathogenic strain to Lovrin10[J].ActaCllegiiSeptentrionaliOccidentaliAgriculturae, 1984, 4(1): 18-28.
[19]LI C, LIN H, DUBCOVSKY J. Factorial combinations of protein interactions generate a multiplicity of florigen activation complexes in wheat and barley[J].ThePlantJournal, 2015, 84(1): 70-82.
[20]FENG H, WANG X, SUN Y,etal. Cloning and characterization of a calcium binding EF-hand protein geneTaCab1 from wheat and its expression in response toPucciniastriiformisf. sp.triticiand abiotic stresses[J].MolecularBiologyReports, 2011, 38(6): 3 857-3 866.
[21]THODAY-KENNEDY E L, JACOBS A K, ROY S J. The role of the CBL-CIPK calcium signaling network in regulating ion transport in response to abiotic stress[J].PlantGrowthRegulation, 2015, 76(1): 3-12.
[23]WANG C F, HUANG L L, BUCHENAUER H,etal. Histochemical studies on the accumulation of reactive oxygen species (O2-and H2O2) in the incompatible and compatible interaction of wheat-Pucciniastriiformisf. sp.tritici[J].PhysiologicalandMolecularPlantPathology, 2007, 71(4): 230-239.
[24]DENG X, HU W, WEI S,etal.TaCIPK29, a CBL-interacting protein kinase gene from wheat, confers salt stress tolerance in transgenic tobacco[J].PLoSONE, 2013, 8(7): e69881.
(编辑:宋亚珍)
文章编号:1000-4025(2016)06-1073-07
doi:10.7606/j.issn.1000-4025.2016.06.1073
收稿日期:2016-04-27;修改稿收到日期:2016-05-23
基金项目:国家“973”计划(2013CB127700);国家自然科学基金(31371889);教育部新世纪优秀人才支持计划(NCET-12-0471);高等学校学科创新引智计划(B07049)
作者简介:MYO THWIN(1990-),男,硕士,主要从事植物免疫学研究。E-mail: myothwinpatho@gmail.com *通信作者:康振生,教授,博士生导师,主要从事小麦条锈病的防控研究。E-mail: kangzs@nwsuaf.edu.cn
中图分类号:Q785;Q786
文献标志码:A
Characterization of a CBL-interacting Protein Kinase GeneTaCIPK16 in Wheat
MYO THWIN, LIU Peng, MA Wei, XUE Qinghe, GUO Jun, KANG Zhensheng*
(State Key Laboratory of Crop Stress Biology for Arid Areas, College of Plant Protection, Northwest A&F University, Yangling, Shaanxi 712100, China)
Abstract:The calcineurin B-like (CBL)-CBL-interacting protein kinase (CIPK) network plays a pivotal role in regulating the physiological as well as developmental processes in plants. In this study, we obtained a CIPK gene, TaCIPK16, from the wheat leaves infected by Puccinia striiformis f. sp. tritici (Pst) using in silico cloning and RT-PCR. The full-length cDNA of TaCIPK16 was 1 606 bp, which encoding 433 amino acids. Multi-sequence alignment showed that TaCIPK16 share high similarity with OsCIPK16 and AtCIPK16 in rice and Arabidopsis, respectively. Analysis of the protein domain features indicated that TaCIPK16 contained conserved regulatory domain in C-terminal and protein kinases domain in N-terminal. Subcellular localization assays revealed that TaCIPK16 displayed a localization pattern similar to that of the GFP control, indicating a cytoplasmic and nuclear localization. Yeast two-hybrid assays showed that TaCIPK16 strongly interacts with TaCBL4 and TaCBL9, respectively. qRT-PCR assays indicated that TaCIPK16 was induced by Pst infection and differentially expressed during incompatible and compatible interactions between wheat and Pst. Our results suggest that TaCIPK16 has a positive role in regulating wheat resistance against Pst.
Key words:wheat; CBL; protein kinase; TaCIPK16; Puccinia striiformis f. sp. tritici
