抑郁小鼠大脑GABA及谷氨酸囊泡转运蛋白表达变化*
2016-06-15宋靖方张小玲蔡弘扬常全忠
吴 琼,宋靖方,张小玲,蔡弘扬,常全忠
抑郁小鼠大脑GABA及谷氨酸囊泡转运蛋白表达变化*
吴琼1,宋靖方2,张小玲1,蔡弘扬1,常全忠3△
[摘要]目的研究抑郁症发病中GABA及谷氨酸囊泡转运蛋白表达的变化。方法小鼠随机分为对照组和造模组,使用社会失败模型对对照组小鼠造模,之后将其分为对抑郁症敏感组及对抑郁症不敏感组。将3组小鼠分别提取突触蛋白,并使用Western blot检测VGAT及VGLUT1表达丰度。结果对抑郁症敏感组小鼠与对照组小鼠相比,在接触区域的停留时间显著降低,且在角落区域的停留时间显著增加,差异有统计学意义(P<0.05);在前额叶皮层、海马中和对照组与对抑郁症不敏感组比较,对抑郁症敏感组VGLUT1及VGAT的表达水平显著增加,差异有统计学意义(P<0.05)。对抑郁症不敏感组与对照组相比其VGAT及VGLUT1水平的表达差异均无统计学意义(P>0.05)。在纹状体,虽然对抑郁症敏感组的小鼠VGAT及VGLUT1的表达水平均有增加(P<0.05),但是对抑郁症不敏感的小鼠这些蛋白表达也显著增加。结论在抑郁症发病中前额叶皮层及海马兴奋性与抑制性囊泡转运异常,这可能与抑郁症神经递质紊乱有关。
[关键词]抑郁症;谷氨酸囊泡转运蛋白;γ-氨基丁酸囊泡转运蛋白
抑郁症是目前最高发的精神疾病,它的发病以青壮年为主,并且自杀率高、诊断率低,为社会带来了沉重的负担;目前其发病机制尚不完全清楚,以往的研究多数集中在5-HT及胆碱能神经元的研究上[1-2]。近年来的研究认为,抑郁症发病中GABA能与谷氨酸能突触相关蛋白的表达发生变化,神经递质失调,突触可塑性也发生了改变[3-4];而对突触体中GABA及谷氨酸的表达尚少见研究。GABA囊泡转运体(VGAT)及谷氨酸囊泡转运体(VGLUT)是参与GABA能与谷氨酸能神经递质包装的重要囊泡蛋白,在GABA能与谷氨酸能神经递质释放中发挥重要作用[5]。本研究分别在前额叶皮层、海马、纹状体中提取突触体,并针对抑郁症发病后突触体中VGAT与VGLUT的水平变化进行探讨,为研究抑郁症发病中GABA能与谷氨酸能神经递质的变化提供了依据。
1材料与方法
1.1材料
1.1.1实验动物10周龄C57BL/6雄性小鼠30只,体质量22~25 g;24周龄CD1雄性退役种鼠22只。小鼠饲养于标准动物房,3~4只小鼠一笼,自由摄食、饮水。
1.1.2实验试剂VGAT单克隆抗体(抗鼠)、VGLUT单克隆抗体(抗鼠)购于Synaptic system公司,HRP-羊抗鼠二抗购于英韦创津公司,BCA试剂盒购于Thermo scientific,蛋白酶抑制剂购于罗氏公司。
1.2方法
1.2.1社会失败模型根据Nestler教授提供的方法[6],在该模型中,单笼饲养的被测小鼠与一只完全不熟悉的带有攻击性的CD1小鼠放在一起10 d,每天10 min攻击时间,其余时间用一个刻有漏洞的塑料盖板分开。对照小鼠与其相同品系但不熟悉的小鼠放在一起,每天接触10 min,后用刻有漏洞的塑料隔板分开。其行为学指标通过社会回避实验测定。社会回避实验的方法如下:首先将小鼠放入一50 cm×50 cm×20 cm的塑料盒中,盒子一侧中央放置一个透明有孔的有机玻璃盒子,高15 cm。有机玻璃盒周围划出接触区域,而对面角落的区域则划为角落区域。记录2.5 min内小鼠在接触区域的停留时间,记为T1,然后将小鼠放回原鼠笼休息1 min,用70%的乙醇擦拭塑料盒,以去除小鼠的气味。然后将CD1小鼠放于有机玻璃盒中,将实验小鼠再次放回盒内,记录2.5 min内小鼠在接触区域的停留时间,记为T2。以T2与T1的比值作为检测指标,当比值大于1时,将小鼠定为对该刺激不敏感的小鼠,反之当比值小于1时,定为对该刺激敏感的小鼠。
1.2.2突触小体的提取根据社会回避实验的结果,选取对照组、对抑郁症敏感组和对抑郁症不敏感组的小鼠,每组各4只。根据Duman教授提供的方法[7],被选取的小鼠分别断头取脑,在冰上快速分离前额叶皮层、海马、纹状体。取出的组织块放在溶液(0.32 mol/L蔗糖,20 mmol/L HEPES缓冲液,1 mmol/L EDTA,5 mmol/L 氟化钠,1 mmol/L 矾酸钠及0.1 g/L蛋白酶抑制剂)中研磨,其匀浆液在 4 ℃,2 800 r/min 转速下离心10 min。取其上清液再次离心10 min,转速为12 000 r/min。弃上清液,沉淀用溶液(50 mmol/L Tris-HCl,150 mmol/L 氯化钠,1% Triton X-100,0.1% SDS,2 mmol/L EDTA,1 mmol/L偏矾酸钠,5 mmol/L氟化钠及0.1 g/L蛋白酶抑制剂)溶解并超声。蛋白浓度通过BCA蛋白定量试剂盒检测。
1.2.3Western blot电泳将提取的突触小体的样品与上样缓冲液混合,放入沸水中加热10 min。取20 μg蛋白上样,通过10% SDS-PAGE胶电泳分离蛋白,电转至PVDF膜上。取脱脂奶粉封闭1 h,一抗(稀释比例1∶1 000)4 ℃过夜。次日,膜通过TBST溶液清洗3遍后,加入二抗孵育1 h,再次用TBST溶液清洗并转移至暗室内通过显影仪显影。蛋白丰度使用FluorChemTM软件分析测定。

2结果
2.1小鼠的社会回避行为小鼠被分为8只对照组小鼠和22只造模小鼠,通过社会失败模型,剔除死亡小鼠后,将造模组小鼠分为对抑郁症敏感组及对抑郁症不敏感组。在社会回避行为实验中,对抑郁症敏感组小鼠与对照组小鼠相比,在接触区域的停留时间显著降低且在角落区域的停留时间显著增加,差异有统计学意义(P<0.05),见图1。

A:小鼠分为对照组、敏感组和不敏感组。B、C:小鼠经过造模后,对抑郁症敏感组的小鼠与对照小鼠相比,当有CD1存在时在中间区域停留时间缩短,在角落区域停留时间显著增加。*:P<0.05,与对照组相比。
图1小鼠的社会回避行为
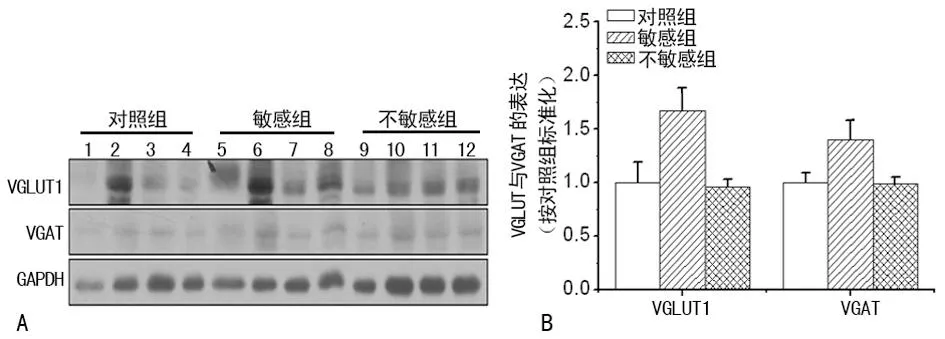
图2 抑郁症中前额叶皮层突触小体中囊泡转运蛋白的变化
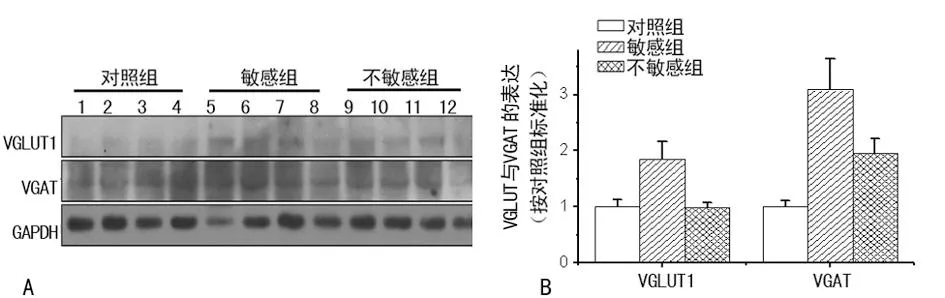
图3 抑郁症中海马突触小体中囊泡转运蛋白的变化
2.2皮层、海马及纹状体突触囊泡蛋白表达的变化在前额叶皮层、海马中与对照组和对抑郁症不敏感组比较,对抑郁症敏感组VGLUT1及VGAT的表达水平显著增加,差异有统计学意义(P<0.05);对抑郁症不敏感组与对照组相比其VGAT及VGLUT1水平的表达均差异无统计学意义,见图2、3。
在纹状体,虽然对抑郁症敏感组的小鼠VGAT及VGLUT1的表达水平均有增加(P<0.05),但是对抑郁症不敏感的小鼠这些蛋白表达也显著增加(图4)。

图4 抑郁症中纹状体突触小体中囊泡转运蛋白的变化
3讨论
大脑兴奋的传递和神经递质表达紊乱是近年来抑郁症的研究热点。但是,目前的研究大多数基于脑组织的全蛋白的提取,并不能确切反映突触中各种兴奋性和抑制性递质的变化。与前人的工作相比,作者在造模后对提取的突触蛋白进行检测,这使得样本更加单纯,并且提高了结果的特异性。前额叶皮层与海马均与多个脑区有突触的联系,是抑郁症发病的最关键脑区,纹状体是本研究的对照脑区。
VGLUT在突触系统中介导谷氨酸的传递,有3种存在形式:VGLUT1、2、3[8]。其中VGLUT1主要分布于大脑皮层、海马、小脑中,VGLUT2主要分布于皮层下及部分多巴胺神经元中[9-10],而VGLUT3则主要分布与特定的神经元中,如5-羟色胺能神经元、胆碱能神经元及GABA能和部分谷氨酸能神经元等[11]。研究发现,在抑郁症发病中,谷氨酸水平升高,NMDA受体过度激活;NMDA受体拮抗剂开他敏可产生快速的抗抑郁效应[7]。前额叶皮层和海马是抑郁症发病的两个重要脑区;作者选用VGLUT1探索前额叶皮层及海马的VGLUT表达,并发现抑郁症发病中该蛋白水平的上调。这提示抑郁症发病中兴奋性传递升高,并可能因此引起神经传递的紊乱。
GABA是大脑中最重要的抑制性神经递质,主要由谷氨酸脱羧酶合成[12]。除了作为神经递质的功能外,它还是神经发生及神经修复中重要的营养因子,并且参与调节氧化还原电位[13-14]。GABA与谷氨酸共同维持神经兴奋的平衡,这是避免情绪障碍等发生的关键。目前的研究普遍认为,在抑郁症发病中,谷氨酸浓度升高,GABA浓度降低并因此引起兴奋的失调。在本研究中并未发现VGAT水平的下降,这与以往的研究并不冲突,本样本来自突触,故VGAT不能代表总的GABA水平。对前额叶皮层和海马的研究结果提示抑郁症发病时负责GABA突触传递的囊泡蛋白VGAT水平显著增加,这可能是机体为了对抗谷氨酸受体的激活而采取的保护性措施。
经过社会失败造模后,小鼠纹状体内VGLUT1及VGAT表达均增加,而对抑郁症敏感组和对抑郁症不敏感组小鼠蛋白表达水平相比则差异无统计学意义(P>0.05)。这提示,在抑郁及抗抑郁过程中纹状体内VGLUT1和VGAT表达变化均增加,不具有脑区特异性。
本研究认为,经过社会失败造模后,小鼠前额叶皮层、海马等脑区突触前活动增多、神经递质紊乱,使得突触中VGLUT1及VGAT表达均增高。这提示,在抑郁症发病中兴奋性与抑制新神经递质释放增多,并可能引起突触功能紊乱,在临床工作中维持相应的神经递质的水平有可能有助于抗抑郁治疗。作者将对这些结论再进行深入探讨,探索治疗抑郁症的新突破点。
参考文献
[1]Holden C.Neuroscience.Mutant gene tied to poor serotonin production and depression[J].Science,2004,306(5704):2023-2025.
[2]Marona-Lewicka D,Nichols DE.The effect of selective serotonin releasing agents in the chronic mild stress model of depression in rats[J].Stress,1997,2(2):91-100.
[3]Smith KS,Rudolph U.Anxiety and depression:mouse genetics and pharmacological approaches to the role of GABA(A) receptor subtypes[J].Neuropharmacology,2012,62(1):54-60.
[4]Lussier AL,Romay-Tallón R,Caruncho HJ,et al.Altered GABAergic and glutamatergic activity within the rat hippocampus and amygdala in rats subjected to repeated corticosterone administration but not restraint stress[J].Neuroscience,2013,231(1):38-50.
[5]Zander JF,Münster-Wandowski A,Brunk I,et al.Synaptic and vesicular coexistence of VGLUT and VGAT in selected excitatory and inhibitory synapses[J].J Neurosci,2010,30(22):7634-7640.
[6]Krishnan V,Han MH,Graham DL,et al.Molecular adaptations underlying susceptibility and resistance to social defeat in brain reward regions[J].Cell,2007,131(2):391-400.
[7]Li N,Lee B,Liu RJ,et al.mTOR-dependent synapse formation underlies the rapid antidepressant effects of NMDA antagonists[J].Science,2010,329(5994):959-962.
[8]Moriyama Y,Yamamoto A.Glutamatergic chemical transmission:look! Here,there,and anywhere[J].J Biochem,2004,135(2):155-160.
[9]Fremeau RT,Troyer MD Jr,Pahner I,et al.The expression of vesicular glutamate transporters defines two classes of excitatory synapse[J].Neuron,2001,31(2):247-250.
[10]Fremeau RT,Voglmaier S,Seal RP,et al.VGLUTs define subsets of excitatory neurons and suggest novel roles for glutamate[J].Trends Neurosci,2004,27(2):98-100.
[11]Herzog E,Gilchrist J,Gras C,et al.Localization of VGLUT3,the vesicular glutamate transporter type 3,in the rat brain[J].Neuroscience,2004,123(4):983-990.
[12]Watanabe M,Maemura K,Kanbara K,et al.GABA and GABA receptors in the central nervous system and other organs[J].Int Rev Cytol,2002,213(1):1-5.
[13]Lamigeon C,Bellier JP,Sacchettoni S,et al.Enhanced neuronal protection from oxidative stress by coculture with glutamic acid decarboxylase-expressing astrocytes[J].J Neurochem,2001,77(2):598-600.
[14]Waagepetersen HS,Sonnewald U,Schousboe A.The GABA paradox:multiple roles as metabolite,neurotransmitter,and neurodifferentiative agent[J].J Neurochem,1999,73(4):1335-1340.
(漯河医学高等专科学校:1.病理生理学教研室;2.医疗系;3.生理学教研室,河南漯河 462002)
Expression of GABA and glutamate vesicular transporter in depressed mice*
Wu Qiong1,Song Jingfang2,Zhang Xiaoling1,Cai Hongyang1,Chang Quanzhong3△
(1.Department of Pathophysiology;2.Department of Medicine;3.Department of Physiology, Luohe Medical College,Luohe,Henan 462002,China)
[Abstract]ObjectiveTo study the expression of vesicular GABA transporter and vesicular glutamate transporter 1 in depression.MethodsMice was divided into control group and defeat group stochastically.By social defeat model and social avoidance,the defeat group was divided into two groups:susceptible group and unsusceptible group.Synaptic proteins were extracted respectively from the 3 groups.We detected the expression abundance of VGAT and VGLUT1 by Western blot.ResultsCompared with the control group,in susceptible group, the residence time in the contact area was significantly reduced, and the residence time in the corner area was significantly increased, with statistical difference(P<0.05).In the prefrontal cortex and hippocampus,Compared with the control group and the unsusceptible group,the expression levels of VGLUT1 and VGAT were increased in the susceptible group(P<0.05).there were no statistically significant in VGLUT1 and VGAT leveles between control group and the unsusceptible group(P>0.05).In the striatum,although the expression levels of VGAT and VGLUT1 were increased in susceptible group,but in unsusceptible group, the expression of these proteins also increased significantly.ConclusionThe prefrontal cortex and hippocampus excitability and inhibitory vesicle transport were changed in depression,which may relate to the transcription disorder.
[Key words]depression;GABA transporter;vesicular glutamate transporter
doi:论著·基础研究10.3969/j.issn.1671-8348.2016.11.009
* 基金项目:河南省高等学校重点科研项目基金(15B180012);漯河医学高等专科学校博士启动基金(2014-DF-004)。
作者简介:吴琼(1985-),副教授,博士,主要从事抑郁症发病机制研究。△通讯作者,E-mail:253043893@qq.com。
[中图分类号]R749.4
[文献标识码]A
[文章编号]1671-8348(2016)11-1470-03
(收稿日期:2015-10-25修回日期:2015-12-30)
