间伐对鼠类扩散林木种子的影响
2016-06-15张义锋王魏瑞苗向东路纪琪
张义锋, 王魏瑞, 李 蔚, 苗向东, 路纪琪
(1.郑州大学 生命科学学院 河南 郑州 450001; 2.郑州大学 基础医学院 河南 郑州 450001;3.国有济源市愚公林场 河南 济源 454650)
间伐对鼠类扩散林木种子的影响
张义锋1,王魏瑞1,李蔚2,苗向东3,路纪琪1
(1.郑州大学 生命科学学院河南 郑州 450001; 2.郑州大学 基础医学院河南 郑州 450001;3.国有济源市愚公林场河南 济源 454650)
摘要:间伐作为一种林业生产活动,将会对区域内的植物群落和动物种群及行为产生影响.为了解间伐对鼠类介导的种子扩散的影响,2012年4月在王屋山区选择未间伐和间伐林地作为样地,标记、释放山杏 (Prunus armeniaca) 种子,调查鼠类对种子扩散的影响.研究表明:1) 未间伐区鼠类的捕获率为6.7%,间伐区为2.7%;2) 间伐区乔木密度降低56.3%、灌丛密度降低72.7%;3) 未间伐区种子的扩散速率 (半存留时间2 d) 显著快于间伐区 (半存留时间10 d);4) 未间伐区,鼠类对种子的埋藏深度为(4.3±0.1)cm,间伐区为(3.9±0.2)cm,二者差异显著;5) 未间伐区,鼠类对种子的多次扩散率为20.3%,间伐区为4.4%,二者差异显著.结果显示,林木间伐导致林区生境和鼠类种群显著变化,造成鼠类贮食行为和种子命运发生改变,最终将影响林区的幼苗更新.
关键词:林木间伐; 鼠类; 贮食行为; 种子命运; 埋藏深度
0引言
鼠类是林木种子的重要捕食者和扩散者,许多鼠类在遇到林木种子时,除取食一定量的种子外,还将部分种子搬运到远离母树的灌丛、草丛等处贮藏以备将来之需[1].对植物而言,鼠类的扩散降低了母树下种子的密度,有利于减弱种子之间的生存竞争,提高种子的发芽率和存活率,从而对植物的幼苗建成、群落演替和生态系统功能的维持具有积极作用[2—3].因此,鼠类对种子的扩散和贮藏方式对林木种子的命运、幼苗更新等有重要影响[4—5].
在自然条件下,干扰能显著影响动物的食物贮藏行为[6—7].例如,干扰能降低种子扩散者的数量、改变动物对种子的扩散量和扩散距离[6];人类活动干扰使鼠类的分散贮藏行为减弱[8];一定程度的放牧干扰能刺激鼠类的分散贮藏行为,进而促进植物种子扩散[9];种间和种内竞争使鼠类食物贮藏策略发生改变或调整[10—11];在捕食风险较低、植被茂密的生境中,动物对种子的捕食率更高[12—13].因此,不同强度和方式的干扰会对森林生境中鼠类的多样性[6—7]和贮食行为产生不同的影响[14—15],最终影响种子扩散、萌发和林木更新.
间伐是一种常见的林业生产活动,按照一定的标准进行选择性采伐,在获得木材的同时,尽量减少对森林生态功能的危害,在不影响生态平衡的前提下实现经济利益[16—18].但是,在现实操作中,经济价值较高的大树和树种往往被优先采伐,以获得更高的经济效益,因此林木间伐往往导致植物群落组成、物种多样性、林分空间结构、植被覆盖度等发生较大改变[6,17].栖息地的改变使以植物叶、种子、果实等为食的林栖鼠类的多样性和种群密度显著降低[7,19],进而影响到森林种子的扩散[20]及植物群落的演替进程[21].
研究表明,间伐能使动物对种子的扩散增加[20]或者减少[7,22],也有研究发现,间伐对种子的扩散速率没有影响,但可使种子的扩散距离减小[22].由此可见,关于间伐对动物食物贮藏行为和种子命运的影响还需要深入研究.
王屋山愚公地区存在大面积次生林,间伐是常见的管理措施之一,而间伐不可避免地会对林区内的植被和林栖动物造成影响,因此,本研究选取未间伐和间伐林为样地进行种子扩散研究,旨在探讨:1) 间伐对森林生境中鼠类种类和相对密度的影响;2) 间伐对鼠类食物贮藏行为的影响;3) 间伐对种子扩散及种子命运的影响.以期完善林业生产与生态保护的理论,为林区林业生产和生态保护提供理论支持,最大化实现林区的经济价值和生态价值.
1材料与方法
1.1研究地区和样地选择
本研究在国有济源市愚公林场 (E 112°16′,N 35°12′) 进行.该林场位于河南省济源市西北约40 km处,地处太行山南麓,平均海拔600 m,属大陆性季风气候,年均气温14.3 ℃,年平均降水量600~700 mm.该地区的植被可分为针叶林、阔叶林和灌丛-灌草丛3种类型.样地植被以栓皮栎 (Quercusvariabilis) 为建群种,另有少量的刺槐 (Robiniapseudoacacia),林下灌丛植被以荆条 (Vitexnegundovar.heterophylla)、黄刺玫 (Rosaxanthina)、黄栌 (Cotinuscoggygria) 等为主[23].鼠类是本地区林木种子的主要取食和扩散者,主要鼠种为岩松鼠 (Sciurotamiasdavidianus)、花鼠 (Eutamiassibiricus)、社鼠 (Niviventerconfucianus)、大林姬鼠 (Apodemuspeninsulae)等[24].
实验设置间伐样地和对照样地进行实验,间伐样地为间伐后30 d的次生林,盖度40%左右.对照样地为未间伐的次生林,盖度85%左右;两类样地坡向一致,坡度约30°,坡向朝东南,每块样地面积不小于2 hm2.
1.2鼠类和植被调查
为了解间伐对林栖鼠类的影响,采用笼捕法 (29 cm×13 cm×12 cm),对间伐和未间伐样地内的鼠类及其相对密度进行调查.在每类样地内沿坡向各选取一条样线,沿样线布放捕鼠笼25个,笼间距约5 m,以花生为诱饵,第二天7:30—8:00和18:30—19:00各检查一次,记录鼠种、性别等信息后原地释放,调查持续3 d.
为了解间伐对植被的影响,在间伐样地和未间伐样地,沿坡向各选取3个10 m×10 m样方,样方距林缘30 m以上,样方间距30 m,调查并记录样方内乔木和灌木(簇)的种类和数量.
1.3种子标记和释放
在样地中,沿坡向设置两条样线,样线间隔50 m,每条样线设置6个种子释放点 (约1 m2),释放点间距为25 m.
山杏(Armeniacasibirica)是当地的主要植被之一,在实验样地内有零星分布,在栓皮栎林边缘成片分布.在种子成熟季节,采集山杏种子阴干备用.种子标记采用塑料片标记法[25—26],在种子外壳(内果皮)边缘钻一小孔,不伤及子叶和胚芽,用长约10 cm的细钢丝将塑料标记牌(2.5 cm×3.5 cm)系于小孔处,对每个标记牌进行编号,以便对种子命运进行追踪和调查[27].
于2012年4月,在每个释放点释放已标记的山杏种子50枚,共计50×12×2=1 200枚.种子释放后前10 d,每天检查种子的扩散情况,10天后每隔5 d检查一次种子状态,调查期为60 d.根据标记牌上的编号检查并记录种子的扩散情况、种子命运、埋藏深度、扩散距离、种子的多次扩散等.
种子命运包括:1) 原地存留 (intactinsitu, IIS);2) 取食 (eateninsituor after removal, E); 3) 弃置地表 (intact after removal, IAR);4) 分散贮藏 (scatter hoarding, SH);5) 丢失 (missing, M).
1.4数据分析
采用SPSS for windows (version 13.0) 对数据进行统计分析.统计数据用均数±标准误表示.用survival analysis from Kaplan-Meier检验两种样地内种子扩散速率的差异.用Mann-WhitneyU检验比较山杏种子在不同样地的原地存留、取食、贮藏及丢失的差异.用independent samplest-test检验鼠类对种子埋藏深度的差异.用Chi-square test检验鼠类对山杏种子多次扩散的差异.
2结果
2.1鼠类和植被
在未间伐样地中,共捕获岩松鼠、社鼠和大林姬鼠3种,总捕获率为6.7%;在间伐样地中仅捕获大林姬鼠,捕获率为2.7%.
未间伐样地中的乔木种类为栓皮栎和刺槐2种,灌丛的组成主要包括荆条、黄栌及黄刺玫3种;间伐样地中乔木只有栓皮栎,灌丛只有荆条.在未间伐样地中,乔木的密度为(0.5±1.2)棵/m2,而在间伐样地中,乔木密度为(0.2±0.9)棵/m2,减少56.3%;未间伐样地中,灌丛密度为(0.2±1.2)簇/m2,而间伐样地中,灌丛密度为(0.1±1.7)簇/m2,减少72.7%.
2.2种子扩散
至调查结束时,未间伐样地中仅有0.3% (2/600) 的种子原地存留,在间伐样地中有50.0% (300/600)的种子原地存留.山杏种子在未间伐样地和间伐样地的半存留时间分别为2 d和10 d,差异极显著(χ2=302.570,df=1,P<0.001) (图1).
2.3种子命运
在未间伐样地(0.3±0.3)%的种子被原地存留,而在间伐样地为(50.0±22.3)%,二者差异显著(U=42.000,Z=-2.087,P<0.05).在未间伐样地(7.3±2.2)%的种子被鼠类取食,而在间伐样地为(3.0±1.7)%,二者差异显著 (U=32.000,Z=-2.605,P<0.01) (图2).在未间伐和间伐样地,分别有(1.0±0.4)%和(1.3±0.6)%的种子被扩散后弃置地表,二者差异不显著 (U=66.000,Z=-0.386,P>0.05) (图2).在未间伐和间伐样地,分别有(60.7±5.1)%和(31.7±15.6)%的种子被鼠类分散贮藏,二者差异不显著 (U=48.000,Z=-1.402,P>0.05) (图2).
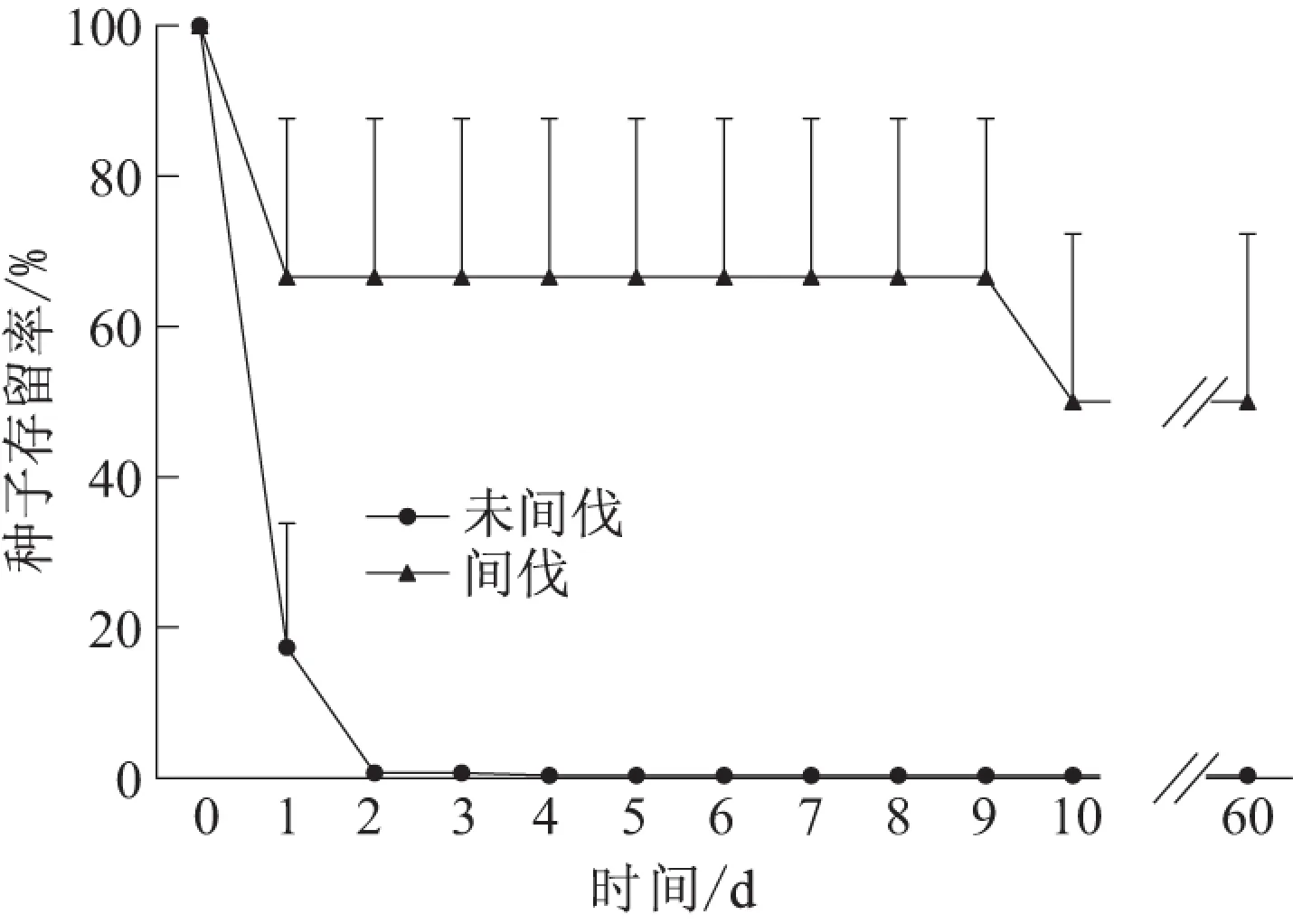
图1 山杏种子的存留动态Fig.1 The survival dynamics of wild apricot seeds
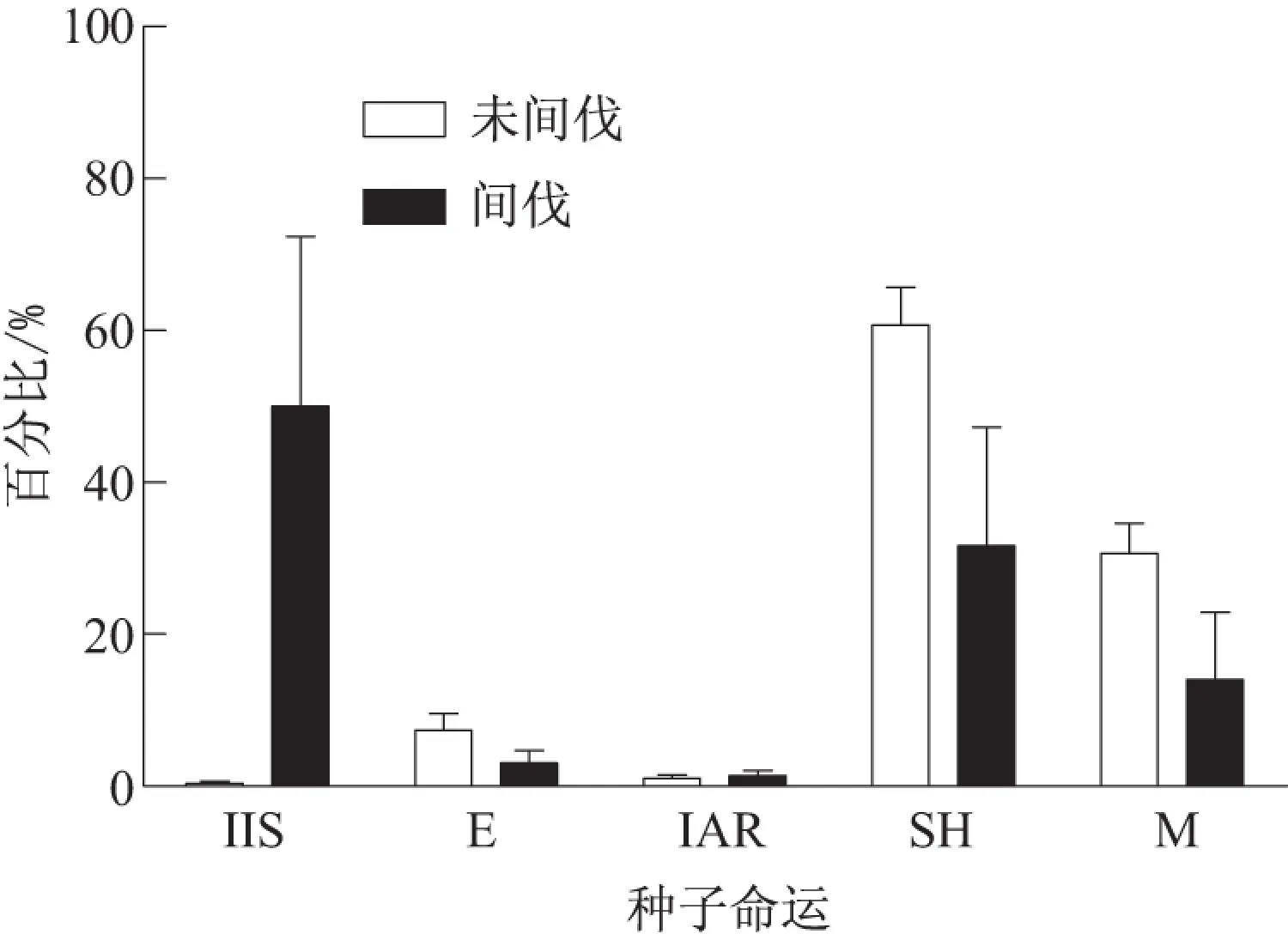
IIS: 原地存留; E: 取食; IAR: 弃置地表;SH: 分散贮藏; M: 丢失图2 两类样地内山杏种子的命运Fig.2 Fates of wild apricot seeds in un-thinning and thinning forest
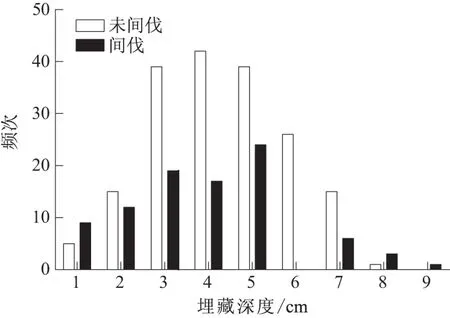
图3 间伐和未间伐样地内山杏种子埋藏深度的差异Fig.3 The difference of cache depth by rodents in un-thinning and thinning forest
2.4埋藏深度
在未间伐样地和间伐样地,鼠类分散贮藏种子的深度均小于9 cm,且两种样地内均有65.9%的种子被埋藏在3~5 cm深处 (图3).在未间伐样地内,鼠类对种子的平均埋藏深度为(4.3±0.1) cm (n=364),而在间伐样地内,平均埋藏深度为(3.9±0.2) cm (n=190),二者差异极显著 (t=2.050,df=271,P<0.01).
2.5种子的多次扩散
未间伐样地内种子被多次扩散的比例 (20.3%,n=74) 显著大于间伐样地内 (4.4%,n=8) (χ2=10.667,df=1,P<0.01).
3讨论
3.1间伐对植被和鼠类相对密度的影响
有研究认为,间伐不会影响森林中植物和鼠类的物种多样性[20].但也有研究认为,间伐作为一种人为干扰,能造成林内的植物多样性和数量明显下降[6—7],进而降低林栖动物食物资源的丰富度[28],最终导致林栖鼠类种类和密度的下降[6—7].本研究表明,间伐能导致植被和鼠类的种类减少、数量下降.鼠类数量的下降可能是由于植被密度下降[7, 16]和间伐过程中产生的干扰导致鼠类迁移.
3.2间伐对种子扩散和种子命运的影响
一些研究表明,森林间伐会减少种子扩散[22, 29],而另一些研究则表明,间伐有利于增加种子扩散[6, 20].本研究发现,山杏种子在未间伐样地扩散较快,而在间伐样地扩散较慢,其原因可能是间伐引起鼠类的种类和密度显著降低,而鼠类的种类和数量在很大程度上决定着种子的扩散速率[30—31];也可能与间伐样地中乔木和灌丛的盖度降低有关,因为盖度降低往往意味着捕食风险的增加[13, 32],而鼠类扩散种子的速率与植被盖度成正相关[13, 33].
本研究表明,在间伐样地,鼠类对种子的取食和分散贮藏均少于未间伐样地,与以往的研究结果一致[14].可能是由鼠类的种类发生变化造成的[4, 34],而不同的鼠种通常表现出不同的贮食特征[35—36];也可能是由于间伐后样地植被盖度的降低导致捕食风险增加[13],进而对鼠类取食和贮食行为产生不利影响[37—38].
分散贮藏种子的动物通常将种子浅埋于基质中,适当深度的埋藏能为种子萌发提供有利条件[1],不同的动物对种子的埋藏深度不同,鸟类埋藏种子的深度通常在0.5~10 cm[39],鼠类埋藏种子的深度在0~8 cm[40—41].本研究表明,鼠类埋藏山杏种子的深度为0~9 cm,主要集中在3~5 cm,而这一深度通常更有利于种子的萌发和幼苗建成[42];而鼠类在未间伐林区内埋藏种子的深度比间伐林区深,可能与间伐林区内落叶层和腐殖质层较薄有关,因为埋藏深度通常和贮藏点基质密切相关[43],也可能与间伐后林区内捕食风险增加有关.
鼠类对种子的多次扩散有助于将种子扩散至更远的地方,从而有利于植物幼苗的建成和种群的扩张[1, 25],而鼠类对种子的多次扩散受多种因素的影响,如种间种内竞争[44]、季节[45]、生境[46]等.本研究表明,未间伐样地内鼠类对分散贮藏种子的多次扩散率远高于间伐样地,可能与间伐样地内鼠类密度下降和捕食风险增加有关.
综上所述,间伐能导致林栖鼠类的种类与密度减少,并对鼠类的贮食行为产生影响,进而导致种子的扩散模式和命运发生改变,这可能对森林的幼苗建成和更新产生负面影响.
致谢:本研究得到了愚公林场刘金栋场长及职工的大力支持,在此表示诚挚的感谢;本文成稿过程中,数据分析得到了郑州大学生物多样性与生态学研究所王振龙教授的帮助,在此感谢.
参考文献:
[1]VANDER WALL S B. Food hoarding in animals [M]. Chicago: University of Chicago Press, 1990.
[2]FORGET P M, LAMBERT J, HULME P E, et al. Seed fate: seed predation, seed dispersal and seedling establishment [M]. Wallingford: CAB International Publishing, 2005.
[3]LAI X, GUO C, XIAO Z S. Trait-mediated seed predation, dispersal and survival among frugivore-dispersed plants in a fragmented subtropical forest, Southwest China [J]. Integrative zoology, 2013, 9(3): 246—254.
[4]CHANG G, ZHANG Z B. Differences in hoarding behaviors among six sympatric rodent species on seeds of oil tea (Camelliaoleifera) in Southwest China [J]. Acta oecologica, 2011, 37(3): 165—169.
[5]DIRZO R, DOMINGUEZ C A. Seed shadows, seed predation and the advantages of dispersal [M]. Dordrecht: Junk publishers, 1986: 237—249.
[6]FARVIG N, BÖHNING-GAESE K, BLEHER B. Enhanced seed dispersal ofPrunusafricanain fragmented and disturbed forests? [J]. Oecologia, 2006, 147(2): 238—252.
[7]KIRIKA J, FARWIG N, BÖHNING G K. Effects of local disturbance of tropical forests on frugivores and seed removal of a small-seeded Afrotropical tree [J]. Conservation biology, 2008, 22(2): 318—328.
[8]刘滨,王嵘,刘映良,等. 不同干扰强度生境中啮齿动物对苦槠种子的取食和扩散 [J]. 生态学杂志, 2011, 30(8): 1668—1673.
[9]ZHANG H M, WANG Y, ZHANG Z B. Domestic goat grazing disturbance enhances tree seed removal and caching by small rodents in a warm-temperate deciduous forest in China [J]. Wildlife research, 2009, 36(7): 610—616.
[10]ZHANG Y F, TONG L, JI W H, et al. Comparison of food hoarding of two sympatric rodent species under interspecific competition [J]. Behavioural processes, 2013, 92(3): 60—64.
[11]BAUDOIN C, HAIM A, DURAND J L. Effect of conspecific and heterospecific urine odors on the foraging behavior of the golden spiny mouse [J]. Integrative zoology, 2013, 8(suppl 1): 1—8.
[12]MENG L Z, GAO X, CHEN J, et al. Spatial and temporal effects on seed dispersal and seed predation ofMusaacuminatain southern Yunnan, China [J]. Integrative zoology, 2012, 7(1): 30—40.
[13]PEREA R, GONZLEZ R, SAN M A, et al. Moonlight and shelter cause differential seed selection and removal by rodents [J]. Animal behaviour, 2011, 82(4): 717—723.
[14]王利清,武晓东,付和平,等. 人为不同干扰方式下荒漠啮齿动物群落格局年间变动趋势 [J]. 干旱区资源与环境, 2011, 25(3): 146—151.
[15]张福顺,付和平,武晓东,等. 不同干扰下荒漠啮齿动物种群动态的空间同步性 [J]. 兽类学报, 2011, 31(2): 211—218.
[16]MARKL J S, SCHLEUNING M, FORGET P M, et al. Meta-analysis of the effects of human disturbance on seed dispersal by animals [J]. Conservation biology, 2012, 26(6): 1072—1081.
[17]陈凤娟,谷建才,黄冬梅,等. 不同间伐强度对华北落叶松人工林林下植物多样性的影响研究 [J]. 林业科学, 2008, 24(11): 169—173.
[18]SMITH D M. The practice of silviculture [M]. Eighth edition. New York: John Wiley & Sons Press, 1986.
[19]POULSEN J R, CLARK C J, BOLKER B M. Decoupling the effects of logging and hunting on an Afrotropical animal community [J]. Ecological applications, 2011, 21(5): 1819—1836.
[20]SCHLEUNING M, FARWIG N, PETERS M K, et al. Forest fragmentation and selective logging have inconsistent effects on multiple animal-mediated ecosystem processes in a tropical forest [J]. PloS one, 2011, 6(11): e27785.
[21]MORRIS R J. Anthropogenic impacts on tropical forest biodiversity: a network structure and ecosystem functioning perspective [J]. Philosophical transactions of the royal society B: biological sciences, 2010, 365(1558): 3709—3718.
[22]GUTIÉRREZ-GRANADOS G. Effect of logging on rodent scatter-hoarding dynamics in tropical forests: implications for plant recruitment [J]. Integrative zoology, 2011, 6(2): 74—80.
[23]路纪琪,王振龙. 河南啮齿动物区系与生态 [M]. 郑州: 郑州大学出版社, 2012.
[24]马庆亮,赵雪峰,孙明洋,等. 啮齿动物作用下退耕地山杏种子扩散与贮藏的季节变化 [J]. 应用生态学报, 2010, 21(5): 1238—1243.
[25]CAO L, XIAO Z S, GUO C, et al. Scatter-hoarding rodents as secondary seed dispersers of a frugivore-dispersed treeScleropyrumwallichianumin a defaunated Xishuangbanna tropical forest, China [J]. Integrative zoology, 2011, 6(3): 227—234.
[26]张知彬,王福生. 鼠类对山杏(Prunusarmeniaca)种子扩散及存活作用研究 [J]. 生态学报, 2001, 21(5): 839—845.
[27]XIAO Z S, JANSEN P A, ZHANG Z S. Using seed-tagging methods for assessing post-dispersal seed fate in rodent-dispersed trees [J]. Forest ecology and management, 2006, 223(1): 18—23.
[28]LAMBERT J E. Primate seed dispersers as umbrella species: a case study from Kibale National Park, Uganda, with implications for Afrotropical forest conservation [J]. American journal of primatology, 2011, 73(1): 9—24.
[29]HOLBROOK K M, LOISELLE B A. Dispersal in a Neotropical tree,Virolaflexuosa(Myristicaceae): does hunting of large vertebrates limit seed removal [J]. Ecology, 2009, 90(6): 1449—1455.
[30]LI H J, ZHANG Z B. Effects of mast seeding and rodent abundance on seed predation and dispersal by rodents inPrunusarmeniaca(Rosaceae) [J]. Forest ecology and management, 2007, 242(2/3): 511—517.
[31]DEMATTIA E A, CURRAN L M, RATHCKE B J. Effects of small rodents and large mammals on Neotropical seeds [J]. Ecology, 2004, 85(8): 2161—2170.
[32]LIMA S L, DILL L M. Behavioral decisions made under the risk of predation: a review and prospectus [J]. Canadian journal of zoology, 1990, 68(4): 619—640.
[33]WADA N. Dwarf bamboos affect the regeneration of zoochorous trees by providing habitats to acorn-feeding rodents [J]. Oecologia, 1993, 94(3): 403—407.
[34]LU J Q, ZHANG Z B. Differentiation in seed hoarding among three sympatric rodent species in a warm temperate forest [J]. Integrative zoology, 2008, 3(2): 134—142.
[35]VIEIRA E M, PIZO M A, IZAR P. Fruit and seed exploitation by small rodents of the Brazilian Atlantic forest [J]. Mammalia, 2003, 67(4): 533—540.
[36]TONG L, ZHANG Y F, WANG Z L, et al. Influence of intra-and inter-specific competitions on food hoarding behaviour of buff-breasted rat (Rattusflavipectus) [J]. Ethology ecology & evolution, 2012, 24(1): 62—73.
[37]LEAVER L A. Effects of food value, predation risk, and pilferage on the caching decisions ofDipodomysmerriami[J]. Behavioral ecology, 2004, 15(5): 729—734.
[38]SIVY K J, OSTOJA S M, SCHUPP E W, et al. Effects of rodent species, seed species, and predator cues on seed fate [J]. Acta oecologica, 2011, 37(4): 321—328.
[39]TOMBACK D F. Foraging strategies of Clark’s Nutcracker [J]. Living bird, 1978, 16: 123—160.
[40]VANDER WALL S B. Seed fate pathways of antelope bitterbrush: dispersal by seed-caching yellow pine chipmunks [J]. Ecology, 1994, 75(7): 1911—1926.
[41]VANDER WALL S B. Sequential patterns of scatter hoarding by yellow pine chipmunks (Tamiasamoenus) [J]. American midland naturalist, 1995, 133(2): 312—321.
[42]GUO C R, WANG Z L, LU J Q. Seed germination and seedling development ofPrunusarmeniacaunder different burial depths in soil [J]. Journal of forestry research, 2010, 21(4): 492—496.
[43]鲁长虎,袁力. 食干果鸟对种子传播的作用 [J]. 生态学杂志, 1997, 16(5): 43—46.
[44]CLARKE M F, KRAMER D L. The placement, recovery, and loss of scatter hoards by eastern chipmunks, Tamias striatus [J]. Behavioral ecology, 1994, 5(4): 353—361.
[45]常罡,邰发道. 季节变化对锐齿栎种子扩散的影响 [J]. 生态学杂志, 2011, 30(1): 189—192.
[46]肖治术,张知彬,路纪琪,等. 啮齿动物对植物种子的多次贮藏 [J]. 动物学杂志, 2004, 39(2): 94—99.
(责任编辑:王浩毅)
Effects of Thinning on Seed Dispersal by Rodents in Secondary Forest
ZHANG Yifeng1,WANG Weirui1,LI Wei2,MIAO Xiangdong3,LU Jiqi1
(1.SchoolofLifeScience,ZhengzhouUniversity,Zhengzhou450001,China; 2.CollegeofBasicMedicalSciences,ZhengzhouUniversity,Zhengzhou450001,China; 3.State-ownedYugongForestFarm,Jiyuan454650,China)
Abstract:As a common management strategy in forestry production, thinning could have a significant impact and interference on phytocoenosium and forest animals, and subsequently affect animal-mediated seed dispersal and forest regeneration. To understand the potential influence of thinning on the process of rodent-mediated seed dispersal, a study was conducted in April 2012. Seeds of Prunus armeniaca were marked and tracked in un-thinning and thinning forest in Mt. Wangwu area. The results showed that: 1) the relative density of rodent population drastically reduced from 6.7% in un-thinning forest to 2.7% in thinning forest; 2) the density of tree and shrub in thinning forest decreases from 56.3% to 72.7%; 3) seed dispersal rate in un-thinning forest (half survival time: 2 days) was significantly higher than in thinning forest (half survival time: 10 days); 4) the difference of burying depth between un-thinning forest (4.3±0.1)cm and thinning forest (3.9±0.2)cm was significant; 5) more scatter-hoarded seeds were retrieved and buried again in un-thinning forest (20.3%) than in thinning forest (4.4%). These results suggested that thinning have inevitable influence on plant community structure and the seed dispersal behavior of rodents, and finally affect the seed fate and seedling recruitment.
Key words:thinning; rodents; hoarding behavior; seed fate; burying depth
收稿日期:2015-10-01
基金项目:国家重点基础研究发展计划项目(2007CB109106).
作者简介:张义锋 (1984—),男,河南伊川人,博士,主要从事生态学、动物行为学研究,E-mail: zhangyf138@163.com;通讯作者,路纪琪 (1964—),男,陕西蒲城人,教授,博士,主要从事生态学、保护生物学研究,E-mail: lujq@zzu.edu.cn.
中图分类号:Q958.1
文献标志码:A
文章编号:1671-6841(2016)01-0067-06
DOI:10.3969/j.issn/1671-6841.201510001
引用本文:张义锋, 王魏瑞, 李蔚,等.间伐对鼠类扩散林木种子的影响[J] .郑州大学学报(理学版),2016,48(1):67—72
