高温胁迫对杧果幼苗生理生化指标的影响
2016-05-30CANVANTOAN罗聪何新华董龙DOMINHPHU
CAN VAN TOAN 罗聪 何新华 董龙 DO MINH PHU
摘 要 通过测定高温胁迫下四季杧与台农一号杧一年生嫁接苗叶片生理生化指标的变化,探讨其耐热的生理生化机制。结果表明: 随着高温胁迫时间的延长,四季杧和台农一号杧的丙二醛(MDA)含量和相对电导率均明显上升,但四季杧的上升幅度和含量均明显高于台农一号杧。保护酶活性方面,高温胁迫处理后四季杧的SOD酶先下降后上升,而台农一号杧则是先上升后下降,且其含量明显高于四季杧; CAT酶的含量在高温胁迫后2个品种均减少,但四季杧CAT酶的含量明显高于台农一号杧。渗透调节物质游离脯氨酸在高温胁迫处理后含量明显高于处理前,但台农一号杧的上升程度明显高于四季杧。可溶性糖的含量在处理后12 h时,四季杧的含量明显高于台农一号杧,但在处理后72 h时,结果却相反。可溶性蛋白含量先下降后上升,并且四季杧的上升程度明显高于台农一号杧。胁迫处理后2个杧果品种的叶绿素含量和叶片相对含水量均下降,但四季杧的含量均明显低于台农一号杧。综合各项指标,初步推测台农一号杧比四季杧更耐热。
关键词 杧果;高温胁迫;生理生化分析
中图分类号 S667.7 文献标识码 A
Abstract To investigate the physiological and biochemical mechanism for heat resistance of mango, two Mangifera indica L cv. ‘Siji and ‘Tainong No.1 were analyzed under high temperature stress. The results showed that MDA content and relative electrical conductivity both increased heat after treatment, but MDA contents and relative electrical conductivity were higher in ‘Siji than ‘Tainong No.1”. SOD activities of‘Sijifirst declined and then increased, but the enzyme activities were contrary in‘Tainong No.1and higher than ‘Sijicultivar. CAT activities decreased in both cultivars, but the activities were higher than‘Sijicultivar. The free proline contents increased both in the two cultivars, but the contents rising degree in‘Tainong No.1was higher than ‘Sijicultivar. The soluble sugar contents were higher in‘Sijicultivar than‘Tainong No.1after treated 12 h, but the results were opposite after treated 72 h. The soluble protein contents of both cultivars first declined and then increased, but the rising degree in ‘Sijicultivar was higher than‘Tainong No.1. The chlorophyll and relative water contents decreased in the two cultivars, but the contents were lower in ‘Sijicultivar than ‘Tainong No.1. Comprehensive comparative analysis showed that‘Tainong No.1was much more heat resistant than ‘Siji cultivar.
Key words Mango;High temperature stress;Physiological index
doi 10.3969/j.issn.1000-2561.2016.01.010
杧果(Mangifera indica Linn)屬于漆树科杧果属常绿大乔木植物,是世界五大热带水果之一。杧果原产于东南亚地区,中国也是杧果的原产地之一,在中国的云南、广西等地拥有丰富的野生杧果种质资源[1-2]。中国是杧果主要生产国之一,拥有悠久的种植历史,主要分布在海南、广西、广东、云南、四川和福建等地。近年来由于全球气候的不断变化,导致非生物胁迫时常发生。杧果对非生物胁迫比较敏感,特别是花期低温阴雨,严重影响杧果的授粉受精,坐果后初期的高温天气则易导致杧果果实发育畸形。
目前对杧果在干旱胁迫、低温胁迫和盐胁迫下的表现情况已经有报道[3-5],但尚未见对高温胁迫下杧果生理生化指标方面的相关报道。本研究以一年生盆栽四季杧与台农一号杧嫁接苗为材料,进行高温处理,采集不同时间样品,比较分析其生理生化指标,从而为杧果苗期抗热性鉴定以及探索杧果对高温的影响机理提供理论和实验依据。
1 材料与方法
1.1 材料
长势基本一致的一年生盆栽(40 cm×50 cm)四季杧和台农一号杧(均为嫁接苗,砧木为土杧)。杧果苗由广西大学农学院标本园供应。
1.2 方法
1.2.1 样品采集 将长势基本一致的一年生盆栽四季杧与台农一号杧放入人工气候室进行44 ℃高温处理,光照强度为3 000 lx,选取不同方向的叶片进行样品采集,采样的时间分别为0(CK)、2、4、8、12、24、48、72 h,分别测定丙二醛、相对电导率、SOD酶、CAT酶、游离脯氨酸、可溶性糖、可溶性蛋白、叶绿素和相对含水量。每个处理3棵苗,重复3次。
1.2.2 测定方法 根据《植物生理学实验指导》[6],用巴比妥酸(TBA)显色法测丙二醛(MDA)含量、用电导仪率法测电导率、用氮蓝四唑(NBT)显色法测过氧化物歧化酶(SOD)、采用紫外分光光度计法测量过氧化氢酶(CAT)和游离脯氨酸含量、用茚酮法测定可溶性糖含量、用考马斯亮蓝G-250染色法测定可容性蛋白质含量、采用叶绿素测定仪ECA-051测定叶绿素、用浸泡法测定相对含水量。
1.3 数据处理
采用Excel 2010、DPS7.0软件完成试验数据统计,采用新复极差法进行显著性分析。
2 结果与分析
2.1 高温胁迫对杧果幼苗叶片中膜透性的影响
2.1.1 MDA的测定 MDA含量可以直接反映植物膜脂过氧化的水平和对细胞膜的伤害程度。由图1-a可知,常温下四季杧的MDA含量高于台农一号杧,高温处理后MDA含量随着处理时间的延长而增加,但四季杧的MDA含量和升高倍数明显高于台农一号杧,四季杧处理2 h后,其MDA含量就极显著高于处理前,12 h时,MAD含量极速上升,72 h时MDA含量达到处理前的3.16倍;而台农一号杧处理2 h时的含量与处理前变化不明显,4 h时其含量极显著高于处理前,而后MAD含量缓慢上升,48 h时达到高峰是处理前的2.52倍。
2.1.2 质膜透性的变化 相对电导率是膜透性大小的指标,同一胁迫处理下,其变化水平反映植物生物膜受伤害的程度以及植物抗逆性的大小,数值越大,质膜受伤害越大。由如图1-b可知,处理前2个杧果品种的相对电导率差异不明显,处理后2 h,四季杧相对电导率变化不明显,4 h时其相对电导率极显著高于处理前,而后四季杧相对电导率随处理时间的延长而明显上升,72 h时达到高峰,是处理前的4.74倍;台农一号杧处理后2 h,相對电导率极显著高于处理前,而后有所下降,相对电导率随处理时间的延长阶梯式上升,72 h时达到高峰,是处理前的2.66倍。
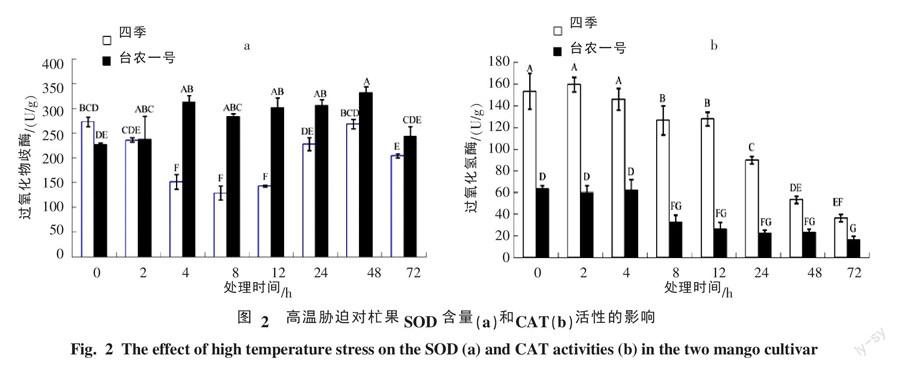
2.2 高度胁迫对杧果幼苗叶片中保护酶活性的影响
2.2.1 SOD酶活性的变化 由图2-a可知,四季杧在高温处理0~8 h后,随着处理时间的延长,其SOD含量逐渐下降,8 h时达到最低值,极显著低于处理前水平,比处理前降低了52.76%,随后SOD含量开始上升,48 h时恢复到处理前水平,而后又开始下降;台农一号杧在高温处理后含量开始上升,4~48 h保持较高的水平,SOD含量均极显著高于处理前水平,72 h时含量又开始下降,但其SOD含量也比处理前高;处理前四季杧的SOD含量极显著高于台农一号杧,但4~48 h时四季杧的SOD含量极显著低于台农一号杧。
2.2.2 CAT 酶活性的变化 由图2-b可知,CAT酶活性的大小与植物代谢强度以及抗性有密切的关系[7]。高温处理0~4 h,2个杧果品种CAT酶的活性与处理前变化不显著,4 h后2个杧果品种的CAT含量均出现极显著下降,72 h时四季杧和台农一号杧的CAT含量比处理前分别下降了76.12%和73.67%,但四季杧的CAT酶活性均极显著高于高于台农一号杧。
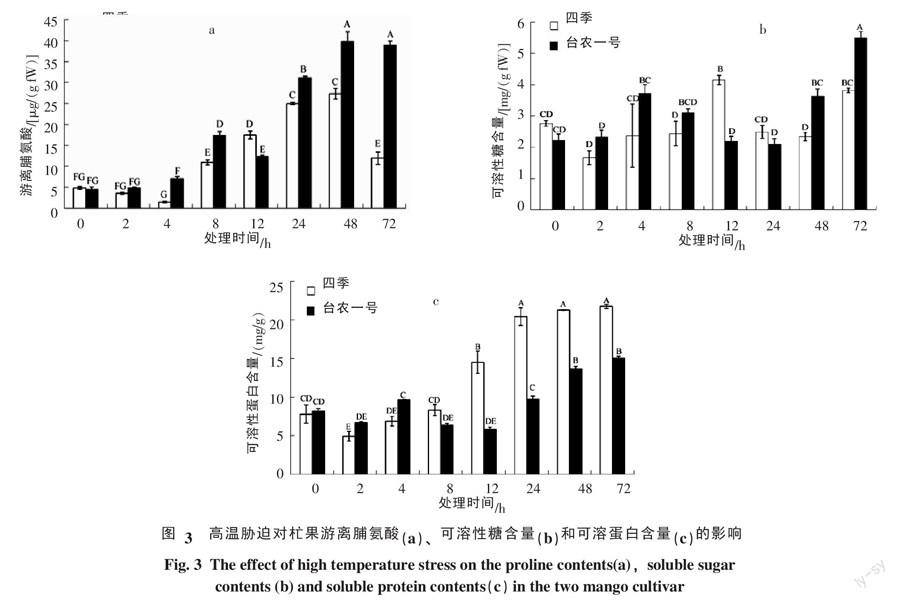
2.3 高温胁迫对杧果幼苗叶片渗透调节物质的影响
2.3.1 游离脯氨酸变化 由图3-a可知,高温处理后4 h,四季杧的游离脯氨酸含量达最低值,比处理前下降了38.7%,极显著低于处理前水平,而后期含量开始逐步上升,48 h时脯氨酸含量出现最高值,比处理前提高了5.68倍,而后下降,但8~72 h其脯氨酸含量均极显著高于处理前;台农一号杧的游离脯氨酸含量在高温处理后表现为先上升,8 h时出现第一个小高峰,比处理前高1.84倍,12 h时有所下降,然后又开始上升,48 h时脯氨酸含量出现最高值,比处理前提高了9.97倍,而后又有所下降;游离脯氨酸含量均高于处理前,8~72 h其脯氨酸含量均极显著高于处理前;四季杧的游离脯氨酸含量在处理前和处理12 h时略高于台农一号杧,2、4、8、24、48和72 h游离脯氨酸含量均低于台农一号杧。
2.3.2 可溶性糖含量变化 由图3-b可知,高温胁迫后2 h时,四季杧的可溶性糖含量下降到最低,是处理前的60.4%,而后开始上升,12 h时含量达到高峰,是处理前的1.5倍,而后又开始下降,24和48 h时可溶性糖含量与处理前差异不显著,72 h时可溶性糖含量略高于处理前水平;台农一号杧在处理后2 h时,其可溶性糖含量与处理前变化不明显,4 h时其可溶性糖含量极显著上升,而后又开始下降,24 h时降低到处理前水平,之后开始逐渐上升,72 h时达到最高值,是处理前的2.47倍;在高温处理前四季杧的可溶性糖含量略高于处理前水平,但差异不显著,12 h时四季杧的可溶性糖含量显著高于台农一号杧, 48和72 h时台农一号杧的可溶性糖含量显著高于四季杧,其他时间2个品种差异不显著。
2.3.3 可溶性蛋白变化 由图3-c可知,高温胁迫后,四季杧的可溶性蛋白呈先下降后上升的趋势,2 h时出现最低值,之后开始逐步上升,72 h时达到最高峰,是处理前的2.79倍;台农一号杧的可溶性蛋白含量呈先下降后上升再下降再上升的趋势,12 h时最低,72 h时出现最高峰,是处理前的1.83倍;高温胁迫后12~72 h四季杧的可溶性蛋白上升幅度极显著高于台农一号杧。
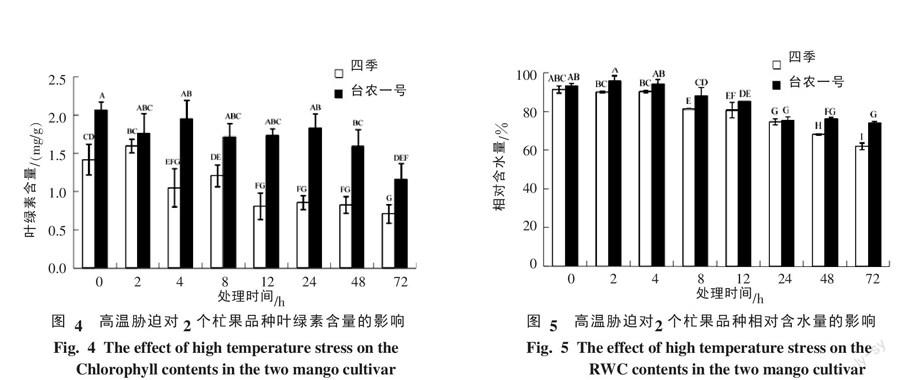
2.4 高温胁迫下杧果种叶片中叶绿素含量的变化
由图4可知,高温胁迫后,四季杧叶片中的叶绿素含量呈现先稍微上升而后下降再上升再下降的趋势,在处理后72 h时下降到最低水平,其含量只有处理前的34.4%;台农一号杧叶片中的叶绿素含量出现先下降而后又上升又下降再上升再下降的过程,但高温胁迫处理后,其叶绿素含量均低于处理前水平;高温胁迫处理72 h时出现最低值,其含量只有处理前的60.6%;0~72 h时台农一号杧的叶绿素含量均明显高于四季杧。
2.5 高温胁迫下杧果叶片中相对含水量的变化
由图5可知,高温胁迫0~4 h时,2个杧果品种叶片中的相对含水量变化不明显,8 h时叶片中的相对含水量均出现持续下降,72 h时两者的相对含水量都达到最低值,其中四季杧的相对含水量比处理前降低了32.15%,而台农一号杧只降了20.76%;处理前2个杧果品种叶片中的相对含水量差异不明显,而在处理后2、8、48、72 h时,四季杧的相对含水量极显著低于台农一号杧。
3 讨论与结论
丙二醛是由于植物器官衰老或者在逆境条件下受伤害,其组织或器官膜脂质发生过氧化反应而产生的,其含量与植物衰老及逆境伤害有密切关系[8]。膏桐幼苗在45 ℃高温胁迫下,MDA和相对电导率含量随着胁迫时间的延长而升高[9]。大花三色堇幼苗在35 ℃高温处理后电导率与MDA含量均上升[10]。有研究结果表明, 耐热品种的电导率上升明显小于不耐热品种,耐热性强的植物在高温和正常条件下叶片MDA含量均比不抗热性植物叶片中的含量少,且温度越高这种趋势越明显[11]。本研究结果表明,随着高温胁迫时间延长,2个杧果品种叶片中的MDA与相对电导率而逐渐上升,这结果跟前人研究结果一致。四季杧叶片中的MDA含量和电导率上升幅度均极显著大于台农一号杧,这说明在高温胁迫下,四季杧细胞膜受破坏比台农一号杧更厉害。
高温胁迫初期,植物为了适应环境条件会提高自身抗氧化酶活性来清除活性氧,这是植物的保护性应激反应[12-13]。但随着高温胁迫程度加强,植物自身的抗氧化酶逐渐减少,使酶活性下降,导致植物活性氧的积累[14-15]。植物体在长期进化过程中形成了完善和复杂的自身抗氧化保护酶系统来清除活性氧,其中SOD和APX是主要抗氧化保护酶类[16]。受高温胁迫时,抗热和不抗热的大花三色堇幼苗的CAT活性下降,SOD活性显著上升,而且抗热品种的SOD活性均高于不抗热品种[10]。在高溫胁迫下,月李耐热品种的SOD活性高于不耐热品种[8]。金露梅幼苗在高温胁迫下,SOD活性出现下降,CAT活性先上升后下降趋势[17]。本研究结果表明,高温处理后,四季杧叶片中的SOD均低于处理前,而台农一号杧SOD含量明显高于处理前的水平。而高温胁迫下2个杧果品种的CAT酶活性均明显下降,但四季杧的CAT酶活性下降幅度高于台农一号杧。说明台农一号杧相对于四季杧在高温胁迫处理后其保护酶的保护能力更强。
游离脯氨酸、可溶性糖和可溶性蛋白含量的变化反映植物细胞的渗透调节能力。耶兴元等[18]认为脯氨酸是一种活性氧清除剂,其对植物细胞有保护作用,在逆境胁迫下抗逆强的植物游离脯氨酸含量积累比抗逆弱的植物多。含糖量高的株系,表现出较强的耐热性[19-20]。随着高温胁迫程度增强游离脯氨酸和可溶性蛋白不断积累,但胁迫温度过高会导致含量降低。金露梅可通过增强细胞中的可溶性糖和游离脯氨酸渗透调节物质来抵御高温胁迫[17]。本研究结果表明,高温胁迫下2个杧果叶片中的脯氨酸含量均出现不同程度的上升,并极显著高于处理前水平。而可溶性糖含量和可溶性蛋白含量也出现上升,这一结果跟前人的结果一致[17-18, 21]。高温胁迫下台农一号杧的游离脯氨酸含量和可溶性糖含量积累高于四季杧,因此台农一号杧的耐热能力强于四季杧。
叶绿素含量常用来鉴定植物的抗热性[14, 22]。温度的升高能诱导植物体内叶绿素分解使其含量明显下降[19, 23-24]。周中亮等[25]研究结果表明,随着高温胁迫程度加强,6个高羊茅株系叶绿素含量先上升后下降,其原因是温度升高可以使植物光合作用增强,因此叶绿素含量上升,但持续的高温胁迫会造成叶绿体的变形和片层结构的破坏。而罗少波等[26]认为,高温胁迫后叶绿素含量升降没有一定规律性,不能作为大白菜耐热鉴定指标。本研究结果表明,高温胁迫初期,2个杧果品种幼苗叶片的叶绿素含量变化不明显,这可能由于胁迫时间短未达到抑制光合作用的程度,但随着胁迫时间延长,两者的叶绿素含量出现降低,在胁迫后期两者下降更加明显,其中四季杧的叶绿素含量下降幅度高于台农一号杧。Scafaro等[27]指出耐热品种的净光合速率下降幅度明显小于不耐热品种。说明台农一号杧耐热性比四季杧强。
叶片相对含水量(RWC)是植物水分状况的重要指标。不同品种的耐热能力跟细胞的相对含水量成正相关,耐热性高的品种细胞持水能力更强[10, 28]。本研究结果表明,随着高温胁迫时间延长,2个杧果品种幼苗叶片的RWC均出现下降现象,而四季杧下降幅度比台农一号杧大,这说明台农一号杧细胞持水能力比四季杧更强。
总之,高温胁迫下四季杧叶片中的MDA含量和相对电导率的上升幅度均显著高于台农一号杧,而SOD含量、脯氨酸含量、叶绿素含量和相对含水量则明显低于台农一号杧,说明以上指标是比较研究杧果品种之间耐热性高低的最佳指标,并且台农一号杧幼苗的耐热能力比四季杧强。
参考文献
[1] 雷新涛, 姚全胜, 徐雪荣, 等. 中国野生杧果种质资源及其AFLP分子标记[J]. 热带作物学报, 2009, 30(10): 1 408-1 412.
[2] 樊卫国, 罗 燕, 吴素芳,等. 南北盘江河谷野生杧果种质资源的分布与形态特征[J]. 西南农业学报, 2012, 25(6): 2 244-2 247.
[3] Sukhvibul N, Whiley A W, Smith M K. Effect of temperature on seed and fruit development in three mango(Mangifera indica L.)cultivars[J]. Sci Hortic, 2005, 105: 467-474.
[4] Nabil I, Cao K F. Gas exchange, chlorophyll?uorescence, and osmotic adjustment in two mango cultivars under drought stress[J]. Acta Physiol Plant, 2008, 30: 769-777.
[5] Gaelle D, Marc V, Laurent U. Long-term drought results in a reversible decline in photosynthetic capacity in mango leaves, not just a decrease in stomatal conductance[J]. Tree Physiol,2009, 29: 675-684.
[6] 周祖富, 黎兆安. 植物生理学实验指导[M]. 广西大学, 2005.
[7] 刘德良,赖万年. 夏季梅花品种耐热性生理生化指标研究[J]. 北方园艺, 2012(2): 57-61.
[8] 蒋昌华, 胡永红, 秦 俊,等. 高温胁迫对月季品种部分生理指标的影响研究[J]. 种子, 2008(6): 31-34.
[9] 吴 涛, 郎南军,彭明俊. 高温胁迫对膏桐幼苗部分生理指标的影响[J]. 东北林业大学学报, 2008(11): 29-31.
[10] 彭华婷, 高 悦, 杜红梅,等. 高温胁迫对大花三色堇幼苗相关生理指标的影响[J]. 上海交通大学学报(农业科学版), 2012 (6): 66-71.
[11] 项延军, 李新芝. 扶芳藤, 等. 3种藤本植物耐热性生理生化指标初探[J]. 江苏农业科学, 2011(3): 241-243.
[12] 段九菊, 王云山,康黎芳. 高温胁迫对观赏凤梨叶片抗氧化系统和渗透调节物质积累的影响[J]. 中国农学通报, 2010(8):164-169.
[13] 刘易超, 杨际双, 肖建忠,等. 高温胁迫对菊花叶片部分生理参数的影响[J]. 河北农业大学学报, 2011(6): 46-49.
[14] 黄显波, 严 寒, 胡建林,等. 高温胁迫下水稻幼苗几个相关生理指标的变化[J]. 长江大学学报(自然科学版)农学卷, 2008 (2): 50-53.
[15] 吴友根, 林尤奋, 李绍鹏,等. 热胁迫下菊花生理变化及其耐热性指标的确定[J]. 江苏农业学报, 2009(2): 362-365.
[16] 周 广, 孙宝腾, 张乐华,等. 井冈山杜鹃叶片抗氧化系统对高温胁迫的响应[J]. 西北植物学报, 2010(6): 1 149-1 156.
[17] 郭盈添, 范 琨, 白 果,等. 金露梅幼苗对高温胁迫的生理生化响应[J]. 西北植物学报, 2014(9): 1 815-1 820.
[18] 耶兴元, 马锋旺, 王顺才,等. 高温胁迫对猕猴桃幼苗叶片某些生理效应的影响[J]. 西北农林科技大学学报(自然科学版),2004(12): 33-37.
[19] Liu X Z, Huang B R. Heat stress injury in relation to membrane lipid peroxidation in creeping bentgrass[J]. Crop Sci,2000, 40(2): 503-510.
[20]张庆峰,徐 胜,李建龙. 高温胁迫下高羊茅生理生化特性研究[J]. 草业科学, 2006(4): 26-28.
[21] 马英姿, 张 慧, 宋 荣, 等. 高溫胁迫对蛇足石杉生理特性的影响[J]. 中草药, 2013(2): 224-228.
[22] 周伟辉, 薛大伟,张国平. 高温胁迫下水稻叶片的蛋白响应及其基因型和生育期差异[J]. 作物学报, 2011(5): 820-831.
[23] 王 庆, 刘安成, 张瑞博,等. 高温胁迫对多叶羽扇豆生长及生理指标的影响[J]. 北方园艺, 2013(24): 52-54.
[24] 黄闽敏, 张 强,刘晓芳. 高温胁迫对春兰耐热性相关生理指标的影响[J]. 山西农业科学, 2013(12): 1 330-1 332.
[25] 周中亮, 包满珠, 王文恩. 高温胁迫对6个高羊茅株系生理指标的影响[J]. 草业科学, 2011(7): 1 284-1 290.
[26] 罗少波, 李智军, 周微波,等. 大白菜品种耐热性的鉴定方法[J]. 中国蔬菜, 1996(2): 16-18.
[27] Scafaro A P, Haynes P A, Atwell B J. Physiological and molecular changes in Oryza meridionalis Ng.,a heat-tolerant species of wild rice[J]. J Exp Bot, 2010, 61(1): 191-202.
[28] 胡永红, 蒋昌华, 秦 俊, 等. 高温对月季部分形态、生理指标的影响研究[J]. 种子, 2008(7): 26-28.
责任编辑:古小玲
