嗜酸性氧化亚铁硫杆菌亚铁氧化酶基因研究
2016-06-18张鹏华徐玲玲
张鹏华, 李 江, 徐玲玲
东华理工大学, 省部共建核资源与环境教育部重点实验室, 南昌 330013
嗜酸性氧化亚铁硫杆菌亚铁氧化酶基因研究
张鹏华,李江*,徐玲玲
东华理工大学, 省部共建核资源与环境教育部重点实验室, 南昌 330013
摘要:嗜酸性氧化亚铁硫杆菌(Acidithiobacillus ferrooxidans)是一种重要的浸矿微生物,以亚铁离子或还原态硫化物为能源物质。亚铁氧化酶在其亚铁离子氧化过程中起着极其重要的作用。对来源于不同矿区的4株菌(YHC、721、737和745)进行了研究,16S rDNA序列比对结果显示4株菌与A. ferrooxidans的同源性在98%以上,确定该4株菌均为Acidithiobacillus属的不同株系。亚铁氧化能力测定结果表明YHC菌和737矿菌相近,721矿菌与745矿菌相近,YHC菌和737矿菌的亚铁氧化能力明显强于721矿菌和745矿菌。通过PCR扩增得到4种菌亚铁氧化酶基因序列,将基因序列与GenBank 中的参考序列进行比对分析,共3个碱基具有差异,但大部分为同义密码子,仅第73位氨基酸有不同,YHC菌和737矿菌亚铁氧化酶基因第73位氨基酸是Val,721矿菌和745矿菌则是Ala, Val突变成Ala使得热稳定性下降,导致半衰期缩短,所以酶的持续作用时间会减少,进而影响酶活。
关键词:嗜酸性氧化亚铁硫杆菌;亚铁氧化酶基因;热稳定性
嗜酸性氧化亚铁硫杆菌(Acidithiobacillusferrooxidans)是一种革兰氏阴性菌,具有化能自养、好氧、嗜酸、适于中温环境等特性,广泛存在于酸性矿山水及含铁或硫的酸性环境中,以氧化亚铁离子或还原态硫化物为能源物质,是一种重要的矿物浸出微生物[1,2]。亚铁氧化能力对A.ferrooxidans的生长、代谢周期、环境适应能力起着非常重要的作用,氧化亚铁酶是A.ferrooxidans菌氧化亚铁离子的关键酶[3]。为了使A.ferrooxidans菌更好的服务于工业生产, 还必须进一步了解该细菌的遗传、生理生化等特性, 形成优良菌种选育的科学理论和方法。为此,一些学者对A.ferrooxidans菌的铁硫代谢系统进行了研究,如其中的铜兰蛋白[4],但对于和铜兰蛋白一样重要的亚铁氧化酶的研究报道却比较少。目前, 人们通过对亚铁氧化系统中的多种功能组分的分离纯化和分子生物学方面的研究, 对亚铁氧化系统中的关键酶及其分子生物学作用途径有了比较深入的了解, 认为亚铁氧化后大部分的电子顺电势梯度传递给O2, 同时少量电子逆电势梯度传递, 产生还原力NAD(P)H 参与CO2的固定和有氧代谢[5,6],张成桂等[7]认为亚铁氧化酶是A.ferrooxidans菌亚铁氧化电子传递链上最初的电子受体。尽管其在进化过程中非常保守,但不同地域的A.ferrooxidans菌不论是在表现型还是在基因型上都与标准菌株(SS3)有差异[8]。
本文通过取自4处不同地域铀矿的嗜酸性氧化亚铁硫杆菌的菌株,对其进行了亚铁氧化活性的研究及16S rDNA序列分析,进一步从不同菌株中克隆得到亚铁氧化酶基因(Iro),并对其进行氨基酸序列比对,以期揭示不同种A.ferrooxidans菌亚铁氧化酶基因的亚铁氧化能力及其关键氨基酸的初步分析。
1材料与方法
1.1实验材料
1.1.1供试菌株(721来源于江西乐安某铀矿,745矿菌来源广东韶关某铀矿,YHC菌和737矿菌来源于新疆某铀矿)。
1.1.2试剂总DNA提取试剂盒Fast DNA spin kit for soil购自MP Biomedical公司,离心柱型琼脂糖凝胶DNA回收试剂盒DP209购自北京天根生物科技有限公司,dNTP、Taq酶、250 bp DNA Ladder均购自TaKaRa公司,GoldView TM核酸染料购自BM BioMed公司,琼脂糖、乙二胺四乙酸(EDTA)、TBE电泳缓冲液等均为国产或进口试剂,PCR引物由生工生物工程(上海)有限公司合成。
1.1.39K培养基配方A液(g/L):(NH4)2SO43.0 g, K2HPO40.5 g,KCl 0.1 g,MgSO4·7H2O 0.5 g, Ca(NO3)20.01 g;去离子水800 mL,H2SO4调节pH至2.0, 121℃高压灭菌20 min。 B液:FeSO4·7H2O 44.78 g,去离子水200 mL, H2SO4调节pH至2.0,孔径为0.22 μm的滤膜过滤除菌。将灭菌后的A液与B液混匀后分装[9]。
1.2实验方法
1.2.1亚铁氧化能力的测定分别按90 mL的9K+10 mL上一代菌液的比例接菌培养[10],根据A.ferrooxidans菌的生长特性及生长状况(培养过程中颜色的变化)选取6个时间点,运用乙二胺四乙酸(EDTA)滴定法[10]测定培养液中的亚铁离子的含量。
1.2.2细菌基因组提取对纯化后培养的菌株进行菌体收集,13 000 r/min离心收集菌体,用pH 1.8的无菌9K清洗菌体,TE洗到中性[11]。使用Fast DNA spin kit for soil基因组DNA抽提试剂盒提取基因组DNA 。1.2.3菌株的16S rDNA基因序列测定16S rDNA扩增选用以下通用引物:1387R:5′-AGAGTTTGATCTGGCTAG-3′;27F:5′-GGGCGGGTGTACA-AGGC-3′。以总DNA模板进行16S rDNA PCR扩增,总体积为25 μL PCR扩增反应体系为:18.3 μL的双蒸水,2.5 μL 10×PCR Buffer,2.0 μL dNTP(2.5 mmol/L),0.5 μL引物1387R和27F(20 μmol/L),0.2 μL rTaq酶,1 μL模板DNA。PCR反应程序:95℃预热处理5 min;94℃变性45 s,53℃退火45 s,72℃延伸90 s,30个循环;72℃延伸10 min。PCR产物验证后离心柱型琼脂糖凝胶DNA回收试剂盒DP209切胶回收,回收产物送生工生物工程(上海)股份有限公司测序。
1.2.4亚铁氧化酶基因(Iro)序列测定根据NCBI数据库中A.ferrooxidans菌株(SS3)的亚铁氧化酶基因序列设计如下Iro引物:IroF:5′-ATG-AGTGAGAAACTGAAGGAAACCGAAAATTCC-3′;IroR:5′-GTATTGTATAGCGTTTACGCCGATGCCC-GCATAA-3′。以总DNA为模板对Iro进行PCR扩增,总体积为25 μL PCR扩增反应体系为:18.3 μL的双蒸水,2.5 μL 10×PCR Buffer,2.0 μL dNTP(2.5 mmol/L),0.5 μL引物IroF和IroR(20 μmol/L),0.2 μL rTaq酶,1 μL模板DNA。PCR反应程序:95℃预热处理5 min;94℃变性45 s,50℃退火45 s,72℃延伸90 s,30个循环;72℃延伸10 min。PCR产物验证后离心柱型琼脂糖凝胶DNA 回收试剂盒DP209切胶回收。构建pEASY-T3克隆重组质粒:将目的基因与pEASY-T3 Cloning Vector以4∶1的比例混合配成5 μL体系,室温连接20 min。转化感受态细胞大肠杆菌Trans1,挑菌斑培养并且进行菌落PCR验证,菌落PCR反应体系(20 μL):10 μL 2×TaqPCR Master Mix,1 μL引物M13F和M13R,8 μL双蒸水。PCR反应程序同Iro基因扩增程序。验证正确后送生工生物工程(上海)股份有限公司测序,与NCBI数据库及自身相互比对分析。
1.2.5序列分析及三维结构预测使用NCBI上的ORF Finder网站预测Iro基因的开放阅读框,使用ClustalX进行多序列比对分析,并利用 ESPript(http://espript.ibcp.fr/ESPript/cgi-bin/ESPript.cgi)对多重比对序列着色。蛋白三维结构的预测利用I-TASSER方法进行(http://zhanglab.ccmb.med.umich.edu/I-TASSER/)。利用Protparam Tool进行蛋白质理化性质分析。
2结果与分析
2.14种菌亚铁氧化能力比较
A.ferrooxidans菌亚铁氧化能力通过亚铁离子消耗速率比较分析得出:前20 h 4种菌的亚铁氧化氧化能力相似,而20 h后亚铁氧化速率表现出较明显的差异,YHC和737矿菌的亚铁离子消耗速率接近,721矿菌和745矿菌的亚铁离子消耗速率接近,到80 h亚铁离子均被完全氧化(图1)。结果表明其氧化能力与4种菌所处地理位置相吻合,且YHC菌和737矿菌的亚铁离子消耗速率大于721矿菌和745矿菌。58 h时737与YHC已将亚铁消耗完全,而721与745矿菌要培养超过80 h才完全消耗亚铁离子。
2.216S rDNA基因序列分析及系统进化树构建
测序结果在NCBI上Blast比对,得到的测序结果表明4种菌与A.ferrooxidansstrain CK 16S的同源性都在98%以上,说明4种菌属于同一种的不同亚种。构建16S rDNA系统发育树,结果显示(图2),本实验研究的4株菌在进化上比较接近,进一步分析发现721与YHC之间遗传距离较近,737与745遗传距离较近。
2.3亚铁氧化酶(Iro)基因克隆
根据基因组序列设计Iro基因的特异引物,通过PCR技术从4种菌的基因组DNA中扩增得到亚铁氧化酶基因(Iro),纯化产物琼脂糖凝胶电泳图谱如图3,条带清晰单一,片段大小符合预期,结果表明4种菌均存在特异的Iro基因。
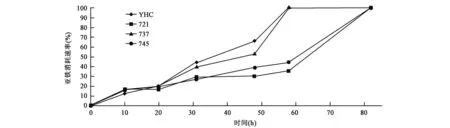
图1 亚铁氧化速率Fig.1 Ferrous oxidation rate.
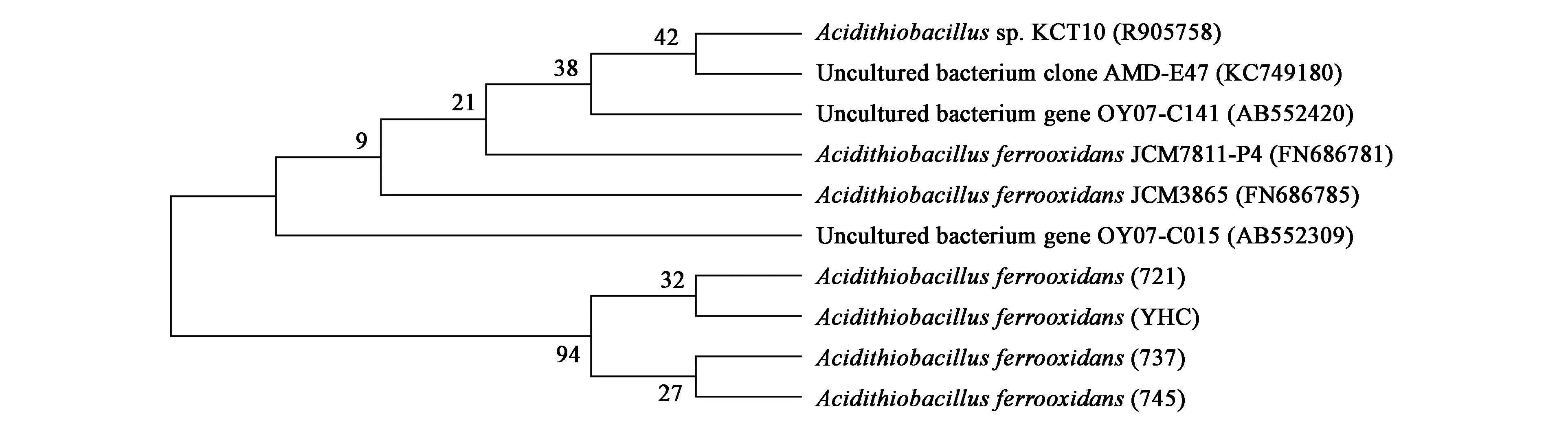
图2 A. ferrooxidans部分16S rDNA序列构建的系统发育树Fig.2 Phylogenetic tree of partial 16S rDNA from A. ferrooxidans.
2.4亚铁氧化酶基因(Iro)进化树构建及氨基酸序列比对
4种菌Iro基因核酸序列相互比对,结果显示亚铁氧化酶基因编码区在721、737、745、YHC中核酸长度为315 bp,编码104个氨基酸。构建Iro系统发育树,结果显示(图4),与其亚铁氧化能力
相一致,721与745较近,737与YHC较近。本文主要是研究A.ferrooxidans菌亚铁氧化酶基因多态性,特别是编码区差异位点与菌株亚铁氧化活性之间的关系,实验结果显示,上述4种菌Iro基因在第172、218、265号碱基位置变化发生,但只有第218位碱基的突变引起了第73位氨基酸编码发生改变,721、745菌的第218位碱基是C,而737、YHC菌的第218位碱基是T,相对应的是721、745菌的第73位氨基酸为丙氨酸(Ala),737、YHC的第73位氨基酸为缬氨酸(Val)。通过多种亚铁氧化酶氨基酸序列比对分析(图5)后发现Iro基因N端数十个氨基酸序列差别较大,属易突变区域,从第20 位开始氨基酸序列变得十分保守,从第73位氨基酸开始的C端有少量差异。
2.5亚铁氧化酶家族生物信息学分析
将获得的亚铁氧化酶基因全长序列以FASTA格式输入到NCBI的ORF Finder,Blastx比对结果显示该基因编码104个氨基酸。由于该蛋白没有合适的晶体结果做模板,因此使用I-Tasser对该结构进行从头预测[12~14],模拟其三维结构如图 6(彩图见图版三)所示,该预测的三维结构由两个小α螺旋和无规则卷曲组成,结构上有一定的新颖性,值得进行晶体结构解析。利用Protparam Tool分析4种A.ferrooxidans菌Iro序列,推导其分子式分别为C906H1501N315O376S81、C909H1505N315O378S79、C906H1499N315O376S81和C960H1589N333O398S85。

图4 A. ferrooxidans Iro系统发育树Fig.4 Phylogenetic tree of Iro from A. ferrooxidans.

图5 亚铁氧化酶氨基酸序列比对Fig.5 Amino acid sequence alignment of ferrous oxidase.注:α代表α螺旋,TT代表β转角,β代表β折叠。
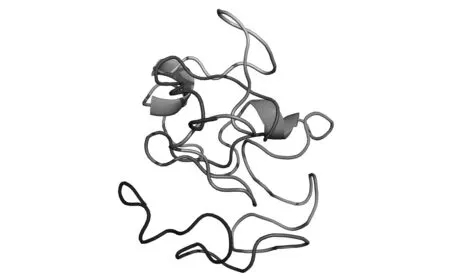
图6 亚铁氧化酶三维结构预测模型Fig.6 Three dimensional structure prediction model of ferrous oxidase.(彩图见图版三)
3讨论
在本实验中A.ferrooxidans菌通过氧化亚铁离子获得能量,故可根据亚铁离子的消耗速率判断A.ferrooxidans菌的生长速率,而A.ferrooxidans菌的生长速率与其亚铁氧化能力成正比[8],本研究通过A.ferrooxidans菌消耗亚铁离子的定量检测来比较本文4种菌的亚铁氧化酶的活性。根据实验结果可以推断721、745矿菌亚铁氧化酶的活性强于YHC和737矿菌。
根据测序及比对结果,A.ferrooxidans菌氧化酶基因第172、265位碱基发生突变,但没有引起相应氨基酸的改变,属于同义密码子,这类突变可能反映了菌种的地域多态性。第218位碱基改变(胞嘧啶变为胸腺嘧啶),导致第73位氨基酸由721、745矿菌亚铁氧化酶中的丙氨酸(Ala)变为YHC和737矿菌的缬氨酸(Val)。据Tomonobu等[15]推断亚铁氧化酶属于高还原氧化铁硫蛋白家族(HiPIP家族)中的周质蛋白,HiPIP家族蛋白是一种非常普遍的电子传递载体,具有很高的氧化还原电势,前37个氨基酸属于信号肽序列,信号肽影响亚铁氧化酶的跨膜转运效率,进而影响该菌的亚铁氧化效率[16],本研究多序列比对结果也证实亚铁氧化酶基因前37个氨基酸序列属于易突变序列,这为今后的突变研究提供了一定的参考。根据预测的蛋白质三维结构模型,第73位氨基酸位于蛋白质表面,亚铁氧化酶是由8~10个分子量为6 kDa的亚基组成的同聚体,缬氨酸(Val)在与其他氨基酸作用时产生的范德华力强于丙氨酸(Ala), 缬氨酸(Val)突变成丙氨酸(Ala)使得热稳定性下降, 热稳定性下降必然导致半衰期缩短,所以酶的持续作用时间会减少,推测第73位丙氨酸(Ala)变为缬氨酸(Val)会使作为A.ferrooxidans菌亚铁氧化电子传递链上最初的电子受体的亚铁氧化酶活性升高。以上是编码区的差异,研究表明非编码区域的上游区域,经比对发现所有菌株的序列都与参考序列相同,说明这个区域是严格保守的。 而在编码区的下游区域,比对结果表明差异位点比较多,该区域在转录、翻译、蛋白质调控等方面有调节作用,具体情况还需进一步研究[8]。
由于4种菌亚铁氧化酶基因的差异极小,结合4种菌明显的亚铁氧化能力差异表现及前人的研究,推断亚铁氧化酶中第73位的丙氨酸变为缬氨酸和其亚铁氧化能力提高正相关,下一步还需通过突变研究对其进行功能验证。本研究也进一步佐证了Iro基因的N端氨基酸和酶活密切相关。
总的来看,A.ferrooxidans菌的亚铁氧化酶基因非常保守,是其能量代谢系统中的关键酶基因,但其存在的差异也表明不同来源的A.ferrooxidans菌氧化亚铁酶基因具有分子遗传多样性。本文的结果能为以后的分子生物学研究及菌种的选育提供参考。
参考文献
[1]Nestor D, Valdivia U, Chaves A P. Mechanism of bioleaching of a refractory mineral of gold withThiobacillusferrooxidans[J].Int. J. Mineral Proc.,2001,62:187-198.
[2]Sacdowski Z E,Jazdzyk H K. Bioleaching of copper oreflotation concentrates[J].Minerals Engin.,2003,16:51-53.
[3]欧阳建华,陈新华.氧化亚铁硫杆菌亚铁氧化系统的研究进展[J].生物技术通报,2009,03:002.
[4]Bengrine A, Cuiliani N, Appia A C,etal.. Sequence and expression of the rusticyanin structural gene fromThiobacilusferrooxidansATCC33020 stain[J].Biochim. Biophys. Acta,1998,1443:99-112.
[5]Elbehti A,Brasseur G,Lemesle M D.First evidence for existence of an uphill electron transfer through the bc1 and NADH Q oxidoreductase complexes of the acidophilic obligate chemolithotrophic ferrous ion oxidizing bacteriumT.ferrooxidans[J]. J. Bacteriol.,2000,182:3602-3606.
[6]Rawlings D E. Characteristics and adaptability of iron and sulfur oxidizing microorganisms used for the recovery of metals from minerals and their concentrates [EB/OL].http: //www.microbialcellfactories.com/content/4/1/13.
[7]张成桂, 夏金兰, 邱冠周,等.嗜酸氧化亚铁硫杆菌亚铁氧化系统研究进展[J].中国有色金属学报,2006,7: 1239-1249.
[8]杨 宇,彭 宏,孙 斌,等.嗜酸氧化亚铁硫杆菌亚铁氧化酶基因分子多态性研究[J]. 遗传,2005.27(5):787-791.
[9]张在海. 铜硫化矿生物浸出高效菌种选育及浸出机理[D]. 长沙:中南大学,博士学位论文,2002.
[10]刘亚洁,李 江,牛建国,等.铀矿石生物浸出中氟对铁-硫氧化细菌的影响[J].有色金属,2006,4:18-21.
[11]郭 勤,韩文艳,李 江,等.氯离子对氧化亚铁硫杆菌和氧化亚铁钩端微螺菌活性的影响[J].有色金属:冶炼部分,2015,1: 42-45.
[12]Yang J, Yan R, Roy A,etal.. The I-TASSER Suite: Protein structure and function prediction[J]. Nat. Methods, 2015,12: 7-8.
[13]Roy A, Kucukural A, Zhang Y. I-TASSER: a unified platform for automated protein structure and function prediction[J]. Nat. Protocols, 2010, 5: 725-738.
[14]Zhang Y. I-TASSER server for protein 3D structure prediction[J]. BMC Bioinform., 2008,9: 40.
[15]Tomonobu K, Toshiyuki T,Kazuyuki S,etal.. Molecular cloning of the gene encodingThiobacillusferrooxidansFe(I1) oxidase[J]. J. Biol. Chem., 1992, 267(16):11242-11247.
[16]Williams K,Michael R C. Concepts of Genetics(Fifth Edition)[M].New Jersy:Prentice Hal,1997.
Study on Ferrous Oxidase-encoding Genes ofAcidithiobacillusferrooxidans
ZHANG Peng-hua, LI Jiang*, XU Ling-ling
LaboratoryofNuclearResourcesandEnvironment(ECIT),MinistryofEducation,EastChinaUniversityofTechnology,Nanchang330013,China
Abstract:Acidithiobacillus ferrooxidans is an important mineral-bioleaching bacterium with ferrous ion or reducible sulfide as the energy substance. Ferrous oxidase plays a key role in the ferrous ion oxidation process. In this study, four strains (YHC, 721, 737 and 745) were isolated from different mines showed more than 98% nucleotide sequence with the 16S rDNA of A. ferrooxidans. Ferrous oxidase activity assays indicated that strains YHC and 737 had similar activity, which were obviously higher than that of strains 721 and 745. Four ferrous oxidase-encoding genes from these Acidithiobacillus strains were amplified by PCR, sequenced and compared with reference sequences in GenBank. Three bases showed difference and accounted for the only divergence at position 73, Val for strains YHC and 737 and Ala for strains 721 and 745. When substituted Val with Ala by using the site-directed mutagenesis, the mutant enzymes showed decreased thermostability and shorter half-life. As results, the ferrous oxidase had reduced functional duration and less enzyme activity.
Key words:Acidithiobacillus ferrooxidans; ferrous oxidase gene; heat stability
收稿日期:2016-01-11; 接受日期:2016-03-03
基金项目:江西省教育厅科技计划项目(KJLD13048)资助。
作者简介:张鹏华,硕士研究生,研究方向微生物学。E-mail:1316154343@qq.com。*通信作者:李江,教授,研究方向为微生物冶金。E-mail:li66001@163.com
DOI:10.3969/j.issn.2095-2341.2016.03.10
