氮素形态对油麦菜幼苗光合作用及其荧光特性的影响
2016-05-30胡海非张巧柔吴卫东王晋占丽英曹立钟凤林林义章
胡海非 张巧柔 吴卫东 王晋 占丽英 曹立 钟凤林 林义章

摘 要 采用水培技术,以两叶一心的油麦菜幼苗为材料,研究铵态氮和硝态氮不同配比对油麦菜幼苗叶片光合色素含量、光合作用和叶绿素荧光参数的影响。结果表明:(1)不同氮素处理对油麦菜幼苗叶片中叶绿素a、叶绿素b和类胡萝素的含量均产生显著影响,当NO3- ∶ NH4+=7 ∶ 3时叶绿素a、叶绿素b和类胡萝素的含量明显增加,显著高于其他处理。(2)各处理下油麦菜幼苗的净光合速率、气孔导度、蒸腾速率随处理时间的增长呈上升趋势,而胞间CO2浓度则呈下降趋势,当NO3- ∶ NH4+=7 ∶ 3时净光合速率、气孔导度和蒸腾速率显著高于其他处理,而胞间CO2浓度则顯著低于其他处理。(3)油麦菜幼苗叶片荧光特性呈现出较大差异,当硝态氮与铵态氮的配比为7 ∶ 3时,油麦菜幼苗叶片PSⅡ的Fv/Fm和Fv/F0最高;由相关性分析可知,Fv/Fm、Fv/F0与叶绿素a和类胡萝卜素的相关性高于叶绿素b;Fv/Fm和Fv/F0三元回归模型的决定系数均在0.9以上,准确反映出油麦菜幼苗叶片叶绿素荧光动力学参数与光合色素之间的关系。
关键词 油麦菜;氮素;光合作用;荧光特性
中图分类号 S636 文献标识码 A
Abstract The hydroponics technique was used to observe the influence of different mixing ratios of ammonium nitrogen and nitrate nitrogen on the photosynthetic pigment content, photosynthesis, and chlorophyll-fluorescence parameters of the leaf of lettuce seedlings. The result showed that:(1)The content of chlorophyll a, chlorophyll b and carofenoid of the leaf were all significantly affected by different nitrogen treatments. The content of chlorophyll a, chlorophyll b and carotenoid increased significantly higher than other treatments when the ratio of nitrate nitrogen and ammonium nitrogen was 7 ∶ 3.(2)Net photosynthetic rate, stomatal conductance and transpiration rate increased over time, while intercellular CO2 concentration decreased. Net photosynthesis, stomatal conductance and transpiration rate were significantly higher than other treatments, and intercellular CO2 was significantly lower than other treatments when the ratio of nitrate nitrogen and ammonium nitrogen was 7 ∶ 3.(3)The fluorescence characteristic showed obvious difference, the photochemical efficiency and potential activity in PSⅡwere the highest, when the ratio of nitrate nitrogen and ammonium nitrogen was 7 ∶ 3; The determination coefficient of PSⅡ photochemical efficiency and potential activity ternary linear regression model were both above 0.9, accurately showed the relationship between chlorophyll fluorescence kinetics parameters and photosynthetic pigments. This study lays the foundation for the further study of the influence of nitrogen on photosynthesis.
Key words Lactuca sativa L.; Nitrogen; Photosynthesis; Fluorescence characteristic
doi 10.3969/j.issn.1000-2561.2016.01.002
氮素是植物生长过程中的必须营养元素之一,是构成蛋白质、核酸、叶绿素、酶、维生素等重要有机化合物的主要成分[1]。然而,植物对氮素的吸收和利用不仅取决于自身的生理特性,还与其生长环境中所提供的氮源密切相关[2]。NH4+-N与NO3--N是植物吸收氮素的两种主要形态[3],植物对不同形态的氮素营养因植物种类、器官和发育阶段的不同而做出不同的反应。
光合作用是指绿色植物利用光能,将CO2和H2O造成有机物并释放出氧气的过程,是作物产量形成的基础。大量研究表明,氮素形态对小麦、玉米等植物的生长和光合作用有显著影响[4]。在单独供应NH4+-N时,植物叶片具有叶面积小[5]、叶片比面重较大[6]和叶绿体体积大[7]等特征。不同氮素形态对植物光合速率的影响因种类不同而有异。大量研究表明供应NH4+-N时叶片的光合速率比供应NO3--N时高[8],而Lenz[9]研究表明不同氮素形态对草莓叶片的光合速率没有显著影响。
油麦菜(Lactuca sativa L.)属菊科莴苣属,是以嫩梢、嫩叶为食用器官的叶用莴苣。油麦菜含有大量的维生素、胡萝素等营养成分,具有抗病性和适应性强、生长周期短等特点,是无土栽培研究的模式植物之一。近年来,很多国内外学者开展了氮素营养对油麦菜影响的研究,主要集中在栽培效果[10]、产量和品质[11]、硝酸盐积累[12]、油麦菜生长[13]等方面,关于氮素形态对油麦菜光合机理的影响尚不清晰,鉴于此,本研究通过设计不同氮素形态配比营养液栽培试验,分析油麦菜幼苗叶片光合色素含量、光合速率的变化规律,并探讨不同氮素形态下油麦菜幼苗叶片叶绿素荧光动力学参数的響应机制,为深入研究油麦菜氮素营养的调控机理及提高氮素营养的利用效率奠定理论基础。
1 材料与方法
1.1 材料
供试油麦菜品种为四季高产油麦菜。
1.2 方法
1.2.1 试验设计 试验于2014年10月在福建农林大学园艺学院温室中进行。试验在自主设计的层架式立体管道(3列4层)中进行水培。将植株用海绵固定悬挂在定植孔内,根系深入到营养液深层以吸收养分和水分。该装置营养液层高7cm,营养液量较多,不需经常补充和更换营养液,可以根据植物的大小合理控制水位来达到理想的种植效果。
营养液配备采用华南农业大学的叶菜B营养液配方[14],在此基础上设5个氮素配比处理,分别为NO3- ∶ NH4+=10 ∶ 0,NO3- ∶ NH4+=7 ∶ 3,NO3- ∶ NH4+=5 ∶ 5,NO3- ∶ NH4+=3 ∶ 7,NO3- ∶ NH4+=0 ∶ 10,依次记为P1、P2、P3、P4、P5。5个处理的营养液配方中的微量元素含量均为:微量元素(NH4)6Mo·4H2O 0.002 mg/L、H3BO3 2.86 mg/L、MnSO4·4H2O 2.13 mg/L、ZnSO4·7H2O 0.222 mg/L,CuSO4·5H2O 0.08 mg/L,NaFe-EDTA 20 mg/L。大量元素配方见表1。
2014年10月20日穴盘育苗,当苗长至2片真叶时定植于PVC管道,用全营养液栽培,定植初期,营养液管理为电导度0.8~1.0 mS/cm,栽培后期用不同氮素形态处理,并将电导度调至 1.2~1.8 mS/cm。营养液供给使用定时控制器控制,设定自动间歇循环供液,白天每隔 4 h 循环供液 30 min,夜间停止循环。
1.2.2 测定项目与方法
(1)油麦菜幼苗光合特性参数。采用CIRAS-3光合测定系统测定油麦菜幼苗的净光合速率(A)、气孔导度(Gs)、胞间CO2浓度(Ci)和蒸腾速率(E),每个处理3次重复。
(2)叶绿素荧光动力学参数。叶绿素荧光动力学参数采用便携式调制叶绿素荧光仪PAM-2500进行测定,叶片暗适应时间为20 min。选取不同处理相同部位的功能叶片进行指标测定,每个处理测试10株。通过测定和计算获取相应的荧光参数:初始荧光F0、最大荧光Fm、可变荧光Fv、PSⅡ的光化学效率Fv/Fm和潜在活性Fv/F0。
(3)叶片光合色素含量。在叶绿素荧光动力学参数采集完成后,将所测试的叶片摘下,装入自封袋,带入实验室用于光合色素含量分析,利用分光光度法测定其叶绿素a、叶绿素b和类胡萝卜素的含量。
1.3 数据处理
试验数据采用DPS的LSD多重比较进行差异显著性分析,显著水平为0.05。
2 结果与分析
2.1 不同氮素形态对油麦菜幼苗叶片光合色素含量的影响
2.1.1 叶绿素a 随着油麦菜幼苗的生长,功能叶片叶绿素a含量逐渐增加。不同氮素营养形态处理下叶片叶绿素a含量变化趋势基本相同,但从处理的第2天开始各处理均表现出显著差异。P2处理较其他处理能够显著提高叶绿素a含量,P3处理下油麦菜幼苗叶片叶绿素a含量的变化趋势与P2处理相同,但对叶绿素a形成的促进作用显著低于P2处理,P4处理的油麦菜幼苗叶片叶绿素a含量在处理的第8天达到最高,第10天稍有下降,P1处理对油麦菜幼苗叶绿素a含量的影响在处理的第6天才表现出显著差异(表2)。通过比较不同氮素营养形态处理下油麦菜幼苗叶片中叶绿素a含量的差异可以发现,P2处理对应的植株叶片中叶绿素a含量显著高于其他4个处理,在处理的第10天比P1、P3、P4和P5处理分别高出1.03、1.12、1.05和1.22倍。
2.1.2 叶绿素b 油麦菜幼苗功能叶片叶绿素b含量的变化趋势与叶绿素a相似,但叶绿素b含量较低,在处理的第2天开始各处理间表现出显著差异。P2处理下油麦菜幼苗叶绿素b含量显著高于其他处理,且在处理的第8天起叶绿素b含量迅速增长,到处理的第10天,P2处理的叶绿素b含量比P1、P3、P4和P5处理分别高出1.19、1.18、1.18和1.15倍(表2)。P3、P4对叶绿素b含量的影响无明显差异,略低于P2处理,P5处理的油麦菜幼苗叶绿素b含量一直处于最低水平。
2.1.3 类胡萝卜素 类胡萝卜素含量能够反应叶片的光呼吸作用能力。不同氮素营养形态处理下油麦菜幼苗叶片类胡萝卜素含量的变化趋势基本一致,从处理的第2天开始各处理间差异显著。P2处理对类胡萝卜素含量的影响与叶绿素b相似,在处理的第8天起迅速增加,在P1处理的第4天开始,油麦菜幼苗类胡萝素含量迅速增加, P3、P4 和P5处理下的类胡萝素含量缓慢上升(表2)。
2.1.4 叶绿素a/b值 由表2可知,除P5处理在处理的第6天叶绿素a/b值开始下降外,其他处理下的叶绿素a/b均一直呈下降趋势,到处理的第10天,P2处理的油麦菜幼苗叶片叶绿素a/b值比P1、P3、P4和P5处理分别降低0.95、0.87、0.94和0.92倍。
2.1.5 叶绿素a+b 在不同形态的氮素营养条件下,油麦菜幼苗叶片中叶绿素a、叶绿素b和类胡萝卜素的含量均存在显著差异,其中在处理的第2、8、10天表现得更为明显,不同形态的氮素处理对叶绿素a与叶绿素b含量比值的影响在处理的第2天表现出显著差异(表2)。通过比较不同处理油麦菜幼苗叶片中叶绿素总含量(叶绿素a+b)的差异可以发现,P2处理对应的植株叶片中叶绿素总含量要高于其他4个处理。
2.2 不同氮素形态对油麦菜幼苗叶片光合特性的影响
2.2.1 净光合速率 由图1-A可知,不同氮素营养形态处理的油麦菜幼苗叶片净光合速率的变化趋势基本一直。从不同施氮处理来看,随着混合氮素中比例的增加,油麦菜幼苗叶片的光合速率也逐渐升高,P2处理的显著优于其他氮素形态比例的处理,以处理的第8天为例,P2处理的油麦菜幼苗叶片净光合速率依次是P1、P3、P4、P5处理的1.3、1.13、1.37和1.55倍。
2.2.2 气孔导度 油麦菜幼苗叶片气孔导度随植株的生长不断升高。由图1-B可知,不同氮素营养形态对油麦菜幼苗气孔导度的影响差异显著。P2处理下的气孔导度在第8天时达到最高,比P1、P3、P4和P5处理分别高出1.12、1.11、1.04、1.09倍。P3处理气孔导度持续增长,略低于P2处理,P5处理下气孔导度的变化趋势与P2处理相似,但气孔导度一直保持在各处理的最低水平。
2.2.3 胞间CO2浓度 随油麦菜幼苗的生长,胞间CO2浓度呈下降趋势,各氮素营养形态处理下的胞间CO2浓度变化趋势基本一致,从处理的第2天开始各处理间差异显著。P2处理的胞间CO2浓度一直保持较低水平,在处理的第8天迅速下降。P1、P4、P5处理的胞间CO2浓度没有显著差异。P3处理的胞间CO2浓度略高于P2处理(图1-C)。
2.2.4 蒸腾速率 随油麦菜幼苗的生长,各处理下油麦菜幼苗叶片的蒸腾速率均呈上升趋势,差异显著。P1、P5处理的蒸腾速率均保持在所有处理的较低水平,且变化趋势基本相同。P2处理在处理的第4天蒸腾速率迅速上升,到第8天时,上升速率变慢,但依然保持上升趋势。P3处理下的油麦菜幼苗叶片的蒸腾速率呈平稳上升趋势。P4处理的油麦菜幼苗叶片的蒸腾速率在处理的第4~6天和8~10天呈快速上升趋势,上升速率高于P2,第6~8天基本没有上升(图1-D)。
2.3 不同氮素形态对油麦菜幼苗叶绿素荧光动力学参数的影响
2.3.1 初始荧光F0 不同氮素营养形态对油麦菜幼苗叶片的初始荧光F0有明显影响。P1和P2处理下的F0在处理时间内呈缓慢上升趋势,从处理的第4天开始P2处理显著高于P1处理的F0。P3处理在处理的第8天达到最大值,第10天保持不变,整体呈上升趋势,但在处理的第2天和第6天都有不同程度的下降。P4和P5处理下的F0在处理的第6天都有不同程度的下降,随后上升。在处理的第2天,不同氮素形态对油麦菜幼苗叶片初始荧光F0影响不大,各处理间的差异不显著,但在处理的第4天开始,P2、P3和P4处理的F0明显高于P1和P5处理(图2-A)。
2.3.2 可变荧光Fv 各处理的可变荧光Fv在处理进行到第4天时开始表现出差异。从处理的第2~8天各处理对应的油麦菜幼苗叶片可变荧光Fv均显著上升,但在处理的第10天,除P2处理外,其他几个处理对应的油麦菜幼苗叶片可变荧光Fv值均有稍微下降(图2-B)。在处理的第8天,P2处理较P1、P3、P4和P5处理分别高出1.06、1.02、1.05和1.07倍。
2.3.3 最大光化学效率Fv/Fm 由图2-C可知,油麦菜幼苗在不同氮素营养形态的处理下,PSⅡ最大光化学效率Fv/Fm呈上升趋势,变化趋势基本一致,但各处理间上升的程度不同,表现出差异性。具体表现为P2处理下的油麦菜幼苗的Fv/Fm在处理的第4天迅速上升,并一直保持在最高水平,P3处理较P2处理略低,其次是P4处理,再次为P1处理,而P5处理下的油麦菜幼苗的Fv/Fm没有明显变化。
2.3.4 潜在活性Fv/Fo 由图2-D可知,不同氮素营养形态处理下油麦菜幼苗PSⅡ潜在活性的变化趋势与Fv/Fo的变化趋势基本一致。除P5处理外,其余各处理均呈现持续上升的趋势,但各处理上升的程度不同,P2处理始终保持在各处理的最高水平,P5处理在处理的第6天稍有下降,随后缓慢上升。
2.4 叶绿素荧光动力学参数与油麦菜幼苗叶片光合色素含量相关性分析
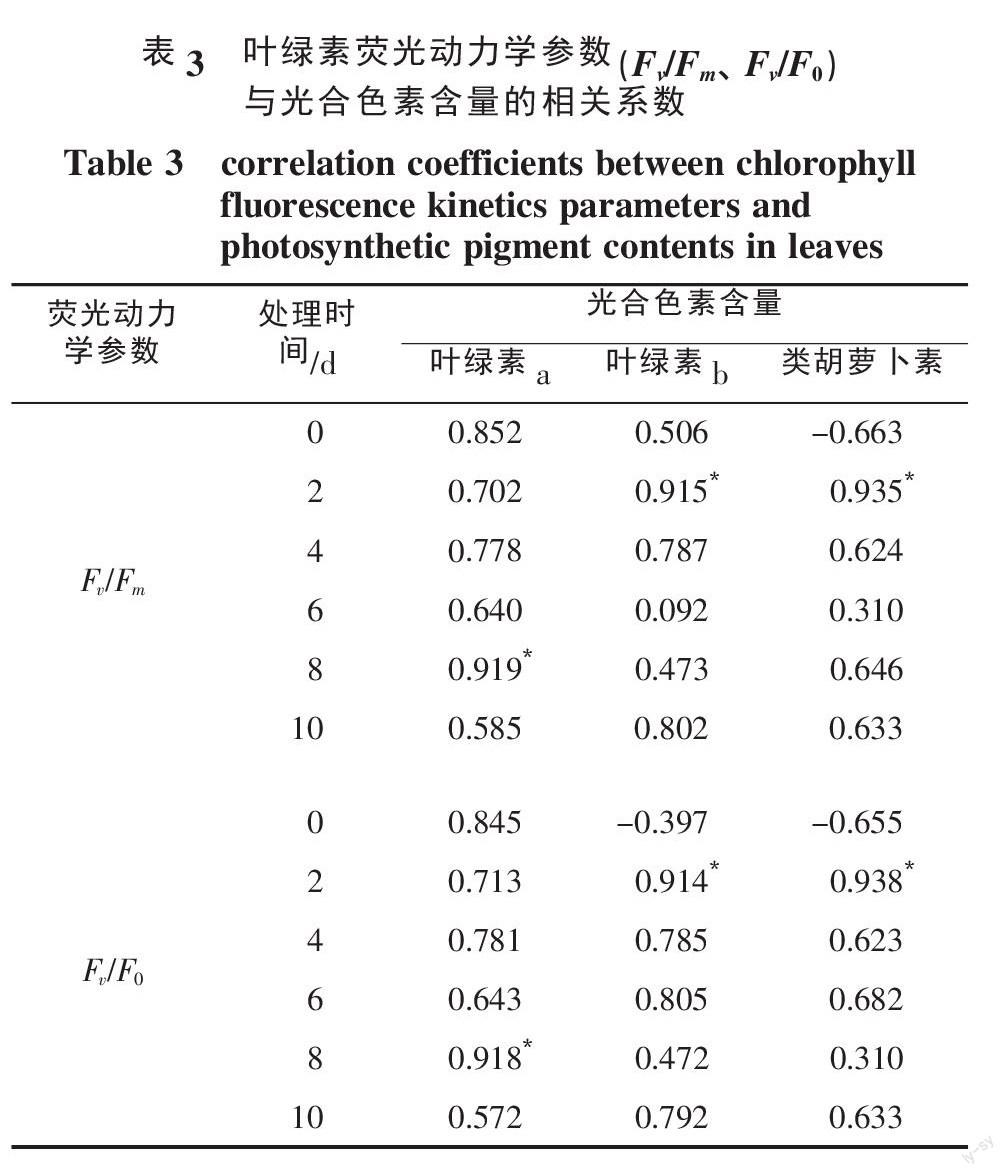
由表3叶绿素荧光动力学参数(Fv/Fm、Fv/F0)与油麦菜幼苗光合色素含量的相关性检验结果可以看出,PSⅡ的Fv/Fm与叶绿素a含量的相关系数在处理的第8天最高,达到0.919,与叶绿素b和类胡萝卜素含量的相关系数均在处理的第2天最高,分别达到0.915和0.935;PSⅡ的Fv/F0与叶绿素a含量的相关系数在处理的第8天最高,达到0.918,与叶绿素b和类胡萝卜素含量的相关系数均在处理的第2天最高,分别达到0.914和0.938。与叶绿素a和类胡萝卜相比,油麦菜幼苗叶片中叶绿素b含量与叶片叶绿素荧光动力学参数的相关系数略低。
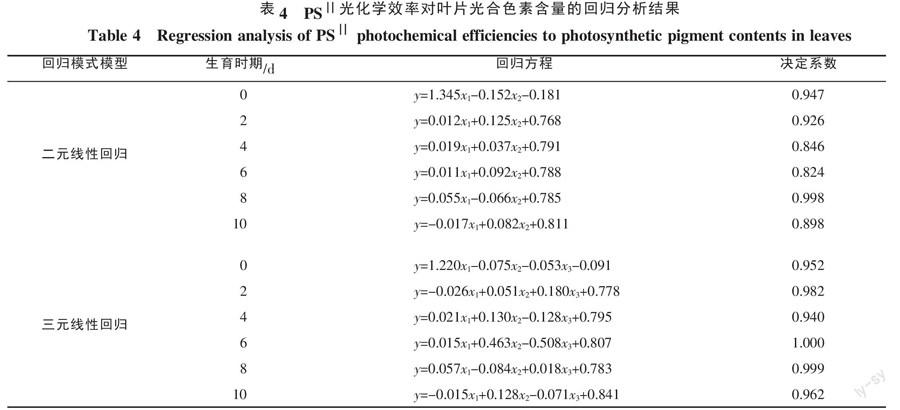
以不同处理时间所测试的油麦菜幼苗叶片作为样本,建立叶绿素荧光动力学参数关于油麦菜幼苗叶片光合色素含量的回归模型(表4、5),分析结果表明,Fv/Fm、Fv/F0三元线性回归模型的决定系数R2基本较二元线性回归模型的决定系数要高,因此,三元线性回归模型更适合用来描述油麦菜幼苗叶片叶绿素荧光动力学参数与光合色素含量之间的关系。
3 讨论与结论
光合作用是植物产量形成的基础,光合色素吸收、传递和转换光能,是植物进行光合作用的物质基础。光合作用除了受光照、温度等影响外,还受到矿质元素的影响。氮素对作物叶片光合速率、叶绿素以及气孔导度等均有明显影响,直接或间接影响植物的光合作用[15-16]。大量研究表明,氮素营养对植物叶片光合作用和荧光动力学特性具有显著影响。魏猛等[17]发现,每盆叶菜型甘薯施尿素6.5时其叶片叶绿素含量、光合速率和气孔导度均最高,有利于叶菜型甘薯光合作用和生物产量的形成;李晓静等[18]发现NO3--N对芫荽生长的促进作用优于NH4+-N,且NO3--N ∶ NH4+-N为3 ∶ 1时气孔导度增大,净光合速率为所有处理最大,说明氮素形态影响气孔导度,并通过气孔导度进而影响叶片的光合速率;伍松鹏等[19]发现,在总氮量供应相同的情况下,随营养液中硝态氮比例的增加,黄瓜幼苗叶绿素含量、光合速率、总生物量呈上升趋势,说明硝态氮更有利于黄瓜幼苗叶片的光合作用。本研究重点探讨了不同氮素形态比例对油麦菜幼苗叶片光合生理的影响,结果表明,不同氮素形态对油麦菜幼苗叶片中叶绿素a、叶绿素b和类胡萝卜素的合成都有显著影响,其中叶绿素a在处理的第6、8、10天表现的较为明显,类胡萝素素在处理的第4、6、8、10天表现的比较明显,而叶绿素b和叶绿素a/b在处理的第10天才表现出显著差异,这说明不同的氮素营养形态影响了油麦菜幼苗光合色素的合成与代谢,但对不同光合色素的合成与代谢影响不同。作物产量的形成大部分来自光合作用,光合速率能够直接反应光合作用强弱[20]。本研究结果表明,不同氮素营养形态对油麦菜幼苗的净光合速率的影响不同,当硝态氮与铵态氮的配比为7 ∶ 3时的净光合速率明显高于其他几个处理,不同氮素营养形态对气孔导度的影响和光合速率基本一致,说明硝态氮比铵态氮更有利于促进光合作用。叶绿素荧光可以快速灵敏的反应出植物光合系统光能利用效率等光合生理的变化[21]。本研究结果表明,氮素营养形态不同,油麦菜幼苗叶片荧光特性呈现出较大差异,通过分析叶片叶绿素荧光动力学参数的变化可知,不同氮素形态对油麦菜幼苗叶片光能吸收与转化活性及原初电子受体的光还原活性均产生了较大影响,其中P2和P3处理显著提高油麦菜幼苗叶片的PSⅡ光化学效率及潜在活性。当硝态氮与铵态氮的配比为7 ∶ 3时,油麦菜幼苗叶片PSⅡ的光化学效率和潜在活性最高,有利于提高油麥菜幼苗的光能利用效率,促进产量形成。此外,通过叶绿素荧光动力学参数与光合色素相关性分析可知,Fv/Fm、Fv/F0与叶绿素a和类胡萝卜素的相关性要高于叶绿素b。本研究还发现Fv/Fm和Fv/F0三元回归模型的决定系数均在0.9以上,能够准确的反映出油麦菜幼苗叶片叶绿素荧光动力学参数与光合色素之间的关系。研究结果为进一步探讨氮素对光合作用机理的影响奠定基础。
参考文献
[1] Cruz J L, Mosquim P R, Pelacani C R, et al. Photosynthesis impairment incassava leaves in response to nitrogen deficiency[J]. Plant and Soil, 2003, 257: 417-423.
[2] 乔建磊, 于海业, 宋述尧, 等. 氮素形态对马铃薯叶片光合色素及其荧光特性的影响[J]. 中国农业大学学报, 2013, 18(3): 39-44.
[3] 陈 巍, 罗金葵, 姜慧梅, 等. 不同形态氮素比例对不同小白菜品种生物量和硝酸盐含量的影响[J]. 土壤学报, 2004, 419(3): 421-425.
[4] Wiesler F. Agronomical and physiological aspects of ammonium and nutrition of plants[J]. Plant Nutri-tion and Soil Science, 1997, 160: 227-238.
[5] Walch-Liu P, Neumann G, Bangerth F, et al. Rapid effects of nitrogen form on leaf morphogenesis in tobacco[J]. Expermental Botany, 2000, 51: 227-237.
[6] Bowler J M, Press M C. Effect of elevated CO2, N form and concentration on growth and photosynthe-sis of a fast-and slow-growing grass[J]. New Phytologist, 1996, 132: 391-401.
[7] Raab T K, Terry N. Nutrition source regulation of growth and photosunthesis in Beta vulgaris L[J]. Plant Physiology, 1994, 105: 1 159-1 166.
[8] Guo S W, Schinner K, Sattelmacher B, et al. Different apparent CO2 compensation points in n-itrate and ammonium-growth Phaseolus vulgaris and the relationship to non-photorespiratory CO2 evolution[J]. Physiologia Plantarum, 2005, 123: 288-301.
[9] Claussen W, Lenz F. Effect of ammionium or nitrate nutrition on net photosynthesis, growth, and aciti-vity of the enzymes nitrate reductase and glutamine synthetase in blueberry, raspberry and strawberry[J]. Plant and Soil, 1999, 208: 95-102.
[10] 林俊芳, 林义章, 邱武昌, 等. 不同氮素水平对油麦菜栽培效果的综合评价分析[J]. 热带亚热带植物学报, 2013, 21(5): 432-438.
[11] 孙自航, 艾绍英, 刘春生, 等. 氮肥种类和用量对油麦菜生物和品质的效应[J]. 土壤肥料科学, 2007, 23(9): 366-371.
[12] 章君果, 达良俊, 张彩仙, 等. 氮磷水平对于油麦菜产量及其硝酸盐积累的影响[J]. 华东师范大学学报(自然科学版), 2010(2): 43-47.
[13] 宁建凤, 邹献中, 杨少海, 等. 有机无机氮肥配施对土壤氮淋失及油麦菜生长的影响[J]. 农业工程学报, 2007, 23(11): 95-100.
[14] 郭世荣. 无土栽培学[M]. 北京: 中国农业出版社, 2003: 111-116.
[15] 郑 洁, 胡美君, 郭延平. 光质对植物光合作用的调控及及其机理[J]. 应用生态学报, 2008, 19(7): 1 619-1 624.
[16] 徐济春, 林钊沐, 罗 微, 等. 矿质营养对光合作用影响的研究进展[J]. 安徽农学通报, 2007, 13(7): 23-25.
[17] 魏 猛, 唐忠厚, 陈晓光, 等. 不同氮素水平对叶菜型甘薯光合作用及生长特性的影响[J]. 江苏农业学报, 2014, 30(1): 87-91.
[18] 李晓静, 樊明寿. 氮素形态对芫荽光合作用和硝酸盐积累的影响[J]. 华北农学报, 2015(5): 221-223.
[19] 伍松鹏, 张秀娟, 吴 楚, 等. 不同氮素形态比例对黄瓜幼苗生长和光合特性的影响[J]. 安徽农业科学, 2006, 34(12): 2 697-2 707.
[20] 赵黎明, 李 明, 郑殿峰, 等. 水稻光合作用研究進展及其影响因素分析[J]. 北方水稻, 2014, 44(5): 67-71.
[21] 陈建明, 俞晓平, 程家安, 等. 叶绿素荧光动力学及其在植物逆境生理研究中的应用[J]. 浙江农业学报, 2006, 18(1): 51-55.
责任编辑:沈德发
