促芽肥对头季稻灌浆后期叶片蛋白质表达的影响
2016-07-28陈鸿飞张志兴林文雄
陈鸿飞,张志兴,林文雄
(1.福建农林大学生命科学学院,福建福州 350002;2.福建农林大学农业生态研究所,福建福州 350002;3.福建农林大学作物生态与分子生理学福建省高校重点实验室,福建福州 350002)
促芽肥对头季稻灌浆后期叶片蛋白质表达的影响
陈鸿飞1,2,3,张志兴1,2,3,林文雄1,2,3
(1.福建农林大学生命科学学院,福建福州 350002;2.福建农林大学农业生态研究所,福建福州 350002;3.福建农林大学作物生态与分子生理学福建省高校重点实验室,福建福州 350002)
为探究促芽肥对头季稻的调控效应,以Ⅱ优1273为材料,在头季稻齐穗后18 d设置施用促芽肥和未施用促芽肥2种处理,采用比较蛋白质组学的方法和技术,并结合相关生理指标测定,分析了促芽肥对头季稻灌浆后期叶片蛋白质表达及相应生理特性的影响。不同促芽肥处理的头季稻灌浆后期叶片蛋白质经双向电泳分离后共获得了15个差异表达的蛋白质点,涉及头季稻灌浆后期叶片的光合碳同化、电子传递与光合磷酸化和抗逆抗衰老响应等,分析结果表明,头季稻后期剑叶中与光合碳同化、电子传递与光合磷酸化相关蛋白随灌浆进程呈下降趋势,施用促芽肥能明显减缓其下调幅度,从而使头季稻灌浆后期剑叶具有相对较高的叶绿素含量和净光合速率,提高了头季稻灌浆后期的干物质积累和源供应能力;还能明显提高头季稻灌浆后期剑叶中与抗性相关蛋白的表达量,从而增强稻株的活性氧清除能力,抑制功能叶的膜脂过氧化作用,延缓头季稻灌浆后期剑叶的衰老速率,从而显著提高了头季稻的结实率和产量。
再生稻;叶片;促芽肥;蛋白质组学;产量
叶片是水稻最重要的光合源器官,籽粒灌浆所需物质的60%~80%来自于叶片的光合作用[1],翟虎渠等[2]研究认为决定产量差异的最主要时期不是在穗增重速率最快的灌浆前中期,而是在灌浆后期至成熟期,灌浆后期叶片的衰老导致光合作用下降,直接影响水稻的结实率和高产潜力的发挥[3];水稻叶片衰老速率除了受遗传因子影响外,很大程度上也受环境因素(如温度、光照、水分、营养胁迫等)的调控[4-8]。我们发现生产上高产再生稻的头季稻结实率(≥90%)明显较双季稻或单季稻高,分析2种水稻的栽培措施,头季稻齐穗后15~20 d时施用促芽肥是导致高产再生稻头季稻与单季稻或双季稻结实率差异的主要栽培措施,可见,促芽肥不但能促进再生芽的萌发[9],对头季稻也有着显著的影响。关于促芽肥对头季稻的影响,国内外众多学者从生理生态特性等方面进行了研究[10-12],但对于促芽肥调控头季稻灌浆后期叶片的分子机制仍有待进一步研究。蛋白质是生理功能的执行者,是生理代谢和基因间联系的重要桥梁,因而,蛋白质组学技术目前已被广泛应用于特定生理、生态环境下水稻代谢机制的研究[13-14];为了深入揭示促芽肥对头季稻的影响机理,本研究从蛋白质组学角度出发,分析促芽肥对头季稻灌浆后期叶片蛋白质表达特性的影响,以期为深入了解高产再生稻头季稻高结实率的形成及栽培调控提供理论依据和实践指导。
1 材料和方法
1.1 试验材料与设计
试验于2012年在福建农林大学教学农田进行,供试土壤为壤土,土壤基础肥力为含有机质25.53 g/kg、全氮2.02 g/kg、全磷0.65 g/kg、全钾0.75 g/kg、速效氮125.6 mg/kg、速效磷43.5 mg/kg、速效钾97.5 mg/kg,pH值5.75。
供试水稻品种为Ⅱ优1273,播种期为3月21日,移栽期为4月26日,栽插规格为16.5 cm×19.8 cm,头季稻齐穗期为7月6日,头季稻齐穗后18 d进行2种促芽肥处理(NA、NNA),NA处理:头季稻齐穗后18 d施300.00 kg/hm2尿素作为促芽肥,头季稻割后3 d施75.00 kg/hm2尿素作为壮苗肥;NNA处理:头季稻齐穗后18 d不施用促芽肥,头季稻割后3 d施75.00 kg/hm2尿素作为壮苗肥;每个处理设3个小区,每个小区面积16 m2,各小区周边用塑料材质制作的立板和农膜包被的土筑成田埂,2个处理除促芽肥外其他管理措施相同。
1.2 试验方法
1.2.1 取样 于头季稻抽穗开花期,每处理选取长势基本一致的70株稻株挂牌标记,自头季稻齐穗后18 d即促芽肥施用当天至头季稻成熟,每隔5 d于下午17:00左右剪挂牌标记15株稻株的剑叶,置于液氮中保存,用于蛋白质的提取和生理生化测定,共取样4次。
1.2.2 产量及其构成因子 头季成熟时每小区调查50株穗数,并取代表性的5丛稻株风干后考种,单株计产。
1.2.3 叶绿素含量 利用日产SPAD-502叶绿素测定仪自头季稻齐穗后18 d即促芽肥施用当天至头季稻成熟每隔5 d测定15株挂牌标记的水稻植株剑叶SPAD值,共4次,测定时避开叶脉分别测定剑叶的上中下3个部位,取其平均值。
1.2.4 净光合速率 用LI-6400便携式光合作用测定仪自头季稻齐穗后18 d,即促芽肥施用当天至头季稻成熟每隔5 d于上午9:00-11:00每处理测定15株挂牌标记的水稻植株剑叶净光合速率,共4次。
1.2.5 细胞保护酶活性和MDA含量的测定 细胞保护酶活性和MDA含量测定参照张志良[15]的方法。可溶性蛋白质含量测定采用考马斯亮蓝染色法,超氧化物歧化酶(SOD)活性测定采用氮蓝四唑还原法,过氧化氢酶(CAT)活性测定采用碘量法,过氧化物酶(POD)活性测定采用愈创木酚法,丙二醛(MDA)含量测定采用硫代巴比妥酸法,以样品中含有的蛋白质含量计算酶活性。
1.2.6 叶片蛋白质样品的制备 取0.5 g头季稻剑叶样品,参照邵彩虹等[16]的TCA-丙酮沉淀方法制备;蛋白质样品浓度测定采用Bradford方法进行[17]。
1.2.7 双向电泳与图谱分析 双向电泳参照邵彩虹等[16]的方法进行。等电聚焦(IEF)采用18 cm自制管状胶条,上样量为150μg,SDS-PAGE凝胶规格为180 mm×180 mm×1.5 mm,凝胶染色采用硝酸银染色法。
染色后采用Umax powerlookⅢ扫描仪进行扫描,构建蛋白表达图谱。利用ImageMaster 5.0软件对获得的剑叶蛋白表达图谱进行蛋白质表达差异分析,以不同蛋白表达图谱间同一蛋白质点表达丰度值差异(差异=(最大表达量-最小表达量)/最小表达量×100%)达50%以上即认为该蛋白点是差异表达蛋白点。
1.2.8 质谱分析 切取2个处理的差异表达蛋白质点,采用串联质谱(ESI-Q MS/MS、复旦大学蛋白组学研究中心)对差异表达蛋白质点进行质谱分析。
1.3 统计分析
采用Microsoft Excel 2007和统计分析软件SPSS 19.0对试验数据进行录入、处理和方差显著性分析。
2 结果与分析
2.1 促芽肥对头季稻产量及其构成因子的影响
由表1可知,头季稻产量施用促芽肥NA处理的为12 546.91 kg/hm2,未施促芽肥NNA处理的为11 355.15 kg/hm2,NA处理比NNA处理高10.50%,达显著差异水平,进一步对产量构成因子进行分析还可看出,头季稻2个处理的单位面积有效穗数、穗粒数和千粒质量基本相近,差异未达显著水平;但结实率NA处理的显著高于NNA处理,表明NA处理促进头季稻增产主要是在前期建立适宜穗数和大穗的基础上通过显著提高结实率来发挥品种高产潜力,从而显著提高了稻谷产量。

表1 不同促芽肥处理对头季稻产量及其构成因子的影响Tab.1 Effects of different nitrogen app lication for bud developm ent on grain yield and its com ponents in first cropping rice
2.2 头季稻灌浆后期叶片蛋白质表达图谱的构建与比较分析
比较NA和NNA 2个不同促芽肥水平处理下促芽肥施用当天至头季稻成熟的4个不同时期(施用促芽肥后0,5,10,15 d)头季稻剑叶叶片电泳图谱,在不同促芽肥处理下发现15个出现差异表达的蛋白质点,并以1~15进行编号(图1)。

图1 头季稻灌浆后期叶片的蛋白表达图谱Fig.1 The 2-DE m ap of flag leaf at late grain filling stage in the first cropping rice
2.3 差异表达蛋白质的质谱鉴定及功能分析
根据质谱鉴定数据,15个差异蛋白点中共有10个差异蛋白质点得到鉴定,通过MASCOT软件(Matrix Science,http://www.matrixscience.com)查询,获得差异蛋白质点鉴定结果(表2)。10个蛋白质依据功能的不同可划分为3个类群:3个差异蛋白点与光合碳同化相关,分别为核酮糖二磷酸羧化酶大亚基(spot2)、核酮糖二磷酸羧化酶/加氧酶大链(spot4)、磷酸甘油酸激酶(spot9);电子传递与光合磷酸化相关蛋白1个,为细胞色素b6/f复合体铁硫蛋白(spot3);抗性抗衰老相关差异蛋白点6个,分别为过氧化物酶(spot1)、锰超氧化物歧化酶(spot5)、伴侣蛋白21前体(spot6)、过氧还蛋白过氧化物酶(spot8)和抗坏血酸过氧化物酶(spot11,12)。
2.4 促芽肥对头季稻灌浆后期叶片蛋白质点表达丰度的影响
2.4.1 光合碳同化相关蛋白 水稻籽粒的灌浆物质主要来源于花后的光合产物,光合产物供应不足是影响籽粒灌浆结实和限制水稻产量提高的重要因素,因此,水稻叶片在灌浆结实期间光合能力下降的快慢会严重影响其结实率及产量[3,18]。本研究中共鉴定到3个与光合碳同化相关的蛋白质(图2),由图2可知,在头季稻灌浆后期随着生育进程的进行,NA处理和NNA处理剑叶中核酮糖二磷酸羧化酶大亚基(spot2)、核酮糖二磷酸羧化酶/加氧酶大链(spot4)和磷酸甘油酸激酶(spot9)表达量均呈下降趋势,但施用促芽肥处理和未施用促芽肥处理3个蛋白的下调幅度不同,NA处理的核酮糖二磷酸羧化酶大亚基(图3-A)、核酮糖二磷酸羧化酶/加氧酶大链、磷酸甘油酸激酶表达量的下调幅度明显小于NNA处理。

表2 头季稻灌浆后期剑叶响应不同促芽肥处理的差异表达蛋白质鉴定结果Tab.2 Resu lts of differen tially exp ressed p roteins of flag leaf in response to different nitrogen app lication for bud developm ent at late grain filling stage in the first cropping rice
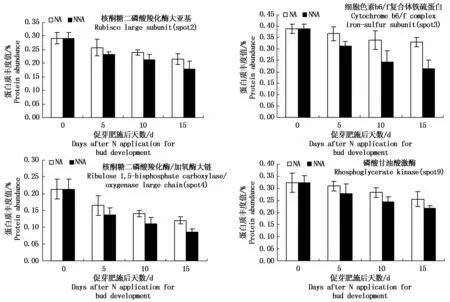
图2 光合作用相关差异蛋白在不同促芽肥处理下的表达丰度Fig.2 Effect of d ifferent nitrogen app lication for bud developm ent on expression of proteins involving photosynthesis of flag leaf at late grain filling stage in the first cropp ing rice

图3 相关蛋白表达变化图Fig.3 Exp ression changes of p roteins
2.4.2 电子传递与光合磷酸化相关蛋白 光合碳同化固定的CO2还需要利用光反应中产生的同化力ATP和NADPH才能完成CO2的还原,实现光合作用的贮能过程;本研究中鉴定到的细胞色素b6/f复合体铁硫蛋白(spot3)与同化力ATP和NADPH的合成有关(图2),由图2可知,无论是NA处理还是NNA处理,细胞色素b6/f复合体铁硫蛋白表达量在头季稻灌浆后期随灌浆的进行均呈下降趋势,但下降幅度NA处理明显小于NNA处理。
2.4.3 抗性抗衰老相关蛋白 水稻叶片在进入灌浆后期时衰老开始加速,本试验中鉴定到6个与抗性抗衰老相关的蛋白,由图4可知,在头季稻灌浆后期,NNA处理的过氧化物酶(spot1)、锰超氧化物歧化酶(spot5)、伴侣蛋白21前体(spot6)、过氧还蛋白过氧化物酶(spot8)(图3-B)和抗坏血酸过氧化物酶(spot11,12)6个蛋白表达量均随生育进程呈下调趋势;NA处理的过氧化物酶(spot1)、锰超氧化物歧化酶(spot5)、过氧还蛋白过氧化物酶(spot8)和抗坏血酸过氧化物酶(spot11,12)5个蛋白表达量在施用促芽肥后呈先上升后缓慢下降趋势,而伴侣蛋白21前体(spot6)蛋白表达量自促芽肥施用后至头季稻成熟则一直呈上升趋势;在促芽肥施用后至头季稻成熟的各个时期,NA处理的6个与抗性抗衰老相关的蛋白表达量均较同一时期的NNA处理有明显的上调。

图4 抗性抗衰老相关差异蛋白在不同促芽肥处理下的表达丰度Fig.4 Effect of different nitrogen app lication for bud developm ent on exp ression of p roteins involving resistance of flag leaf at late grain filling stage in the first cropping rice
2.5 促芽肥对头季稻灌浆后期叶片叶绿素SPAD值以及光合速率的影响
不同促芽肥处理对头季稻灌浆后期光合作用相关蛋白的表达有显著影响,光合代谢相关蛋白的降解在NA处理下得到明显减缓,进而影响了叶片的SPAD值以及光合速率。由图5可看出,施用促芽肥后,2个处理的剑叶叶绿素含量变化趋势不同,NA处理的剑叶叶绿素含量呈现先小幅上升再缓慢下降,在整个头季稻灌浆后期保持着较高的叶绿素含量水平;NNA处理的剑叶叶绿素含量在头季稻灌浆后期则呈较大幅度下降,因而,NA处理的剑叶叶绿素含量在施用促芽肥后5,10,15 d分别比NNA处理的高出8.01%,15.53%,23.57%,差异达显著水平,且越接近成熟期两处理间的差异越明显。
从图5还可看出,施用促芽肥后,净光合速率的变化趋势2个处理基本相同,呈逐步下降趋势,但下降幅度2个处理不同,NA处理的剑叶净光合速率下降幅度明显小,施用促芽肥后5,10,15 d分别比NNA处理的高出11.09%,23.68%,22.55%,差异达显著水平,可见,施用促芽肥并不能改变头季稻灌浆后期剑叶衰老、光合能力下降的趋势,但能明显减缓其下降幅度,使头季稻灌浆后期剑叶能维持较高的叶绿素含量和光合速率,具有较强的源供应能力,为增加头季稻籽粒灌浆的物质供应和提高头季稻结实率奠定物质基础。
2.6 促芽肥对头季稻灌浆后期叶片细胞保护酶、可溶性蛋白含量及MDA含量的影响
不同促芽肥处理对头季稻灌浆后期叶片的抗逆性有显著影响,表3显示,施用促芽肥后,头季稻灌浆后期NA处理剑叶的SOD活性、POD活性、CAT活性呈现先上升再缓慢下降的趋势,MDA含量呈现先小幅上升后明显下降、至促芽肥施后15 d又上升的趋势;NNA处理的剑叶中SOD活性、POD活性和CAT活性在头季稻灌浆后期则呈逐渐下降趋势,MDA含量呈逐步上升趋势,因而,在施用促芽肥后至头季稻成熟的各个时期,NA处理的剑叶SOD活性、POD活性和CAT活性均显著高于NNA处理,与上述抗性抗衰老相关蛋白的变化相吻合,表明施用促芽肥确能提高头季稻灌浆后期叶片抵抗逆境和适应不良环境的能力,抑制膜脂的过氧化作用,从而有效地延缓头季稻灌浆后期功能叶的衰老进程。

图5 不同促芽肥水平下头季稻灌浆后期叶片的叶绿素含量和净光合速率Fig.5 Chlorophyll contents and net photosynthetic rate of flag leaf at late grain filling stage in the first cropping rice under different nitrogen app lications for bud developm ent

表3 不同促芽肥水平下头季稻灌浆后期叶片的细胞保护酶活性及M DA含量Tab.3 Cell p rotective enzym es activities and p roline content of flag leaf at late grain filling stage in the first cropping rice under different nitrogen app lication for bud developm en t
3 讨论与结论
植物叶片衰老是叶片逐渐丧失光合作用并进入细胞死亡的一个复杂的叶片发育过程[19],许多研究表明,叶绿素含量和衰老之间存在明显的负相关[20],叶片衰老期间,叶绿体的大小、数目均减少[21],整体光合作用水平下降[22];1,5-二磷酸核酮糖羧化酶/加氧酶(Rubisco)是光合作用中卡尔文循环里催化第一个主要的碳固定反应的酶,是CO2固定的关键酶,决定了光合作用的净效率,高等植物的1,5-二磷酸核酮糖羧化酶/加氧酶是由8个大亚基和8个小亚基组成的寡聚体,每个大亚基上都有催化CO2与1,5-二磷酸核酮糖结合的位点,因而其含量与CO2的固定密切相关,陆巍[23]和张荣铣等[24]研究认为,水稻叶片光合功能衰退时,1,5-二磷酸核酮糖羧化酶/加氧酶活性的下降速度明显快于光合电子传递活性,从而使光合碳循环和能量供需平衡遭到破坏,最终导致叶片丧失光合功能,因而,提高1,5-二磷酸核酮糖羧化酶/加氧酶的羧化效率对延缓水稻叶片光合功能衰退具有重要意义;Ding等[25]研究认为氮素能够调节细胞分裂素(CTK)的代谢,而CTK能够诱导1,5-二磷酸核酮糖羧化酶/加氧酶的合成[26]。羧化阶段产生的3-磷酸甘油酸,是一种有机酸,还需要磷酸甘油酸激酶(PGAK)和甘油醛磷酸脱氢酶利用光反应中产生的同化力ATP和NADPH,使3-磷酸甘油醛转变成磷酸丙糖,才能实现CO2的还原和光合贮能过程;本试验中发现在头季稻灌浆后期,叶片中的核酮糖二磷酸羧化酶大亚基、核酮糖二磷酸羧化酶/加氧酶大链、磷酸甘油酸激酶表达量逐渐下调,但这3个蛋白表达量在促芽肥施用后的下调幅度明显小于未施用促芽肥的,核酮糖二磷酸羧化酶大亚基、核酮糖二磷酸羧化酶/加氧酶大链是1,5-二磷酸核酮糖羧化酶/加氧酶的重要组成部分,1,5-二磷酸核酮糖羧化酶/加氧酶和PGAK都是卡尔文循环的关键酶,卡尔文循环将CO2转化为糖类,并运输到穗部供给籽粒灌浆,这其中1,5-二磷酸核酮糖羧化酶/加氧酶参与CO2的羧化,PGAK参与CO2的还原,可见,施用促芽肥通过提高头季稻灌浆后期稻株的氮素水平,有效地延缓了卡尔文循环中1,5-二磷酸核酮糖羧化酶/加氧酶和PGAK 2个关键酶的降解,使卡尔文循环在头季稻灌浆后期能顺利的运转,保证了头季稻灌浆后期能有较充足的光合同化物质供应。
卡尔文循环中CO2的还原还需要利用光反应中产生的同化力ATP和NADPH,鉴定到的细胞色素b6/f复合体铁硫蛋白参与光合作用原初反应过程,与同化力ATP和NADPH的合成有关,含有Cyt f、Cyt b6(2个,电子传递循环剂)和(Fe-S)R,在光合电子传递及能量转换过程中扮演着重要角色,它一方面作为中间电子载体连接PSⅠ与PSⅡ2个光系统,介导PSⅠ与PSⅡ之间的线性电子传递和围绕PSⅠ的循环电子传递;另一方面作为能量转换器利用电子传递过程中释放出来的电子自由能将质子从类囊体膜外侧(基质侧)跨膜转运到类囊体膜内侧(囊腔侧),形成跨膜质子电化学梯度,为ATP合酶催化合成ATP提供能量[27-28]。此外,它还参与调节LHCⅡ激酶的活化,控制激发能在2个光系统之间的分配及NADPH与ATP的比例,使植物在环境的变化中维持尽可能高的光能转换效率[29]。在水稻生育后期,随着叶片的衰老,产生的同化力NADPH和ATP也逐步下降,致使羧化阶段固定的CO2无法完全转化为磷酸丙糖,而此时是杂交水稻弱势粒灌浆的重要时期,CO2同化能力的下降速度直接影响弱势粒的灌浆。本试验发现,施用促芽肥能相对提高细胞色素b6/f复合体铁硫蛋白在头季稻灌浆后期叶片中的表达,意味着促芽肥能够减缓头季稻灌浆后期光合电子传递活性和光合同化能力的下降速度,保证了头季稻灌浆后期叶片具有较高的光合同化能力。
对头季稻灌浆后期叶片叶绿素SPAD值以及光合速率的考察发现,叶片叶绿素SPAD值以及光合速率随着灌浆的进行总体呈逐渐下降趋势,但在NA处理中,叶片叶绿素SPAD值以及光合速率的下降趋势明显比NNA处理小,与上述光合相关蛋白表达的趋势一致,证明了蛋白鉴定结果的准确性。上述结果说明施用的促芽肥确能有效地延缓头季稻灌浆后期叶片卡尔文循环中CO2羧化和还原的关键酶1,5-二磷酸核酮糖羧化酶/加氧酶(Rubisco)和磷酸甘油酸激酶的降解,保证了头季稻灌浆后期卡尔文循环的运转,相对提高与光合同化能力相关蛋白细胞色素b6/f复合体铁硫蛋白在头季稻灌浆后期叶片中的表达,使该时期叶片仍具有较高的CO2光合同化能力,从而提高头季稻灌浆后期的干物质积累和源供应能力,为增加头季稻籽粒灌浆的物质供应和提高头季稻结实率奠定物质基础。
水稻进入生育后期,一方面稻株体内活性氧的产生在增加,另一方面活性氧的清除在减弱,使活性氧的平衡被打破,不能被及时清除,从而对细胞及组织造成损害,是导致叶片衰老加速的重要原因[30]。叶片衰老速度的快慢与活性氧清除系统存在极其密切的关系[31],锰超氧化物歧化酶、过氧还蛋白过氧化物酶、抗坏血酸过氧化物、过氧化物酶都是活性氧酶促清除系统的成员,对防御活性氧毒害、维持细胞膜结构和功能具有重要作用[32];本试验发现,在头季稻灌浆后期,NNA处理的锰超氧化物歧化酶、过氧还蛋白过氧化物酶、抗坏血酸过氧化物、过氧化物酶蛋白表达量均随生育进程呈下调趋势;NA处理的这5个蛋白表达量在施用促芽肥后呈先上升后缓慢下降趋势,在促芽肥施用后至头季稻成熟的各个时期,NA处理的这5个蛋白表达量均较同一时期的NNA处理有明显的上调。与2个处理的保护酶生理测定结果一致,证明了蛋白鉴定结果的准确性,说明在头季稻灌浆后期稻株体内活性氧大量增多导致叶片衰老加速的情况下,施用的促芽肥能够通过提高稻株体内与活性氧酶促清除系统相关酶蛋白的表达量来防御活性氧的毒害,从而提高了头季稻灌浆后期叶绿体的膜电阻和流动性,延缓了功能叶的衰老和光合作用能力的下降速度。
植物衰老的差误理论认为基因转录和翻译成蛋白质过程中的差误包括氨基酸排列顺序的错误或多肽链折叠的错误,累积达到一定程度时引起衰老;分子伴侣是一类在细胞中能识别正在合成的多肽或部分折叠的多肽并与多肽的一定部位相结合,帮助这些多肽折叠、组装或转运的蛋白质,不仅能帮助新生蛋白质进行正确的折叠和组装,而且在修复受伤害、变性蛋白质的过程中也有重要作用[33-35],分子伴侣含量与分子伴侣前体密切相关,本试验结果发现,施用促芽肥改变了头季稻灌浆后期剑叶中伴侣蛋白前体的表达量趋势,使其表达量从未施用促芽肥处理下的随生育进程逐渐下调趋势转变为逐渐上升趋势,说明施用促芽肥能显著减少头季稻灌浆后期叶片中蛋白质合成过程因多肽链折叠错误而引起的差误积累,还能加速被破坏无法复性蛋白质的降解,减少它们的聚集积累及同功能蛋白质间发生有害作用的可能性,从而延缓头季稻灌浆后期功能叶的衰老。综上研究结果可知,促芽肥对头季稻灌浆后期叶片蛋白质的表达有明显影响,头季稻后期剑叶中与光合碳同化、电子传递与光合磷酸化相关蛋白随灌浆进程呈下降趋势,施用促芽肥能明显减缓其下调幅度,从而使头季稻灌浆后期剑叶具有相对较高的叶绿素含量和净光合速率,提高了头季稻灌浆后期的干物质积累和源供应能力;还能明显提高头季稻灌浆后期剑叶中与抗性相关蛋白的表达量,从而增强稻株的活性氧清除能力,抑制功能叶的膜脂过氧化作用,延缓头季稻灌浆后期剑叶的衰老速率,这是高产再生稻头季稻结实率明显高于单季稻或双季稻的原因,研究结果在一定程度上丰富了再生稻高产栽培的促控机理,可为进一步的相关研究提供理论依据。
[1] 谢金水,邵彩虹,唐秀英,等.养分胁迫对籽粒灌浆期水稻叶片衰老影响的蛋白质组学分析[J].中国水稻科学,2011,25(2):143-149.
[2] 翟虎渠,曹树青,万建民,等.超高产杂交稻灌浆期光合功能与产量的关系[J].中国科学:C辑,2002,32(3):211-217.
[3] 徐芳芳,桑贤春,任德勇,等.水稻早衰突变体esl2的遗传分析及基因定位[J].作物学报,2012,38(8):1347-1353.
[4] Xie X J,Shen SH H,Li Y X,et al.Effect of photosynthetic characteristic and dry matter accumulation of rice under high temperature at heading stage[J].African Journal of Agricultural Research,2011,6(7):1931-1940.
[5] Kim JW,Shon JY,Lee C K,et al.Relationship between grain filling duration and leaf senescence of temperate rice under high temperature[J].Field Crops Research,2011,122(3):207-213.
[6] 李木英,郑 伟,石庆华,等.不同杂交稻灌浆期叶片衰老特性及其对水分亏缺的响应[J].中国农学通报,2011,27(3):44-45.
[7] 王丹英,章秀福,邵国胜,等.不同叶色水稻叶片的衰老及对光强的响应[J].中国水稻科学,2008,22(1):77-81.
[8] 魏海燕,张洪程,马 群,等.不同氮肥吸收利用效率水稻基因型叶片衰老特性[J].作物学报,2010,36(4):645-654.
[9] 陈鸿飞,张志兴,林文雄.促芽肥对水稻再生芽萌发生长过程蛋白质表达的影响[J].中国生态农业学报,2014,22(12):1405-1413.
[10] 马 均,王化新,孙晓辉,等.促芽肥15N在再生稻中的分配及其作用研究[J].西南农业学报,1992,5(1):41-46.
[11] 刘保国,王光明,陈 静,等.促芽肥影响水稻再生芽萌发的生理系列化基础[J].西南农业学报:水稻栽培专辑,1998,11:41-47.
[12] 黄育民,李义珍,蔡亚港,等.再生稻丰产技术研究Ⅷ报-再生稻株对促芽肥的吸收积累[J].福建稻麦科技,1995,13(3):45-47.
[13] 张志兴,陈 军,李 忠,等.水稻籽粒灌浆过程中蛋白质表达特性及其对氮肥运筹的响应[J].生态学报,2012,32(10):3209-3224.
[14] 邵彩虹,唐秀英,李明心,等.6-苄基腺嘌呤延缓水稻衰亡效应的蛋白质组学分析[J].华北农学报,2014,29(1):14-19.
[15] 张志良.植物生理学实验指导[M].北京:高等教育出版社,1995:155-163.
[16] 邵彩虹,王 萍,陈 金,等.养分胁迫对不易早衰类型水稻生育后期叶片影响的蛋白质组学分析[J].华北农学报,2015,30(4):51-58.
[17] 夏其昌,曾 嵘.蛋白质化学与蛋白质组学[M].北京:科学出版社,2004:278.
[18] 段 俊,梁承邺,黄毓文.杂交水稻开花结实期间叶片衰老[J].植物生理学报,1997,23(2):139-144.
[19] 张艳军,赵江哲,张可伟.植物激素在叶片衰老中的作用机制研究进展[J].植物生理学报,2014,50(9):1305-1309.
[20] 杨淑慎,高俊凤,李学俊.高等植物叶片的衰老[J].西北植物学报,2001,21(6):1271-1277.
[21] Hashimoto H,Kura-Hotta M,Katoh S,et al.Changes in protein content and in the structure and number of chloroplasts during leaf senescence in rice seedling[J]. Plant Cell Physiol,1989,30(5):707-715.
[22] 王复标,黄福灯,程方民,等.水稻生育后期叶片早衰突变体的光合特性与叶绿体超微结构观察[J].作物学报,2012,38(5):871-879.
[23] 陆 巍.水稻光合功能衰退与Rubisco结构与功能变化的研究[D].南京:南京农业大学,2000.
[24] 张荣铣,戴新宾,许晓明,等.叶片光合功能期与作物光合生产潜力[J].南京师大学报:自然科学版,1999,22(3):376-386.
[25] Ding C,You J,Wang S,et al.A proteomic approach to analyze nitrogen and cytokinin responsive proteins in rice roots[J].Mol Biol Rep,2012,39:1617-1626.
[26] Ookawa T,Naruoka Y,Sayama A,et al.Cytokinin effects on Ribulose-1,5-bisphosphate carboxylase/oxygenase and nitrogen partitioning in rice during ripening[J]. Crop Sci,2004,44:2107-2115.
[27] Cramer W A,Zhnag H,Yan J,et al.Evolution of Photosynthesis:Time-independent structure of the cytochrome b6/f comp lex[J].Biochemistry,2004,43:5921-5928.
[28] Hope A B.The chloroplasts cytochrome b6 f complex:a critical of focus on function[J].Biochim Biophys Acta,1993,1143:1-22.
[29] 候 程.高等植物菠菜光合膜蛋白Cyt b6 f纯化及其结晶条件初探[D].石家庄:河北科技大学,2012.
[30] Gill S S,Tuteja N.Reactive oxygen species and antioxidantmachinery in abiotic stress tolerance in crop plants[J].Plant Physiol Biochem,2010,48(12):909-930.
[31] 李 健,赵 宇,李锦锦,等.不同基因型玉米叶片衰老与活性氧代谢的关系及其调控[J].华北农学报,2011,26(1):131-135.
[32] 史 静,潘根兴,夏运生,等.镉胁迫对两品种水稻生长及抗氧化酶系统的影响[J].生态环境学报,2013,22(5):832-837.
[33] 彭春瑞,邵彩虹,潘晓华,等.水稻育秧肥的壮秧效应及其蛋白质组学分析[J].中国水稻科学,2012,26(1):27-33.
[34] 杜晓华,黄伶俐,刘会超.植物热激蛋白70(HSP70)研究进展[J].河南科技学院学报,2014,42(3):1-5.
[35] Hartl F U,Hayer-Hartl M.Molecular chaperones in the cytosol:from nascent chain to folded protein[J].Science,2002,295:1852-1858.
Effects of Nitrogen App lication for Bud Developm ent on Protein Expression of Flag Leaf at Late G rain Filling Stage in the First Cropping Rice
CHEN Hongfei1,2,3,ZHANG Zhixing1,2,3,LINWenxiong1,2,3(1.School of Life Sciences,Fujian Agricultural and Forestry University,Fuzhou 350002,China;2.Institute of Agricultural Ecology,Fujian Agricultural and Forestry University,Fuzhou 350002,China;3.Key Laboratory of Crop Ecology and Molecular Physiology of Fujian Universities,Fujian Agricultural and Forestry University,Fuzhou 350002,China)
By using comparative proteom ics combined with bioinformatics and related physiological indicators,this paper studied the regulating effects of nitrogen application for bud development on protein expression of flag leaf at late grain filling stage in the first cropping rice withⅡYou 1273 as a material.Two treatments of urea levels:300.00,0.00 kg/ha applying at18 d after heading stage were included in the experiment in the first cropping rice. A total of 15 differentially expressed proteins were detected in different fertilizer treatments by the 2D maps of flag leaf proteome.And these proteins were grouped into three expression patterns according to their functions in photosynthesis carbon assim ilation,electron-transfer and photophosphorylation,the defense response to stresses etc..Our results indicated that the proteins involving photosynthesis were down-regulated at late grain filling stage in the first cropping rice.But,nitrogen app lication for bud development could significantly slow down the decreased amplitude of photosynthesis-related proteins in expression abundance,which relatively enhanced the chlorophyll content and photosynthetic rate of functional leaf and increased the source supply ability at late grain filling stage in the first cropping rice.The result also showed that nitrogen application for bud development could significantly increase the expression abundance of resistance-related protein at late grain filling stage in the first cropping rice,which helpedenhance the scavenging of active oxygen,retard the peroxidation ofmembrane lipid and delay the senescence rate of flag leaf.As a result,the seed-setting rate and yield of the first cropping rice were significantly increased under NA treatment.
Ratooning rice;Leaf;Nitrogen application for bud development;Proteom ics;Grain-yield
S511.03 文献标识码:A 文章编号:1000-7091(2016)03-0127-08
10.7668/hbnxb.2016.03.019
2016-03-10
国家自然科学基金项目(31401306);福建省生态学重点学科项目(6112C0611);福建农林大学科技发展资金项目(KF2015039)
陈鸿飞(1977-),男,福建莆田人,实验师,博士,主要从事作物生理与分子生态学研究。
林文雄(1957-),男,福建莆田人,教授,博士,博士生导师,主要从事作物生理与分子生态学研究。
