脂多糖对体外培养猪颗粒细胞增殖、凋亡和雌二醇分泌的影响
2016-07-11果双双施振旦马卫明
果双双,李 辉,施振旦,马卫明*
(1.山东农业大学动物科技学院,泰安 271018; 2.江苏省农业科学院畜牧研究所,南京 210014)
脂多糖对体外培养猪颗粒细胞增殖、凋亡和雌二醇分泌的影响
果双双1,李辉2,施振旦2,马卫明1*
(1.山东农业大学动物科技学院,泰安 271018; 2.江苏省农业科学院畜牧研究所,南京 210014)
摘要:旨在探讨脂多糖(LPS)对猪颗粒细胞增殖、凋亡及雌二醇(E2)分泌的影响。采用不同浓度的LPS(0、500、1 000和2 000 ng·mL-1)处理体外培养的猪颗粒细胞,并测定细胞增殖、凋亡及相关基因FN1、IGF2、IGFBP2、CyclinD1、CyclinD2和P27kip的表达,同时对E2的分泌及P450arom的表达进行检测。结果表明,1 000或2 000 ng·mL-1LPS均可显著促进颗粒细胞增殖、抑制细胞凋亡、提高活细胞百分率(P<0.05)。而500 ng·mL-1LPS即可促进与细胞生长增殖、周期相关基因FN1(P<0.01),IGF2、IGFBP2(P<0.05),CyclinD1、CyclinD2(P<0.01)的表达,抑制阻碍细胞周期的基因P27kip(P<0.01)的表达,同时可以通过强烈抑制芳香化酶P450arom基因的表达(P<0.01),显著抑制E2的分泌(P<0.01)。综上表明,LPS可以促进颗粒细胞增殖并抑制细胞凋亡,同时抑制颗粒细胞分泌E2的功能。
关键词:颗粒细胞;LPS;细胞增殖;细胞凋亡;基因表达;E2
内毒素化学本质是脂多糖(Lipopolysaccharide,LPS),是革兰阴性菌细胞壁的组成成分,也是医学上引起机体“发烧”的热源。LPS在细菌正常生活状态时并不释放,仅在细菌自溶或人工方法裂解菌体后才释放出来[1-2]。LPS在动物体内普遍存在,具有多种来源途径,如环境、饲料、消化道内细菌解体、患病、创伤、交配以及分娩过程等[3-5]。大多数内毒素感染在初期并无明显症状,随着时间延长,逐渐表现出不良反应[6-7]。LPS在生殖方面造成的不良影响主要表现为卵巢活动受到抑制、类固醇激素分泌能力下降、胚胎发育受阻、黄体机能降低、抑制胚胎着床和引发流产等[8-9],最终导致动物繁殖性能下降。在奶牛研究中,LPS可以通过影响动物下丘脑-垂体促性腺激素的释放而造成内分泌紊乱,最终抑制LH/FSH分泌的脉冲波并能够推迟或完全抑制排卵前高峰的出现[10-11],阻碍E2分泌并抑制排卵。LPS对动物繁殖活动的影响,也表现在卵泡发育的方面。如具有正常周期性卵巢活动的健康牛卵泡液中LPS含量很低,平均为(0.06±0.04)ng·mL-1,而患有子宫内膜炎、缺乏周期性卵巢活动的牛卵泡液中LPS含量很高,平均为(176.1±112) ng·mL-1,最高可达875.2 ng·mL-1[12]。
卵泡颗粒细胞在卵泡发育和排卵过程中起着极其重要的作用,其通过微绒毛与卵母细胞发生内分泌联系,并对后者提供营养支持。颗粒细胞通过不断增殖、分泌类固醇激素、分泌细胞外基质蛋白、血管生成因子、免疫和趋化因子,促进卵泡的生长、发育和成熟,并通过影响垂体促性腺激素的分泌进行卵泡的募集、选择和成熟排卵等[13]。有研究显示,当LPS作用于牛颗粒细胞时,能激活TLR-4并上调一系列炎症因子如IL-1β、IL-6和TNFα等的表达,同时抑制颗粒细胞E2的分泌[14-16]。LPS对动物繁殖性能的负面影响,除通过下丘脑和垂体的间接作用降低促性腺激素分泌之外[17],是否直接影响卵泡颗粒细胞增殖及其功能?为此本研究采用LPS直接处理离体培养的猪颗粒细胞的方法,通过测定细胞增殖、凋亡、相关基因表达及激素分泌水平等来阐明。
1材料与方法
1.1试验材料
选取15~20头180日龄左右的健康商品母猪,采集卵巢置于37 ℃生理盐水中,2 h内带回实验室。
1.2猪颗粒细胞的原代培养
用无菌生理盐水冲洗卵巢2~3次,灭菌纱布拭干卵巢表面,选取直径>2 mm的卵泡,用10 mL注射器配10号针头抽取卵泡液(注意避开血管)。将收集的卵泡液置于37 ℃水浴中静置5~10 min,收集上清离心,将收集到的细胞用PBS(Gibco)重悬两次。用DMEM/F12培养液(Gibco)重悬细胞,台盼蓝染色计数后调整细胞密度至1×106个·mL-1,添加含10%胎牛血清FBS(Gibco),1%青链霉素(MP)的DMEM/F12培养基并置于37 ℃、5% CO2条件下培养。培养24 h后用PBS轻轻洗去未贴壁细胞,更换新鲜的含有10%FBS,1%青链霉素,猪垂体来源FSH(1 ng·mL-1,Sigma),雄烯二酮(0.1 μmol·L-1,Sigma)且含有不同浓度LPS(0、500、1 000和2 000 ng·mL-1,Extracted fromE.coli055:B5,sigma)的新鲜培养液对细胞进行刺激。E2测定组,在细胞培养24 h换液时,更换为含有2%FBS的上述培养液。
1.3细胞增殖检测
LPS处理细胞48 h,利用CCK8试剂盒(上海翊圣)处理细胞[18-19],在培养箱内孵育4 h后,利用全波长酶标仪测定在450 nm处的吸光度。其吸光度数值与细胞增殖成正比。
1.4细胞凋亡检测
LPS处理细胞48 h,用不含EDTA的胰酶消化并收集细胞,用FITC Annexin V细胞凋亡检测试剂盒(BD Pharmingen)处理细胞,避光孵育15 min后,通过分析型流式细胞仪检测细胞凋亡。
1.5基因相对表达量检测
在LPS处理细胞24及48 h时,分别利用细胞总RNA提取试剂盒(北京天根)提取细胞总RNA,并利用反转录试剂盒(TaKaRa)将其反转录为cDNA,利用ABI7500进行qRT-PCR(试剂购自Transgen Biotech)检测细胞生长增殖相关(FN1、IGF2、IGFBP2),增殖周期相关(CyclinD1、CyclinD2、P27kip)以及芳香化酶P450arom基因的相对表达量。qRT-PCR相关引物序列见表1。
1.6E2测定
LPS处理细胞48 h后回收培养液,用PBS溶液1∶5进行稀释,利用E2定量测定试剂盒(北京北方生物),采用酶联免疫竞争法检测细胞培养液中E2含量,经显色后在酶标仪测定吸光值(OD值),通过拟合浓度—吸光度曲线,计算出待测细胞培养液中E2含量。
表1qRT-PCR相关引物
Table 1Primers used in the real-time quantitative PCR assay
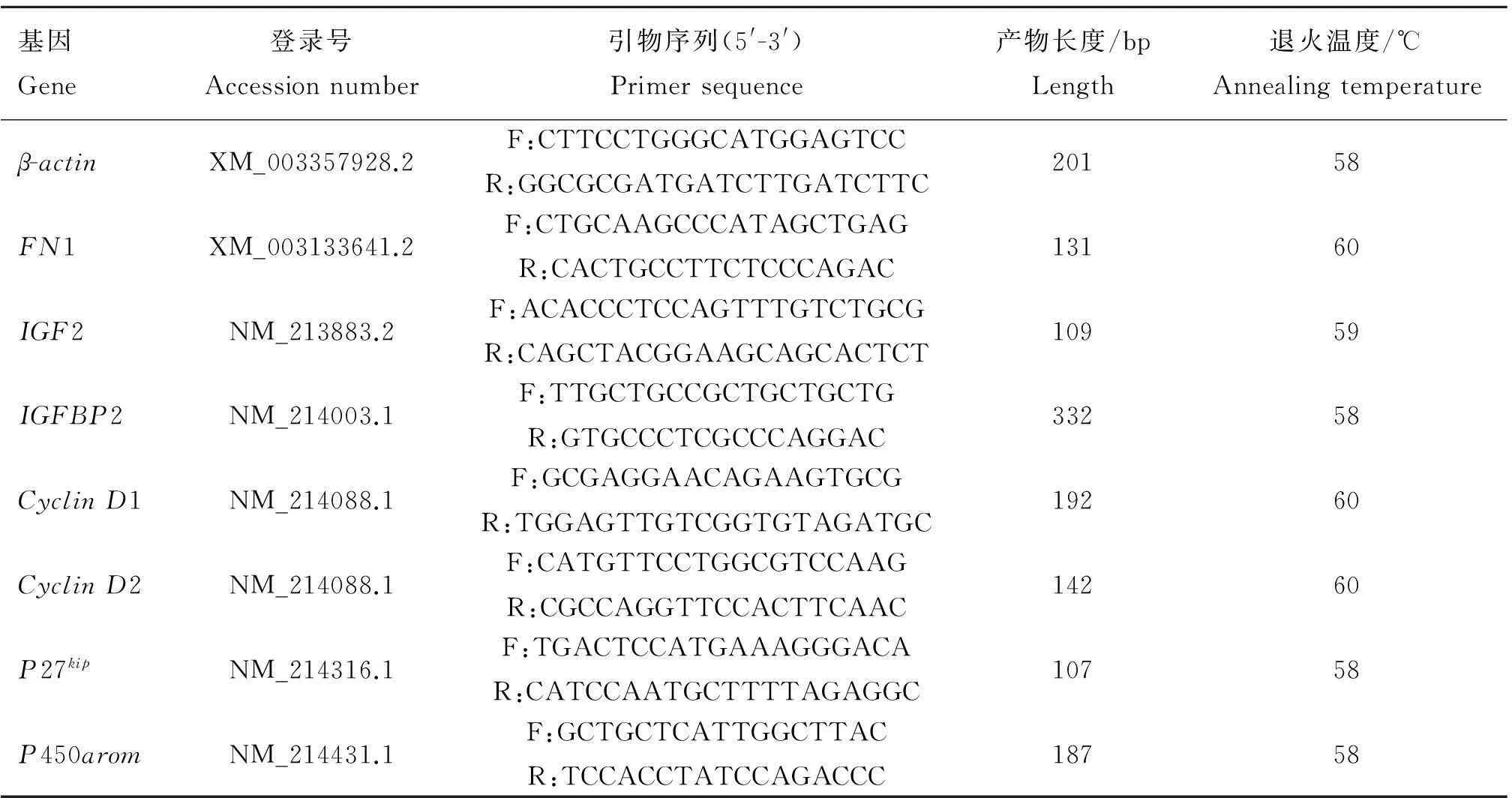
基因Gene登录号Accessionnumber引物序列(5'-3')Primersequence产物长度/bpLength退火温度/℃Annealingtemperatureβ-actinXM_003357928.2F:CTTCCTGGGCATGGAGTCCR:GGCGCGATGATCTTGATCTTC20158FN1XM_003133641.2F:CTGCAAGCCCATAGCTGAGR:CACTGCCTTCTCCCAGAC13160IGF2NM_213883.2F:ACACCCTCCAGTTTGTCTGCGR:CAGCTACGGAAGCAGCACTCT10959IGFBP2NM_214003.1F:TTGCTGCCGCTGCTGCTGR:GTGCCCTCGCCCAGGAC33258CyclinD1NM_214088.1F:GCGAGGAACAGAAGTGCGR:TGGAGTTGTCGGTGTAGATGC19260CyclinD2NM_214088.1F:CATGTTCCTGGCGTCCAAGR:CGCCAGGTTCCACTTCAAC14260P27kipNM_214316.1F:TGACTCCATGAAAGGGACAR:CATCCAATGCTTTTAGAGGC10758P450aromNM_214431.1F:GCTGCTCATTGGCTTACR:TCCACCTATCCAGACCC18758
1.7统计分析
qRT-PCR统计结果以0添加组为对照,运用2-△△Ct的方法计算各检测基因的相对表达量,用内参β-actin进行标准化处理。数据处理采用SPSS Statistics 17.0的单因素方差分析,不同处理间的显著性检验采用Duncan式多重比较。采用Origin 8.6作图。结果采用“平均值±标准误”表示,P<0.05表示差异显著,P<0.01表示差异极显著。
2结果
2.1LPS 对颗粒细胞增殖的影响
不同浓度LPS处理细胞48 h对细胞增殖产生的影响。由图1可以看出,随着LPS浓度的升高,反映细胞增殖活力的CCK8 OD值呈现出剂量依赖性上升趋势,尤其当添加浓度达到1 000和2 000 ng·mL-1时,细胞增殖显著高于对照组及500 ng·mL-1处理组(P<0.01)。
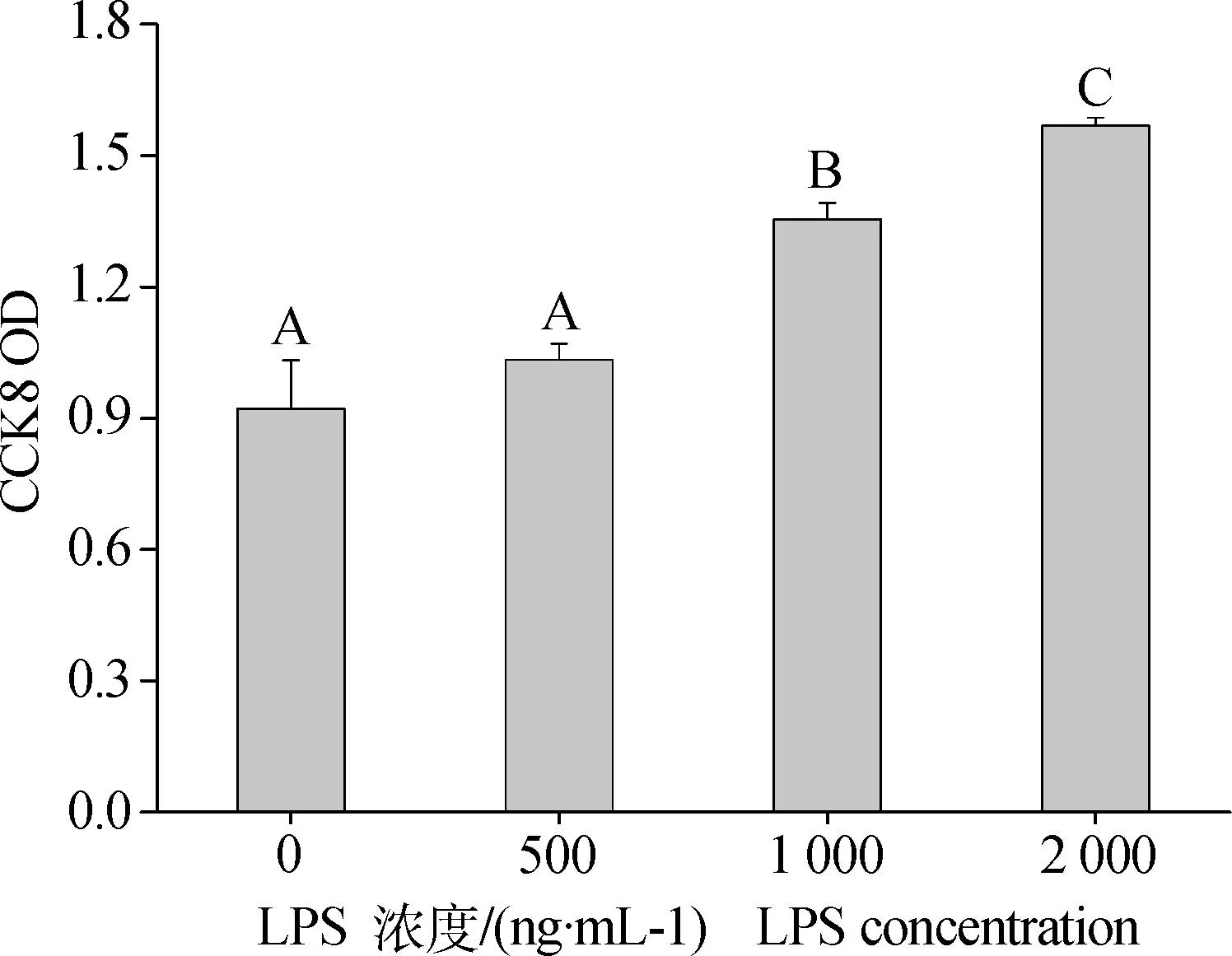
(n=3,P<0.01)图1 不同浓度LPS对颗粒细胞增殖的影响Fig.1 Dose-effect of LPS on Granulosa cells proliferation
2.2LPS对颗粒细胞凋亡的影响
通过分析型流式细胞仪检测细胞凋亡发现,LPS可以一定程度地抑制细胞凋亡,且随浓度的逐渐升高,凋亡细胞明显减少。图2显示,经LPS处理48 h后,流式细胞仪检测的颗粒细胞中活细胞和凋亡细胞情况。图2A~D显示,随LPS添加量逐渐升高,凋亡细胞逐渐减少(右上和右下两个象限),活细胞数逐渐增多(左下象限)。图2E,F结果表明,添加1 000和2 000 ng·mL-1LPS浓度处理组的活细胞率显著高于对照组及LPS浓度为500 ng·mL-1处理组(P<0.05),添加1 000和2 000 ng·mL-1LPS浓度处理组间差异不显著。LPS添加对培养的猪颗粒细胞的总凋亡细胞百分率影响则恰恰相反。LPS添加浓度为1 000和2 000 ng·mL-1时,总凋亡细胞百分率处理组明显低于对照组及LPS浓度为500 ng·mL-1处理组(P<0.05)。
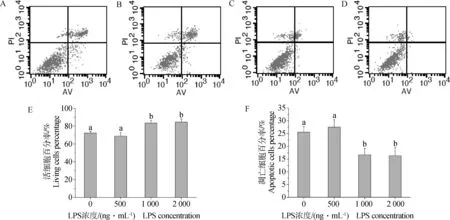
A.空白对照组;B.500 ng·mL-1LPS;C. 1 000 ng·mL-1LPS;D.2 000 ng·mL-1LPS;E.活细胞百分率;F.凋亡细胞百分率(n=3)。不同小写字母表示差异显著。下同A.Blank control group;B.500 ng·mL-1LPS;C.1 000 ng·mL-1LPS;D.2 000 ng·mL-1LPS;E.Percentage of living cells;F.Percentage of apoptotic cells(n=3).Different small letters show significant difference (P<0.05).The same as below图2 不同浓度LPS对颗粒细胞凋亡的影响Fig.2 Dose-effect of LPS on granulosa cells apoptosis
2.3LPS对颗粒细胞增殖及周期相关基因表达的影响
图3A显示,LPS处理细胞24 h,与增殖相关的基因FN1、IGF2和IGFBP2,与周期相关的基因CyclinD1、CyclinD2和P27kip的相对表达量。图3B显示,LPS处理细胞48 h各基因的相对表达量。由图3可知,LPS处理细胞24 及48 h能够显著上调促进细胞增殖的基因FN1(P<0.01,图3A;P<0.05,图3B)和IGF2(P<0.01)的相对表达量,且IGF2基因相对表达量呈现剂量依赖性上调。LPS处理24 h显著(P<0.05,图3A)促进IGFBP2的基因相对表达量,48 h差异不显著(图3B)。
图3A显示,促进细胞周期的基因CyclinD1,CyclinD2在LPS处理24 h时,基因相对表达量差异不显著,而图3B显示LPS处理48 h时两基因相对表达量极显著上调(P<0.01)。抑制细胞周期的基因P27kip在LPS处理24和48 h时都出现了极显著下调(P<0.01)。
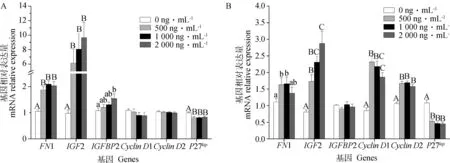
A.LPS处理24 h时各基因相对表达量;B.LPS处理48 h时各基因相对表达量。不同大写字母表示差异极显著。下同A.Relative expression ratio of genes treated by LPS for 24 h;B.Relative expression ratio of genes treated by LPS for 48 h.Different capital letters show extremely significant difference (P<0.01).The same as below图3 不同浓度LPS对颗粒细胞增殖及周期相关基因表达的影响(n=9)Fig.3 Dose-effect of LPS on granulosa cells proliferation and cells cycle genes expression (n=9)
2.4LPS对颗粒细胞E2分泌和芳香化酶表达的影响
LPS处理颗粒细胞48 h后,测定培养液中E2结果见图4A,LPS各处理组颗粒细胞E2分泌量极显著低于对照组(P<0.01),各处理组间差异不显著。与E2分泌相关的芳香化酶P450arom基因相对表达量测定结果见图4B。经LPS处理后,各处理组中P450arom基因相对表达量受到抑制,与对照组差异极显著(P<0.01)。
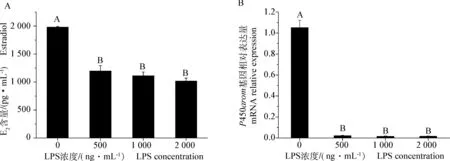
A.颗粒细胞E2分泌量(n=3);B.P450arom基因表达量 (n=9)A.E2secretion of Granulosa cells(n=3);B.Relative expression ratio of P450arom gene (n=9)图4 不同浓度LPS对颗粒细胞E2分泌量和P450arom基因表达量的影响Fig.4 Dose-effect of LPS on Granulosa cells estradiol secretion and P450arom gene expression
3讨论
在畜牧业生产中,各种原因造成的微生物过度繁殖,使大量LPS进入动物体内,导致多种疾病发生,严重损害动物的生产繁殖性能,带来不可估量的经济损失[20-21]。本研究根据LPS对颗粒细胞增殖及E2分泌的直接影响,通过不同的试验手段,发现LPS处理离体猪颗粒细胞后,一定程度地促进细胞增殖及抑制细胞凋亡。促进细胞增殖的基因FN1、IGF2、IGFBP2及促进细胞分裂周期的基因CyclinD1、CyclinD2的相对表达量呈现显著上调,抑制细胞周期的基因P27kip显著下调。以上基因的相对表达量变化说明添加LPS对细胞增殖及周期的基因表达产生影响,从而引起细胞的增殖及凋亡。有研究报道显示,LPS能够促进小胶质细胞[22]、卵巢细胞[23]、小肠上皮细胞[24]等多种细胞的增殖。也有试验表明,LPS能够促进小鼠的睾丸支持细胞增殖[25-26]。在本研究中,LPS能够显著地促进猪颗粒细胞的增殖,抑制细胞的凋亡,推测可能是LPS通过模式识别受体TLR-4途径[7,27]激活下游NF-κB通路导致的。有相关文献证实,LPS能够经由TLR-4途径促进大鼠血管平滑肌细胞[28]和肝癌细胞[29]等增殖。以及激活下游NF-κB通路导致细胞呈现增殖,这在Hela细胞研究中有所报道[30],NF-κB也直接参与调控细胞的增殖凋亡基因的表达等[31-32]。NF-κB通路能够促进一系列抗炎因子及肿瘤坏死因子的表达[16],其中,由TNFα介导的细胞凋亡途径被激活,而LPS能够通过激活NF-κB途径抑制TNFα导致的细胞凋亡现象[33-34],已经在肝细胞[35]、角质细胞[36]、内皮细胞[37]等多种细胞中被证实。本研究通过CCK8和流式细胞仪的检测结果也证实LPS具有促进猪颗粒细胞增殖和抑制凋亡的作用,且与增殖及增殖周期相关基因相对表达量的结果相吻合。
LPS在影响颗粒细胞的正常增殖凋亡程序时,也强烈地抑制颗粒细胞分泌E2的功能。P450arom由CyP19A1基因编码,是雄激素向雌激素转化的关键酶,在颗粒细胞分泌E2中起重要作用。本研究发现LPS能够直接抑制其基因表达,导致E2分泌减少。此结果与K.Onnureddy[14]及T.Shimizu[15]等在牛颗粒细胞上得到的结果相似,证明LPS确实可以直接抑制颗粒细胞分泌E2的功能。而E2的正常分泌被抑制将导致动物生殖内分泌的紊乱,继发一系列不良反应。这一结果也解释了因疾病或因环境问题而引起的LPS感染所造成的安静发情、断续发情、卵泡发育受阻、排卵数下降、产后乏情、产仔数下降等现象。
4结论
LPS通过颗粒细胞的分子调控及基因表达影响其正常的增殖、凋亡,同时LPS可以直接抑制颗粒细胞分泌雌二醇。
参考文献(References):
[1]李桂杰,朱瑞良,郭宝聚.细菌内毒素的研究进展[J].山东农业大学学报,1999,30(2):93-98.
LI G J,ZHU R L,GUO B J.Advances on the study of bacterial endotoxin[J].JournalofShandongAgriculturalUniversity, 1999,30(2):93-98.(in Chinese)
[2]郭萌,李冠民,黄清泉.细菌内毒素研究进展[J].中国实验动物学报,2009,17(5):397-400.
GUO M,LI G M,HUANG Q Q.Advances in research of bacterial endotoxin[J].ActaLaboratoriumAnimalisScientiaSinica, 2009,17(5):397-400.(in Chinese)
[3]MARCHESINI G,DE NARDI R,GIANESELLA M, et al. Effect of induced ruminal acidosis on blood variables in heifers[J].BMCVetRes,2013,9:98.
[4]GILBERT R O.The effects of endometritis on the establishment of pregnancy in cattle[J].ReprodFertilDev,2011,24(1):252-257.
[5]江丹莉,张鑫杰,潘远基,等.水面载鹅密度对水体细菌生长、鹅血液内毒素含量、种鹅繁殖性能和雏鹅生长性能的影响[C].哈尔滨:第十四次全国家禽科学学术讨论会,2009:618-623.
JIANG D L,ZHANG X J,PAN Y J, et al.Effects of surface water load density on the growth of bacteria,the content of blood endotoxin,the breeding performance and the growth performance of geese[C].Harbin:Fourteenth National Symposium on Poultry Science,2009:618-623.(in Chinese)
[6]BILODEAU-BOURGEOIS L,BOSWORTH B G,PETERSON B C.Differences in mortality,growth,lysozyme,and toll-like receptor gene expression among genetic groups of catfish exposed to virulent Edwardsiella ictaluri[J].FishShellfishImmunol,2008,24(1):82-89.
[7]BEUTLER B.Inferences,questions and possibilities in Toll-like receptor signalling[J].Nature,2004,430(6996):257-263.
[8]施振旦,李孝伟,田允波,等.奶牛繁殖杀手——内毒素[J].中国奶牛,2007(3):29-32.
SHI Z D,LI X W,TIAN Y B,et al.The killer of dairy cattle breeding----endotoxin[J].ChinaDairyCattle,2007(3):29-32.(in Chinese)
[9]郭文军,施振旦,梅承,等.提高奶牛受胎率的卵泡和黄体活动调控研究[J].中国奶牛,2006(6):7-11.
GUO W J,SHI Z D,MEI C,et al.Study on regulating ovarian and luteal activities in improving conception rate in holstein cows[J].ChinaDairyCattle, 2006(6):7-11.(in Chinese)
[10]SUZUKI C,YOSHIOKA K,IWAMURA S,et al.Endotoxin induces delayed ovulation following endocrine aberration during the proestrous phase in Holstein heifers[J].DomestAnimEndocrinol,2001,20(4):267-278.
[11]BATTAGLIA D F,KRASA H B,PADMANABHAN V,et al.Endocrine alterations that underlie endotoxin-induced disruption of the follicular phase in ewes[J].BiolReprod,2000,62(1):45-53.
[12]HERATH S,WILLIAMS E J,LILLY S T, et al.Ovarian follicular cells have innate immune capabilities that modulate their endocrine function[J].Reproduction,2007,134(5):683-693.
[13]MCNATTY K P,HEATH D A,HENDERSON K M,et al.Some aspects of thecal and granulosa cell function during follicular development in the bovine ovary[J].JReprodFertil, 1984,72(1):39-53.
[14]ONNUREDDY K,RAVINDE R,ONTERU S K, et al.IGF-1 attenuates LPS induced pro-inflammatory cytokines expression in buffalo (Bubalusbubalis) granulosa cells[J].MolImmunol,2015,64(1):136-143.[15]SHIMIZU T,MIYAUCHI K,SHIRASUNA K, et al. Effects of lipopolysaccharide (LPS) and peptidoglycan (PGN) on estradiol production in bovine granulosa cells from small and large follicles[J].Toxicolinvitro,2012,26(7):1134-1142.
[16]YENUGANTI V R,RAVINDER R,SINGH D.Conjugated linoleic acids attenuate LPS-induced pro-inflammatory gene expression by inhibiting the NF-kappaB translocation through PPARgamma in buffalo granulosa cells[J].AmJReprodImmunol,2014,72(3):296-304.
[17]BATTAGLIA D F,BEAVER A B,HARRIS T G,et al.Endotoxin disrupts the estradiol-induced luteinizing hormone surge:interference with estradiol signal reading,not surge release[J].Endocrinology,1999,140(6):2471-2479.
[18]WANG J,CHEN X,SU L, et al. MicroRNA-126 inhibits cell proliferation in gastric cancer by targeting LAT-1[J].BiomedPharmacother,2015,72:66-73.
[19]LIU W,KONG H,ZENG X,et al.Iptakalim inhibits PDGF-BB-induced human airway smooth muscle cells proliferation and migration[J].ExpCellRes,2015,336(2):204-210.
[20]JIANG D L,LIU L,WANG C L,et al.Raising on water stocking density reduces geese reproductive performancesviawater bacteria and lipopolysaccharide contaminations in “geese-fish” production system[J].AgrSciChina,2011,10(9):1459-1466.
[21]施振旦,刘丽,孙爱东.“禽-鱼”高密度养殖对水禽生产性能危害的研究[J].中国家禽,2011,33(13):1-3.SHI Z D,LIU L,SUN A D.Study on ‘bird--fish’ high density cultivation on production performance of waterfowl harm[J].ChinaPoultry,2011,33(13):1-3.(in Chinese)
[22]张思思,蔡江晖,万敬员,等.羟基积雪草苷对LPS诱导小胶质细胞增殖的抑制作用及机制研究[J].中国病理生理杂志,2015,31(3):428-434.
ZHANG S S,CAI J H,WANG J Y,et al.Inhibitory effect of madecassoside on LPS-stimulated microglia[J].ChineseJournalofPathophysiology,2015,31(3):428-434.(in Chinese)
[23]CHAKRAVORTTY D,KATO Y,SUGIYAMA T, et al.Inhibition of p38 mitogen-activated protein kinase augments lipopolysaccharide-induced cell proliferation in CD14-expressing Chinese hamster ovary cells[J].InfectImmun,2001,69(2):931-936.
[24]OLAYA J,NEOPIKHANOV V,URIBE A.Lipopolysaccharide ofEscherichiacoli,polyamines,and acetic acid stimulate cell proliferation in intestinal epithelial cells[J].invitroCellDevBiolAnim,1999,35(1):43-48.
[25]PETERSEN C,FRÖYSA B,SÖDER O.Endotoxin and proinflammatory cytokines modulate Sertoli cell proliferationinvitro[J].JReprodImmunol,2004,61(1):13-30.
[26]PETERSEN C,SVECHNIKOV K,FROYSA B,et al.The p38 MAPK pathway mediates interleukin-1-induced Sertoli cell proliferation[J].Cytokine,2005,32(1):51-59.
[27]TAKEUCHI O,AKIRA S.Pattern recognition receptors and inflammation[J].Cell,2010,140(6):805-820.[28]YANG G,ZHOU X,CHEN T, et al.Hydroxysafflor yellow A inhibits lipopolysaccharide-induced proliferation and migration of vascular smooth muscle cells via Toll-like receptor-4 pathway[J].IntJClinExpMed,2015,8(4):5295-5302.
[29]WANG L,ZHU R,HUANG Z, et al.Lipopolysaccharide-induced toll-like receptor 4 signaling in cancer cells promotes cell survival and proliferation in hepatocellular carcinoma[J].DigDisSci,2013,58(8):2223-2236.
[30]HAO H,XU F,HAO J,et al.Lipoxin A4 suppresses lipopolysaccharide-induced hela cell proliferation and migration via NF-κB pathway[J].Inflammation, 2015,38(1):400-408.
[32]CHANG C F,ZHAO W M,MEI J X,et al. Branches of NF-κb signaling pathway regulate hepatocyte proliferation in rat liver regeneration[J].GenetMolRes,2015,14(3):7643-7654.
[33]MANNA S K,AGGARWAL B B.Lipopolysaccharide inhibits TNF-induced apoptosis:role of nuclear factor-kappaB activation and reactive oxygen intermediates[J].JImmunol,1999,162(3):1510-1518.
[34]AGGARWAL B B.Apoptosis and nuclear factor-kappa B:a tale of association and dissociation[J].BiochemPharmacol,2000,60(8):1033-1039.
[35]XU Y,BIALIK S,JONES B E,et al.NF-κB inactivation converts a hepatocyte cell line TNF-α response from proliferation to apoptosis[J].AmJPhysiol, 1998,275(4 Pt 1):C1058-C1066.
[36]QIN J Z,CHATURVEDI V,DENNING M F,et al.Role of NF-kappaB in the apoptotic-resistant phenotype of keratinocytes[J].JBiolChem,1999,274(53):37957-37964.
[37]ZEN K,KARSAN A,STEMPIEN-OTERO A,et al.NF-kB activation is required for human endothelial survival during exposure to tumor necrosis factor-α but not to interleukin-1β or lipopolysaccharide[J].JBiolChem,1999,274(40):28808-28815.
(编辑程金华)
Effects of Lipopolysaccharide on Proliferation,Apoptosis and Estradiol Secretion ofinvitroCultured Porcine Granulosa Cells
GUO Shuang-shuang1,LI Hui2,SHI Zhen-dan2,MA Wei-ming1*
(1.CollegeofAnimalScienceandVeterinaryMedicine,ShandongAgriculturalUniversity,Tai’an271018,China;2.InstituteofAnimalScience,JiangsuAcademyofAgriculturalSciences,Nanjing210014,China)
Abstract:In order to study the effects of Lipopolysaccharide(LPS) on proliferation,apoptosis and estradiol (E2) secretion of porcine granulosa cells.Primary porcine granulosa cells were culturedinvitroand treated by LPS at different concentrations (0,500,1 000 and 2 000 ng·mL-1).Then cell proliferation,apoptosis,secretion of E2and related gene expressions,FN1,IGF2,IGFBP2,CyclinD1,CyclinD2,P27kipandP450aromwere determined individually.The results showed that cell proliferation and survival ratio were significantly promoted at LPS concentrations of 1 000 or 2 000 ng·mL-1(P<0.05),on the contrary,cell apoptosis was inhibited.However,gene expression levels ofFN1 (P<0.01),IGF2,IGFBP2 (P<0.05),CyclinD1 andCyclinD2 (P<0.01) were up-regulated at 500 ng·mL-1of LPS,P27kip(P<0.01) which hindered cell cycle was inhibited.P450aromgene expression was decreased,resulting the inhibition of E2secretion when treated by LPS.These results suggested that granulosa cell proliferation could be promoted by LPS,while cell apoptosis and E2secretion were inhibited.
Key words:granulosa cells;LPS;cell proliferation;cell apoptosis;gene expression;E2
doi:10.11843/j.issn.0366-6964.2016.04.008
收稿日期:2015-09-25
基金项目:江苏省自然科学基金(BK20151365);国家自然科学基金(31402080);江苏省农业科学院基本科研业务专项[ZX(15)4031]
作者简介:果双双(1989-),女,山东济南人,硕士生,主要从事动物繁殖内分泌学研究, E-mail: qicaitianping7777@163.com *通信作者:马卫明,副教授,E-mail: mawm@sdau.edu.cn
中图分类号:S828;S814.1
文献标志码:A
文章编号:0366-6964(2016)04-0693-07
