连作荞麦叶片衰老与活性氧代谢研究
2016-06-15高小丽张东旗妙佳源刘凤琴
高 扬,高小丽,张东旗,妙佳源,刘凤琴,赵 涛
(西北农林科技大学 农学院 旱区作物逆境生物学国家重点实验室,陕西 杨凌 712100)
连作荞麦叶片衰老与活性氧代谢研究
高扬,高小丽,张东旗,妙佳源,刘凤琴,赵涛
(西北农林科技大学 农学院 旱区作物逆境生物学国家重点实验室,陕西 杨凌 712100)
[摘要]【目的】 研究不同年限连作荞麦功能叶片衰老和活性氧代谢特性,探讨连作荞麦叶片衰老机理和产量形成的内部生理机制。【方法】 以‘西农9920’苦荞品种为材料,在连续4年定位试验基础上,以轮作荞麦为对照,研究了不同连作年限荞麦功能叶片叶绿素、可溶性蛋白含量、SOD和POD活性及膜质过氧化产物丙二醛(MDA)、氧自由基含量等生理指标的变化,收获时测定不同连作年限和轮作荞麦的产量、千粒质量、株高和主茎节数,最后对荞麦生育后期功能叶片生理指标与产量的相关性进行了分析。【结果】 荞麦开花后,随着生育进程的推移,功能叶片的叶绿素、可溶性蛋白含量、SOD活性均呈先上升后下降趋势,POD活性、MDA和氧自由基含量持续上升。随着连作年限的增加,荞麦开花后功能叶片叶绿素和可溶性蛋白含量明显下降;MDA和氧自由基含量上升,且连作高于轮作;SOD活性随着连作年限的增加呈现先升高再下降趋势;POD活性在开花前中期(开花后7~14 d)随连作年限的增加呈显著降低趋势,而在生育后期(开花后21 d之后)随连作年限的增加呈显著升高趋势。随着连作年限的增加,荞麦籽粒产量、株高、主茎节数均降低,且均低于轮作荞麦,但连作与轮作荞麦之间千粒质量差异不显著。相关分析表明,荞麦产量与花后功能叶片的可溶性蛋白含量呈极显著正相关,与叶绿素含量呈显著正相关,与POD活性、MDA以及氧自由基含量呈极显著或显著负相关。【结论】 随着连作年限的增加,叶片功能期缩短,活性氧伤害程度加剧,籽粒产量降低。
[关键词]荞麦;连作;轮作;活性氧代谢;产量;叶片衰老
荞麦(FagopyrumesculentumMoench)耐旱、耐瘠、适应性强、生育期短,在作物布局中占有特殊地位[1],同时也是备荒救灾、填闲补缺最理想、最经济的优势作物[2]。荞麦营养价值高,富含蛋白质且氨基酸组成平衡,此外还含有其他作物少含或不含的生物类黄酮等生物活性物质[3],食药兼用,是21世纪重要的健康食品资源。但我国荞麦多分布在贫瘠丘陵旱地,受自然条件的限制,一年一作,加之产地的种植传统、市场需求等,导致荞麦连作普遍,致使荞麦生育状况变差、产量降低、品质变劣。
植物细胞代谢过程中会产生活性氧(Reactive oxygen species,ROS),正常生理代谢下ROS的产生与清除处于动态平衡状态,而当植物处于逆境环境时这种动态平衡会被打破,出现氧化胁迫,进而损伤细胞的生物大分子和生物膜[4]。植物体内存在一套完善的抗氧化防御系统,具有清除活性氧的能力,其中研究最广泛的就是超氧化物歧化酶(SOD)、过氧化物酶(POD)等酶促系统保护酶;丙二醛(MDA)是脂膜过氧化过程中产生的新自由基,往往能造成氧化伤害,影响植株正常的生长发育[5]。近些年来,随着生物活性氧毒害的超氧化物学说的创立,有关逆境胁迫下作物叶片活性氧代谢的研究越来越受到人们的重视[6]。有研究认为,在水分胁迫下荔枝植株叶片的可溶性蛋白含量以及SOD、POD、CAT、抗坏血酸还原酶等重要酶的活性都有所下降[7];Dhindsa等[8]则发现,水分胁迫能引起SOD和CAT活性上升;于福军等[9]研究发现,低温处理时苜蓿幼苗叶片内SOD、POD活性始终维持在一个较高的水平,说明苜蓿可通过维持较高水平的保护酶活性来适应低温环境胁迫,缓解低温伤害;张新慧等[10]研究发现,连作条件下当归的SOD、POD、CAT等保护酶活性均呈下降趋势,且随连作年限的增加这种下降趋势加剧。
本试验以不同连作年限荞麦为材料,研究不同连作年限荞麦开花后功能叶片保护酶活性、可溶性蛋白和氧自由基等指标的变化,旨在揭示连作下荞麦植株衰老、产量变化与活性氧代谢、叶绿素含量的关系,为荞麦连作障碍生理机制研究和高产优质栽培提供理论依据。
1材料与方法
1.1材料与试验地概况
试验以“西农9920”苦荞品种为供试材料,试验地设在西北农林科技大学北校区农作一站,该站地处渭河三级阶地,平均气温12.9 ℃,降水量550.8 mm,其中7-9月份降水量占全年降水量的60%以上,年均蒸发量为1 400 mm,属于半湿润易旱地区,2010年播前试验地土壤的基本理化性质为:全氮 1.24 g/kg,全磷1.05 g/kg,速效氮87.1 mg/kg,速效磷4.51 mg/kg,速效钾158.8 mg/kg,pH 8.3。
1.2试验设计与样品采集和处理
2010-2013年进行了连续4年的定位试验,设荞麦连作2年(T1)、连作3年(T2)、连作4年(T3) 3个处理,对照为轮作荞麦,轮作顺序为荞麦-谷子-芸豆-荞麦。小区面积20 m2(4 m×5 m),4次重复,完全随机设计排列,生育期间不施肥,进行常规的田间管理。于2011,2012,2013年荞麦开花期开始,每隔7 d在田间摘取具有代表性的荞麦叶片(取荞麦开花节位上数1~3片叶),每个小区随机取3株,放入冰壶,带回实验室。洗净叶片表面尘土和污物,吸水纸小心擦干表面水分,去除叶脉,剪碎至5 mm以下,混匀待用。
在分析天平上准确称取0.500 g剪碎的样品,置于预冷的小研钵中,将8 mL预冷的0.05 mol/L、pH 7.8的Na2HPO4-NaH2PO4缓冲液分2次加入,然后加入少量石英砂和液氮,研磨成匀浆后,转移至离心管中,于2 ℃、20 000×g下冷冻离心20 min,上清液即为酶提取液,将上清液倒入干燥并已预冷的具塞磨口试管中,盖上塞子,置于冰水浴中,再冷藏于4 ℃冰箱中备用。
1.3测定项目及方法
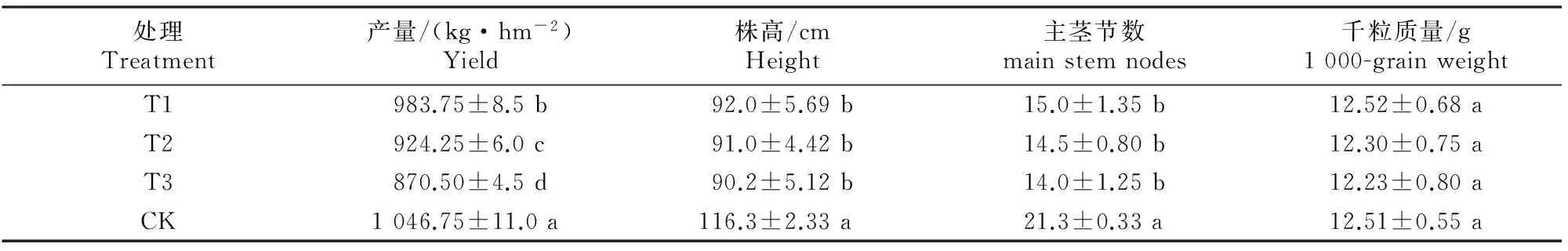
超氧化物歧化酶(SOD)活性测定采用NBT光化还原法[11]。过氧化物酶(POD)活性测定采用Sigma法[12]。可溶性蛋白含量测定采用考马斯亮蓝法[13]。丙二醛(MDA)含量测定采用硫代巴比妥酸法[14]。氧自由基含量测定采用盐酸羟胺法[15]。叶绿素含量测定采用丙酮浸提法[16]。收获每个小区籽粒晒干后称质量换算成单位面积产量,挑选 1 000 粒大小均匀籽粒称质量得到千粒质量,株高和主茎节数均为现场测量。
1.4数据处理与分析
试验数据采用Excel 2003进行统计,DPS 7.05进行方差分析,SigmaPlot 10.0制图。多重比较采用Duncan’s新复极差法测验。
2结果与分析
2.1连作荞麦功能叶片叶绿素及可溶性蛋白含量的变化
由图1可以看出,荞麦开花后功能叶片叶绿素含量随着生育进程的推进表现为先升高后降低的变化趋势,且均于开花后第14天达到高峰。随着连作年限的增加,荞麦功能叶片叶绿素含量呈下降趋势,即T1>T2>T3,且连作低于轮作(CK)。在开花后第14天,轮作(CK)功能叶片叶绿素含量分别是连作2年(T1)、连作3年(T2)和连作4年(T3)的1.0~1.4倍。
由图1还可以看出,随着生育期的推进,荞麦开花后功能叶片可溶性蛋白含量呈先升后降趋势;随着连作年限的增加,可溶性蛋白含量呈明显降低趋势,表现为CK>T1>T2>T3,与轮作相比,连作2年(T1)、连作3年(T2)和连作4年(T3)荞麦可溶性蛋白含量分别下降了4.35%,10.87%和32.61%,且差异显著。
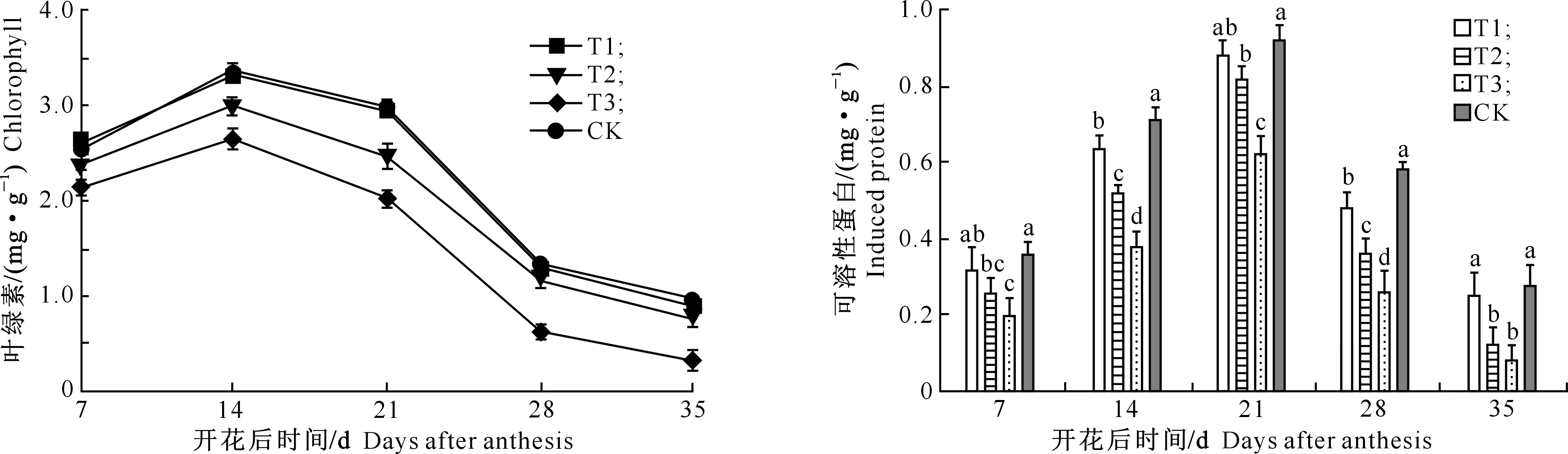
图 1 连作对荞麦功能叶片叶绿素及可溶性蛋白含量的影响
2.2连作荞麦功能叶片氧自由基和丙二醛含量的变化
由图2可以看出,荞麦开花后功能叶片氧自由基含量随着生育进程的推进呈上升趋势。随着连作年限的增加,氧自由基含量显著升高,即T1 由图2还可以看出,荞麦开花后功能叶片MDA含量表现出与氧自由基含量相似的变化趋势,即随着生育进程的推进呈增长趋势。随着连作年限的增加,MDA含量显著升高(P<0.05),且连作高于轮作(CK)。 2.3连作荞麦功能叶片保护性酶活性的变化 由图3可以看出,荞麦开花后功能叶片SOD活性随生育进程的推进总体呈先上升后下降的变化趋势。随着连作年限的增加,SOD活性表现为缓慢增加后急剧下降的变化趋势,具体表现为连作3年(T2)>连作2年(T1)>连作4年(T3),且轮作(CK)SOD活性大体上处于T1、T2之间,T1、T2与CK之间差异不显著,但均与T3差异达显著水平(P<0.05)。 图 2 连作对荞麦功能叶片氧自由基及丙二醛含量的影响 由图3还可以看出,荞麦开花后功能叶片POD活性随生育进程的推进呈上升趋势;在开花后不同时间随连作年限的增加表现不同,开花初中期(开花后7~14 d)连作荞麦功能叶片POD活性较轮作低,且随连作年限的增加呈显著降低趋势,即CK>T1>T2>T3,如开花后第14天,轮作(CK)的POD活性分别是连作2年(T1)、连作3年(T2)和连作4年(T3)的1.09,1.18和1.42倍;而在生育后期(开花后21 d后),连作荞麦POD活性则明显高于轮作(CK),且随连作年限的增加呈升高趋势。 图 3 连作对荞麦功能叶片保护性酶活性的影响 2.4连作对荞麦产量及主要生育性状的影响 由表1可以看出,随着连作年限的增加,荞麦籽粒产量呈下降趋势,表现为T1>T2>T3(P<0.05),且均显著均低于轮作(CK)产量,轮作(CK)产量分别是连作2年(T1)、连作3年(T2)、连作4年(T3)的1.06,1.13和1.20倍,且差异显著。与轮作(CK)相比,连作显著降低了荞麦植株的株高、主茎节数,而二者千粒质量差异不显著。 表 1 不同连作年限荞麦的产量及主要生育性状 注:同列数据后标不同小写字母表示同期轮作和连作处理间在P<0.05水平存在显著性差异。 Note:Values followed by different small letters within a column are significantly different between continuous cropping and rotation cropping at the same stage at theP<0.05 levels respectively. 2.5荞麦生育后期功能叶片生理指标与产量的相关性 由表2可知,荞麦生育后期,功能叶片可溶性蛋白含量与产量呈极显著正相关;叶绿素含量与产量呈显著正相关;SOD活性与产量呈正相关;氧自由基、MDA含量与产量呈显著负相关;POD活性与产量呈极显著负相关。 表 2 荞麦生育后期功能叶片生理指标与产量的相关性 注:*和**分别表示在P<0.05和P<0.01水平有显著性关系。 Nate:* and ** indicate significance atP<0.05 andP<0.01,respectively. 3讨论 许多研究表明,叶片衰老过程是体内活性氧、自由基代谢失调的累积过程[17-20]。本研究与前人在其他作物上的研究结果基本一致[21-23],即随着荞麦植株的衰老,功能叶片的叶绿素、可溶性蛋白质含量和SOD活性均先上升后下降,膜质过氧化产物MDA和氧自由基含量持续上升,导致细胞膜结构和功能受到破坏。与轮作相比,连作会加速功能叶片的衰老进程,且随着连作年限的增加,荞麦功能叶片叶绿素和可溶性蛋白含量明显下降,MDA和氧自由基含量上升,籽粒产量下降。 SOD是植物体内清除活性氧物质的重要酶,被誉为植物抵御逆境伤害的第一道防线[24]。赵淑英等[25]研究表明,连作导致大豆叶片SOD活性下降、MDA含量增多。本研究结果表明,随着连作年限的增加,荞麦功能叶片SOD活性呈现先升高再下降趋势,尤其是连作3年的功能叶片SOD活性高于连作2年和4年,这可能是由于荞麦在短期轮作(3年内)条件下,叶片为了防御活性氧伤害,出现了SOD诱导合成、活性升高现象,但这只是暂时的,之后随着连作年限的增加,这种反应消失,SOD活性下降。其具体原因有待进一步研究。 POD在植物体内的作用具有双重性[26-28]:在衰老前期表达,表现为保护效应;在逆境和衰老后期表达,参与活性氧的生成,表现为伤害效应。但不同植物种类以及同一植物的不同品种在其生长发育及衰老过程中,POD活性表现不同[29]。本研究结果表明,荞麦开花后,随着生育进程的推进,功能叶片POD活性升高,说明POD在荞麦叶片衰老过程可能参与了活性氧的产生,这与宋慧等[28]、高小丽等[30]在小豆、绿豆上的研究结果一致,但与刘晓蕾等[31]、闫利恒等[32]、沙伟等[33]在禾谷类作物上的研究结果不完全一致。本研究结果还表明,POD活性在荞麦开花前中期(开花后7~14 d)随连作年限的增加呈显著降低趋势,而在生育后期(开花后21 d后)随连作年限的增加呈显著升高趋势。这可能与POD功能的多样性有关,具体生理机制有待进一步研究。 同时,作为化感物质的重要来源之一,根系分泌物在土壤养分活化及土壤与环境之间物质交流方面有着重要作用,对作物衰老及增减产具有一定的指导意义。本试验后期对连作4年荞麦进行了整株植物的移栽培养,提取其根系分泌物并进行GC-MS气质联用仪分离鉴定,检测出了2,4-二叔丁基苯酚、邻苯二甲酸二辛酯、2,3 -二羟基丙基酯等相对丰度较高的酚、酯类分泌物,这些分泌物均是自毒作用强于他感作用的化感物质,可能是因为连作胁迫诱导出了这些酚、酯类物质,从而促使连作荞麦体内活性氧累积、自由基代谢失调,并最终导致作物减产。关于连作荞麦根系分泌物对植株衰老及减产的抑制机理还需要更进一步的探索研究。 [参考文献] [1]刘邻渭,章华伟,姜莉.中国荞麦深加工的探索 [J].西北农林科技大学学报:自然科学版,2002,30(1):83-86. Liu L W,Zhang H W,Jiang L.Research into buckwheat deep processing in China [J].Journal of Northwest A&F University:Natural Science Edition,2002,30(1):83-86.(in Chinese) [2]林汝法,周小理,任贵兴,等.中国荞麦的生产与贸易、营养与食品 [J].食品科学,2005,26(1):259-263. Lin R F,Zhou X L,Ren G X,et al.Production and trading of buckwheat in China,nutrition and food [J].Food Science,2005,26(1):259-263.(in Chinese) [3]文平,陈进红.荞麦芦丁的研究进展 [J].中国粮油学报,2006,21(3):107-111. Wen P,Chen J H.Research progress on rutin in buckwheat [J].Journal of the Chinese Cereals and Oils Association,2006,21(3):107-111.(in Chinese) [4]王永军,杨今胜,袁翠平,等.超高产夏玉米花粒期不同部位叶片衰老与抗氧化酶特性 [J].作物学报,2013,39(12):2183-2191. Wang Y J, Yang J S,Yuan C P,et al.Characteristics of senescence and antioxidant enzyme activities in leaves at different plant parts of summer maize with the super-high yielding potential after anthesis [J].Acta Agronomica Sinica,2013,39(12):2183-2191.(in Chinese) [5]张帆,郁继华,颉建明,等.外源ALA和Spd对低温弱光下辣椒幼苗光合作用及抗氧化系统的影响 [J].中国农业科学,2013,46(11):2298-2306. Zhang F,Yu J H,Xie J M,et al.Influence of exogenous ALA and Spd on photosynthesis and antioxidant system of low temperature and poor light affected pepper seedlings [J].Scientia Agricultura Sinica,2013,46(11):2298-2306.(in Chinese) [6]张永征,李海东,李秀,等.水光互作对生姜叶片活性氧代谢的影响 [J].应用生态学报,2013,24(12):3459-3464. Zhang Y Z,Li H D,Li X,et al.Effects of water and light interaction on reactive oxygen metabolism in ginger leaves [J].Chinese Journal of Applied Ecology,2013,24(12):3459-3464.(in Chinese) [7]陈立松,刘星辉.水分胁迫对荔枝叶片氮和核酸代谢的影响及其与抗旱性的关系 [J].植物生理学报,1999,25(1):49-56. Chen L S,Liu X H.Effects of water stress on nitrogen and nucleic acid metabolisms in litchi(LitchichinensisSonn.) leaves and their relations to drough-tresistance [J].Acta Phytophysiologica Sinica,1999,25(1):49-56.(in Chinese) [8]Dhindsa R S,Matowe W.Drought tolerance in two mosses:Co-rrelated with enzymatic defence against lipid peroxidation [J].J Exp Bota,1981,32(126):79-91. [9]于福军,谭大海,冯昌军.6个美国苜蓿品种低温胁迫下SOD、POD活性的变化 [J].齐齐哈尔师范高等专科学校学报,2006(3):101-102. Yu F J,Tan D H,Feng C J.Change of SOD and POD activities about six kinds of American alfalfa varieties in the stress of low temperature [J].Journal of Qiqihar Junior Terchers’ College, 2006(3):101-102.(in Chinese) [10]张新慧,张恩和,王惠珍,等.连作对当归生长的障碍效应及机制研究 [J].中国中药杂志,2010,35(10):1231-1234. Zhang X H,Zhang E H,Wang H Z,et al.Effects of continuous cropping obstacle on growth ofAngelicasinensisand its mechanism [J].China Journal of Chinese Materia Medica,2010,35(10):1231-1234.(in Chinese) [11]王爱国,罗广华,邵从本,等.大豆种子超氧物歧化酶的研究 [J].植物生理学报,1983,9(1):77-84. Wang A G,Luo G H,Shao C B,et al.A study on the superoxide dismutase of soybean seeds [J].Acta Phytophysiologia Sinica,1983,9(1):77-84.(in Chinese) [12]施特尔马赫 B.酶的测定方法 [M].钱嘉渊,译.北京:中国轻工出版社,1992:186-194. Stellmach B.Determination of enzyme [M].Qian J Y,translation.Beijing:China Light Industry Press,1992:186-194.(in Chinese) [13]黄婉玉,曹炜,李菁,等.考马斯亮蓝法测定果汁中蛋白质的含量 [J].食品与发酵工业,2009,35(5):160-162. Huang W Y,Cao W,Li Q,et al.Determination of protein content in juice by coomassie brilliant blue [J].Food and Ferm Entation Industries,2009,35(5):160-162.(in Chinese) [14]Heath R L,Packer L.Photoperoxidation in isolated chloropla-sts:Ⅰ.Kinetics and stoichiometry of fatty acid peroxidation [J].Archives Biochem Biophysics,1968,125:189-198. [15]萧华山,何文锦,傅文庆,等.一种用分光光度计检测氧自由基的新方法 [J].生物化学与生物物理进展,1999,26(2):83-85. Xiao H S,He W J,Fu W Q,et al.A spectrophotometer method testing oxygen radicals [J].Prog Biochem Biophys,1999,26(2):83-85.(in Chinese) [16]陈毓荃.生物化学实验方法和技术 [M].北京:科学出版社,2002:95-97. Chen Y Q.The experimental means and technology of biochemistry [M].Beijing:Science Press,2002:95-97.(in Chinese) [17]Kaiser W.The effect of hydrogen peroxide on CO2fixation of isolated intact chloroplasts [J].Biochemica et Biophysica Acta,1976,440:476-482. [18]Maksimov I V,Troshina N B,Surina O B,et al.Influence of Ca2+ions on metabolism of active oxygen species in wheat calli cocultured with the bunt pathogenTilletiacaries[J].Applied Biochemistry and Microbiology,2010,46(5):530-535. [19]冯佰利,高小丽,王长发,等.干旱条件下不同温型小麦叶片衰老与活性氧代谢特性的研究 [J].中国生态农业学报,2005,13(4):74-76. Feng B L,Gao X L,Wang C F,et al.Leaf senescence and active oxygen metabolism of different-type wheats under drought [J].Chinese Journal of Eco-Agriculture,2005,13(4):74-76.(in Chinese) [20]Leshem Y Y.Oxygen free radicals and plant senescence [J].What’s New in Plant Physiol,1981,12:1-4. [21]高小丽,高金锋,冯佰利,等.不同绿豆品种生育后期叶片衰老的研究 [J].西北植物学报,2007,27(5):947-953. Gao X L,Gao J F,Feng B L,et al.Leaf senescence of different mung bean varieties during their late growth [J].Acta Bot Boreal-Occident Sin,2007,27(5):947-953.(in Chinese) [22]魏国强,朱祝军,方学智,等.NaCl胁迫对不同品种黄瓜幼苗生长、叶绿素荧光特性和活性氧代谢的影响 [J].中国农业科学,2004,37(11):1754-1759. Wei G Q,Zhu Z J,Fang X Z,et al.The effects of NaCl stress on plant growth,chlorophyll fluorescence characteristics and active oxygen metabolism in seedlings of two cucumber cultivars [J].Scientia Agricultura Sinica,2004,37(11):1754-1759.(in Chinese) [23]Yan Y Q,Che D D,Shi X C,et al.Effects of salt-alkali stress on active oxygen metabolism in roots of Spiraea×bumalda ‘Gold Mound’ and Spiraea×bumalda ‘Gold Flame’ [J].Journal of Forestry Research,2011,1:59-64. [24]Savitch L V,Nassacci A,Gray G R,et al.Acclimation to low temperature or light mitigates sensitivity to photo inhibition:Roles of the Calvin cycle and the Mehler reaction [J].Aust J Plant Physiol,2000,27:253-264. [25]赵淑英,赵九洲,陈洁敏,等.连作对大豆生理生化特性的影响 [J].大豆科学,1995,14(2):113-118. Zhao S Y,Zhao J Z,Chen J M,et al.Effect of soybean successive cropping on physiological and biochemical index of soybean [J].Soybean Science,1995,14(2):113-118.(in Chinese) [26]王冬雪,张丽莉,石瑛.不同熟性马铃薯各生育时期功能叶生理指标变化的研究 [J].中国农学通报,2014,30(3):124-128. Wang D X,Zhang L L,Shi Y.Research of the physiological indicators of different maturity potato (SolanumtuberosumL.) functional leaf at each growth stages [J].Chinese Agricultural Science Bulletin,2014,30(3):124-128.(in Chinese) [27]高福元,张吉立,刘振平.冬季低温对4种彩叶植物SOD、POD活性影响的研究 [J].中国农学通报,2010,26(5):169-173. Gao F Y,Zhang J L,Liu Z P.Studies on effects of four species of color-leafed plants on SOD,POD activity of the cold-resistance in winter [J].Chinese Agricultural Science Bulletin,2010,26(5):169-173.(in Chinese) [28]宋慧,冯佰利,高小丽,等.不同小豆品种(系)叶片衰老与活性氧代谢 [J].作物学报,2010,36(2):347-353. Song H,Feng B L,Gao X L,et al.Leaf senescence and reactive oxygen metabolism in different adzuki bean cultivars (lines) [J].Acta Agronomica Sinica,2010,36(2):347-353.(in Chinese) [29]鱼欢,冯佰利,张英,等.不同栽培模式下冬小麦叶片衰老与活性氧代谢研究 [J].作物学报,2007,33(10):1729-1732. Yu H,Feng B L,Zhang Y,et al.Leaf aging and reactive oxygen species metabolism of winter wheat in different cultivation modes [J].Acta Agronomica Sinica,2007,33(10):1729-1732.(in Chinese) [30]高小丽,孙健敏,高金锋,等.不同基因型绿豆叶片衰老与活性氧代谢研究 [J].中国农业科学,2008,41(9):2873-2880. Gao X L,Sun J M,Gao J F,et al.Leaf aging and reactive oxygen metabolism in different genotypes of mung bean [J].Scientia Agricultura Sinica,2008,41(9):2873-2880.(in Chinese) [31]刘晓蕾,赵欣,韩蕊莲,等.水分胁迫下玉米幼苗对外源原儿茶醛的生理响应 [J].西北农业学报,2011,20(1):76-81. Liu X L,Zhao X,Han R L,et al.Physiological responses of maize seedlings to the exogenous protocatechuic aldehyde pretreatment under water stress [J].Acta Agriculturae Boreali-Occidentalis Sinica,2011,20(1):76-81.(in Chinese) [32]闫利恒,代海芳,贺杰.小麦春化脱春化、抗寒处理后POD活性变化的研究 [J].中国农学通报,2014,30(9):99-104. Yan L H,Dai H F,He J.The Research on POD isozyme activity change of wheat after treatmention of vernalization devernalization or cold acclimation [J].Chinese Agricultural Science Bulletin, 2014,30(9):99-104.(in Chinese) [33]沙伟,刘焕婷,谭大海,等.低温胁迫对扎龙芦苇SOD、POD活性和可溶性蛋白含量的影响 [J].齐齐哈尔大学学报,2008,24(2):1-4. Sha W,Liu H T,Tan D H,et al.The effect of low temperature stress on the SOD,POD activity and the soluble protein contents ofPhragmitesaustralisin Zhalong [J].Journal of Qiqihar University,2008,24(2):1-4.(in Chinese) Leaf senescence and reactive oxygen metabolism of buckwheat under continuous cropping GAO Yang,GAO Xiao-li,ZHANG Dong-qi,MIAO Jia-yuan,LIU Feng-qin,ZHAO Tao (CollegeofAgriculture,NorthwestA&FUniversity,StateKeyLaboratoryofAridCropStressBiology,Yangling,Shaanxi712100,China) Abstract:【Objective】 This study aimed to research functional leaf senescence and reactive oxygen metabolism of buckwheat at different years in continuous cropping system,and investigate the mechanism of leaf senescence and the internal physiological mechanism of yield formation.【Method】 Taking ‘Xinong 9920’ tartary buckwheat as material and rotation cropping system as control (CK),a series of experiments were conducted on the basis of 4 years to research the change in contents of chlorophyll and soluble protein,activities of SOD and POD,and accumulation of MDA and OFR of buckwheat’s functional leaf.The yield,1 000 grain mass,height and number of main stem sections were also measured when harvesting and their relationship with leaf physiological indexes was analyzed.【Result】 With the continuation of aging process,SOD activity and contents of chlorophyll and soluble protein decreased,while POD activity and MDA and OFR accumulation increased continuously after anthesis.With the increase of continuous cropping years,contents of chlorophyll and soluble protein decreased,MDA and OFR accumulation increased,and these values of continuous cropping were higher than that of rotation cropping.SOD activity firstly increased followed by decrease.POD activity decreased significantly before anthesis and increased significantly in the late growth stage (21 d after anthesis).The grain yield had a very significant positive correlation with soluble protein content,a significant positive correlation with chlorophyll content,and a very significant or significant negative correlation with POD activity as well as MDA and OFR accumulation.【Conclusion】 With the increase of continuous cropping years,the functional period of leaf shortened,the degree of reactive oxygen damage aggravated,and the grain yield decreased. Key words:buckwheat;continuous cropping;rotation;reactive oxygen metabolism;grain yield;leaf senescence DOI:网络出版时间:2016-01-0810:2210.13207/j.cnki.jnwafu.2016.02.005 [收稿日期]2014-07-11 [基金项目]国家自然科学基金项目(31071472);陕西省科学技术研究发展计划项目(2014K01-28) [作者简介]高扬(1990-),男,安徽宣城人,硕士,主要从事植物安全生产研究。 E-mail:gao.yang@nwsuaf.edu.cn[通信作者]高小丽(1968-),女,陕西宁强人,副教授,博士,主要从事小宗粮豆作物高产栽培生理及产业开发研究。 [中图分类号]S517 [文献标志码]A [文章编号]1671-9387(2016)02-0028-07 E-mail:gao2123@nwsuaf.edu.cn