大别山区毛叶山桐子的群落特征及其健康状况评估
2016-06-07周静波邱燕宁赵子睿唐燕平徐莹梅
周静波,邱燕宁,赵子睿,唐燕平,徐莹梅
山桐子(Idesia polycarpa)属大风子科山桐子属,为落叶乔木[1]。其适应性强,果实产量和含油率高,油料富含维生素 E、亚油酸等,营养价值高[2],工业用途广,市场需求很大,是优良的木本油料树种[3-4]。此外,其树形美观,树干通直,果实成串下挂,秋季红艳夺目,也是理想的观赏树种[2]。野生的山桐子主要分布在中国,朝鲜,日本及俄罗斯远东地区,在中国主要分布于秦岭淮河以南各省,在安徽省大别山地区有零星分布,为其变种毛叶山桐子(I.polycarpa var.vestita)。近 20 年来,人们对于山桐子的研究与开发表现出了浓厚的兴趣[5]。国内外已有一些对山桐子生物及生态学特性[6-10],种子和果实化学成分[11-12]及山桐子培育[13-17]等方面的研究,但在野生群落特征方面,研究较少。谢双喜等[18]对贵州省野生山桐子群落进行了研究。而山桐子健康状况影响因子方面,国内外暂无报道。本研究调查的目的是了解大别山地区山桐子的分布情况,研究野生毛叶山桐子群落特征及对成株毛叶山桐子的健康状况进行评估,为野生毛叶山桐子资源的合理开发和利用提供依据。
1 调查方法
1.1 调查地区概况
调查区域在安徽省大别山地区。大别山位于中国安徽、湖北、河南3省交界处,横跨鄂豫晥3省,呈东南—西北走向,长270 km,是长江流域和淮河流域的分水岭。
大别山属北亚热带温暖湿润季风气候区,具有典型的山地气候特征,气候温和,雨量充沛,雨热同季,具有优越的山地气候和森林小气候特征,具备森林的气候优势。大别山森林海拔差异大,植被变化明显,海拔从400~1 700 m,形成了丰富多彩的森林景观。
1.2 山桐子群落调查
采用样方法。2014-08—2014-10,选择了8处有毛叶山桐子分布的代表性地段做样方,其中3处位于安庆市岳西县鹞落坪自然保护区(样方号1~3),5处位于六安市舒城万佛山(样方号4~8)。有7处样方为成株样方(样方号1~2,4~8),1处为幼苗样方(样方号3),其中1处成株样方兼有幼苗(样方号8)。通过群落中不同物种重要值的计算来确认优势种,从而确定群落类型(图1,图2)。


图1 大别山区毛叶山桐子群落特征研究及成株毛叶山桐子健康状况评估Fig.1 Investigation on community characters of Idesia polycarpa var.vestita and evaluation on health condition of its Adult in the Dabie mountain area of China

图2 大别山区毛叶山桐子群落特征研究及成株山桐子健康状况评估Fig.2 Investigation on community characters of Idesia polycarpa var.vestita and evaluation on health condition of its adult in the Dabie mountain area of China
成株样方大小为10 m×10 m的乔木样方,幼苗为5 m×5 m的灌丛样方。目的物种乔木的测定参数包括树高、胸径、冠幅、地理坐标、健康状况。健康状况方面,从健康枝条/枯枝比例,病虫害程度,以及繁殖情况(本次调查期为山桐子花果期)几方面对山桐子健康状态进行了评估。目的物种幼苗的测定参数包括测定均高、多度、地理坐标和盖度。样方参数有坡位、坡向、坡度、海拔、土壤类型及乔木、灌木、草本层的优势种,并请当地专家对人为干扰强度进行判断(由无至强分4等,标0、1、2、3),并调查了乔木(幼苗灌丛没有此项)、灌木、草本及层间植物的种类、数目、高度和盖度等数据。其中,植物种类按照恩格勒1964年的系统,以文献[19]为标准,进行分类和鉴定。
1.3 统计分析
所有数据均采用SPSS 20.0软件进行相关性分析,计算样方内各物种的重要值,并计算各样方的丰富度指数,Simpson指数,Shannon-Wiener指数和Pielou均匀度指数。依据所采得的环境因子数据是否具有连续正态分布的性质,分别通过Pearson相关分析和Spearman秩相关分析,分析各环境因子对群落特征的影响,(海拔、坡度使用Pearson相关性分析,坡向、坡位、人为干扰使用Spearman秩相关性分析)以及用Spearman秩相关分析,分析各环境因子对山桐子健康情况的影响。由于山桐子分布及其健康状况对环境因子响应情况复杂,因子间存在相互作用,而本次调查仅选取了其中部分相对独立的环境因子数据,故统计检验时,当p<0.3时,研究者即认为其具有相关性[20]。
群落中各数量指标均按文献[21]计算,所有数据统计均利用 Excel 2011和SPSS 20.0完成。计算方法如下。
重要值=相对密度+相对优势度+相对频度;
密度=个体数/样地面积;
相对密度=(1个种的密度/所有种的密度)×100;
相对显著度=(1个种的显著度/所有种的显著度和)×100;
频度=该种出现的样方数/样方总数;
相对频度=(1个种的频度/所有种的频度和)×100;
α多样性指数包括丰富度,Simpson指数,Shannon-Wiener指数和Pielou均匀度指数,其计算方法见下。
物种丰富度指数S,即某群落所有样方中出现的物种总数。

式中:Ni为种i所在样地的重要值,N为种i所在样地的各个种的重要值之和,Pi为Ni与N的比值,S为种i所在样方的物种数。
2 结果与分析
2.1 山桐子群落特征及物种多样性分析
依据调查,山桐子在大别山出现的群落有雷公鹅耳枥(Carpinus viminea)群落,白辛树-盐肤木(Pterostyrax psilophyllus-Rhus chinensis)群落,毛叶山桐子(Idesia polycarpa var.vestita)幼苗群落,香果树-杉木(Emmenopterys henry-Cunninghamia lanceolata)群落,黄山松 -杉木(Pinus taiwanensis-Cunninghamia lanceolata)群落,黄山松(Pinus taiwanensis)群落,毛叶山桐子(Idesia polycarpa var.vestita)群落和毛叶山桐子-银鹊树(Idesia polycarpa var.vestita-Tapiscia sinensis)群落,只有 2 处成株样方,1处幼苗样方山桐子为优势种,其余样方山桐子均为伴生种。
各群落物种多样性指数见表1。各群落类型中,丰富度最高的群落是毛叶山桐子群落,为37,丰富度依次排列为:毛叶山桐子群落>毛叶山桐子-银鹊树群落>雷公鹅耳枥群落=黄山松群落=山桐子幼苗群落>白辛树-盐肤木群落>香果树-杉木群落>黄山松-杉木群落。
Simpson指数依次排列为:黄山松群落>黄山松-杉木群落>毛叶山桐子群落>山桐子幼苗群落>白辛树-盐肤木群落>香果树-杉木群落=雷公鹅耳枥群落>毛叶山桐子-银鹊树群落。
Shannon-Wiener指数依次排列为黄山松群落>毛叶山桐子群落>黄山松-杉木群落>毛叶山桐子-银鹊树群落>香果树-杉木群落>山桐子幼苗群落>雷公鹅耳枥群落>白辛树-盐肤木群落。
Pielou均匀度指数依次排列为:黄山松-杉木群落>黄山松群落>毛叶山桐子群落>山桐子幼苗群落>雷公鹅耳枥群落>白辛树-盐肤木群落>香果树-杉木群落>毛叶山桐子-银鹊树群落。
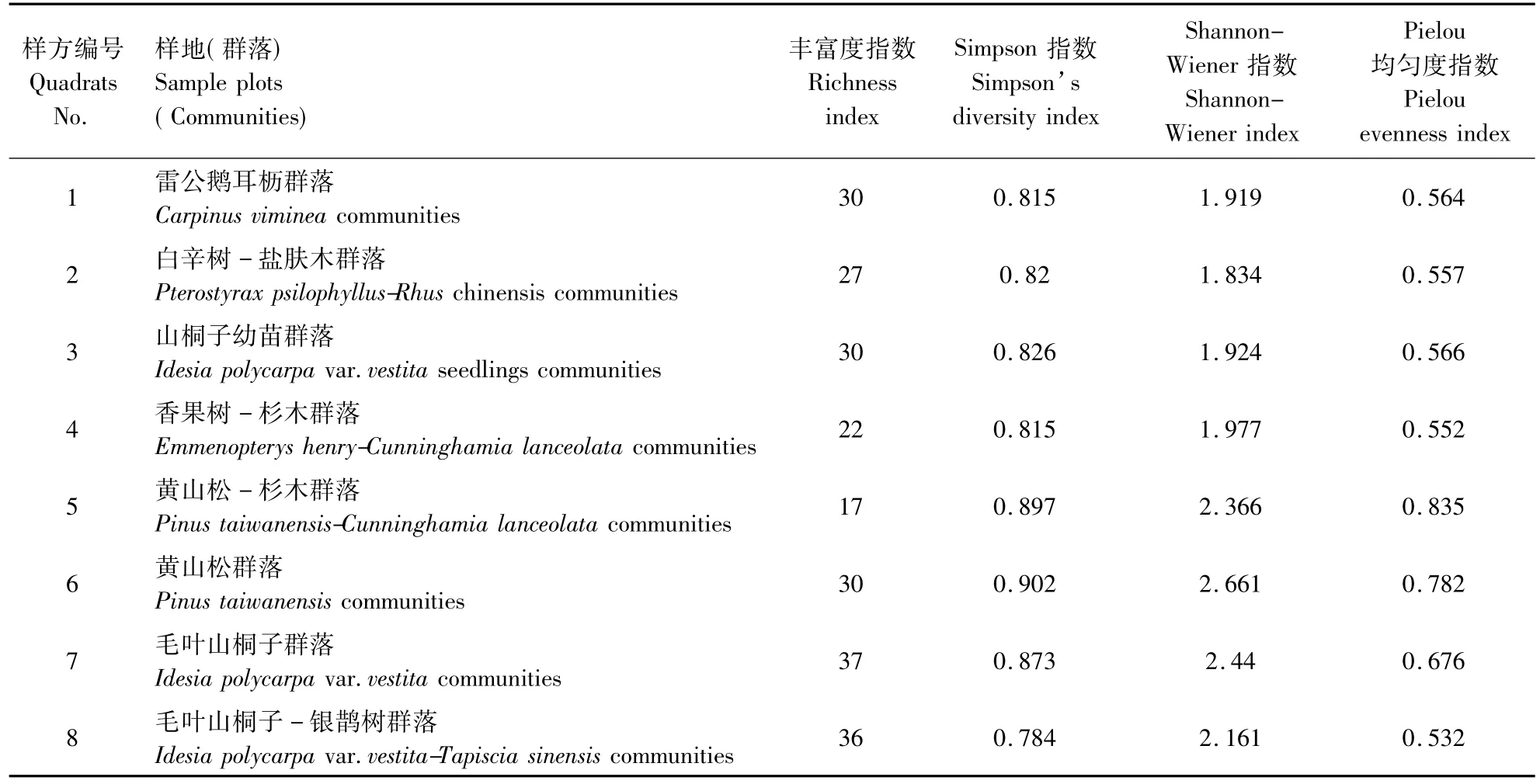
表1 不同群落物种α多样性指数Table 1 α-diversity indices of the investigated communities
由表1可知,各个样方群落的不同多样性指数变化趋势并不趋于一致,Simpson指数,Shannon-Wiener指数和Pielou均匀度指数一致性较好,而丰富度与其他的3个多样性指数具有一定的差异性。因为山桐子在所有群落中不是优势种,有些样方中仅为1株。群落丰富度反映了整个群落的生物多样性和生境优越性程度,对山桐子伴生的阔叶林相比,其伴生的针叶林或针叶-阔叶混交林的丰富度相对较小。但从多样性指数看,针叶林或针叶-阔叶混交林的多样性指数较大,这是由于多样性指数与分布格局有关,说明这类群落内的物种分布较均匀。
利用Pearson相关性分析和Spearman秩相关性分析群落多样性的环境梯度变化对群落多样性的影响,Spearman分析时,将坡向编号数字化,为北坡=1,东坡=2,南坡=3,西坡 =4,坡位编秩数字化,为谷底 =0,下坡=1,中坡=2,上坡 =3。结果见表2。

表2 多样性指数和环境因子相关系数(P值)Table 2 Correlation between diversity indices and the environmental factors(P Value)
由表2可见,坡向和坡位均显著影响了丰富度指数,Simpson指数和Pielou均匀度指数,人为干扰与丰富度指数显著负相关,而海拔和坡度与群落多样性指数相关性不显著。
2.2 山桐子健康状况及其影响因子分析
山桐子的长势、病虫害和繁殖力等健康状况一定程度上会影响其在群落中的功能。在大别山区,山桐子是群落的伴生种,大多数为1或2株孤立生长。野外实地考察,山桐子凡是生长在沟边土壤深厚且坡度小的地方,长势很旺盛,叶色浓郁,无枯枝和病虫害。对7块样方(除3号幼苗样方)内的11株山桐子成株健康状况及环境因子进行相关性分析的结果见表3。
由表3可见,山桐子成株的健康状况与坡度呈显著负相关,而与坡向、坡位、海拔和人为干扰等因子相关性不显著,说明其对海拔和坡向等因子的适应性相对较强。凡是位于自然保护区内的样地,人为干扰小;而在其他地区的样地,因人为活动频繁对山桐子生境造成较大的影响。

表3 山桐子成株健康状态和环境因子相关系数(P值)Table 3 Correlation between health condition of adult Idesia polycarpa var.vestita and environmental factors(P Value)
3 结论与讨论
本研究通过多次调查并对物种重要值的计算,首次探明了毛叶山桐子在大别山地区的分布,发现大别山地区毛叶山桐子在野外群落中较少以优势种出现,多以伴生种出现在针叶阔叶混交林和落叶阔叶林中。这与谢双喜等[14]的调查结果是基本一致的。
各个样方群落的不同多样性指数变化趋势并不趋于一致,Simpson指数,Shannon-Wiener指数和Pielou均匀度指数一致性较好,而丰富度与其他的3个多样性指数具有一定的差异性。例如,黄山松-杉木群落的丰富度指数与其他3个多样性指数差别很大,推测是因为黄山松-杉木群落位于万佛寺附近,群落周边有较多建筑物以及人流,并有人为清理植被的情况,因而丰富度指数较低,物种较少。而由于人为清理植被可能并不具有明确的倾向性,故其群落的均匀性较好。
毛叶山桐子群落多样性受多种因素影响。群落多样性与环境因子的相关性分析表明群落的多样性系数与坡位、坡向和人为干扰相关性显著,而与海拔、坡度相关性不显著。因为坡向在多种方面影响了植被类型和群落多样性。南北坡向差异造成了较大的光照强度的差异,从而导致了植被类型的差异。在北半球,南坡的光照强度大于北坡[17]。如本次调查中,北坡样方多为针阔混交林群落,而南坡样方则为落叶阔叶林群落。同时,坡位对群落生态因子的影响主要体现在温度上,谷底夜晚会形成冷湖,而山腰则相对温暖[22]。较高的温度可以给植物提供更好的生存环境,因此,山体下部和中部的群落多样性高于谷底。
毛叶山桐子成株健康状况与环境因子的相关性分析表明,毛叶山桐子的健康状况与坡度呈显著负相关,与其他环境因子相关性不显著。在调查中发现,几乎所有的健康成株都生长在沟边,土壤和水分因子对其影响较大。
毛叶山桐子喜阳生环境[1],适合于在山地生长。但坡度对群落多样性的影响主要体现在植物根系的生长,过于陡峭的山坡或悬崖植物扎根难度大,且雨水冲刷易流失不易渗透,滑坡风险大等,对植物生长有着较大的负面影响。所以,坡度在一定程度上也影响了毛叶山桐子成株的健康状况。
过度的人为干扰会影响群落多样性[22],本次调查的群落位于大别山腹地,环境条件并不十分优越,过多的人为干扰对群落多样性产生了一定程度的负影响。
本研究探明了毛叶山桐子在大别山地区的分布及其群落特征和健康状况影响因素,可为该地区的毛叶山桐子的种植和保护提供一定的科学依据。
[1] 刘根林,梁珍海,蒋泽平.山桐子研究综述[J].江苏林业科技,2005,32(5):46 -49.
[2] 罗韧.适宜重庆地区生长的新树种介绍[J].重庆林业科技,2003,65(4):57.
[3] 李秀全,徐有明.我国主要木本油料树种资源开发与林业生物能源林建设的探讨[J].生物质化学工程,2006,40(12):229 -234.
[4] 杨志玲,王开良,谭梓峰.值得开发的几种野生木本油料树种[J].林业科技开发,2003,17(2):41-43.
[5] 吴宗兴.山桐子开发与利用研究[J].四川林业科技,2010,31(1):26 -29.
[6] SETO M,YUI S.The amount of dissolved organic carbon in the soil solutions of a forest and a farm soil in situ[J].Japanese Journal of Ecology,1983,33(3):305-312.
[7] HIRABUKI Y.Phenology of woody plants in a fir forest in Sendai,northeast Japan [J].Japanese Journal of E-cology,1984,34(2):235 -238.
[8] KIKUCHI T,MIURA O.Differentiation in vegetation related to micro scale landforms with special reference to the lower side slope [J].Ecological Review,1991,22(2):61-70.
[9] SHIMADA K.Topographical distribution of five pioneer tree species and significance of their tree forms in natural forests on Mt.Takao,central Japan [J].Japanese Journal of Ecology,1994,44(3):293 -304.
[10] SAKAI A,SAKAI S,AKIYAMA F.Do sprouting tree species on erosion prone sites carry large reserves of resources[J].Annals of Botany,1997,79(6):625 -630.
[11]杨幼林,何立莹.毛叶山桐子果实不同发育期含油率测定简报[J].四川林业科技,1989,10(4):74 -75.
[12]周燕,周真明,曹沛,等.水冬瓜果肉的非油脂化学成分[J].天然产物研究与开发,2003,15(1):13 -14,17.
[13]程维泽.山桐子育苗及大苗培育技术[J].林业科技开发,2003,17(5):65 -65.
[14]祝志勇,季永华,沈定夫,等.山桐子育种苗期试验初报[J].江苏林业科技,2001,28(2):628.
[15]刘震,王玲.不同种源山桐子冬芽休眠的温度特性[J].河南农业大学学报,2000,34(3):252 -254.
[16]蒋泽平,梁珍海,刘根林,等.山桐子茎段离体培养技术研究[J].中国农学通报,2006,22(12):393-396.
[17]沈羊城,侯金艳,刘文博,等.山桐子种子快速高效萌发研究[J].中国农学通报,2015,31(13):5 -9.
[18]谢双喜,吴志文.贵州野生山桐子群落特征研究[J].天津农业科学,2013,19(1):94 -100.
[19]中国科学院《中国植物志》编委会.中国植物志[M].北京:科学出版社,2004.
[20] JIM C Y,W Y CHEN.Habitat effect on vegetation ecology and occurrence on urban masonry walls[J].Urban Forestry and Urban Greening,2010,9(3):169 -178.
[21] LEGENDRE P,LEGENDRE L.Numerical ecology[M].3rd ed.Amsterdam:Elsevier,2012.
[22]姜汉桥.植物生态学[M].第2版.北京:高等教育出版社,2010.
