武夷岩茶加工过程茁-葡萄糖苷酶对香气形成的影响
2016-03-22邓慧莉周子维王秋铭范文静福建农林大学园艺学院福建福州350002
邓慧莉,周子维,王秋铭,范文静,孙 云(福建农林大学园艺学院,福建福州350002)
武夷岩茶加工过程茁-葡萄糖苷酶对香气形成的影响
邓慧莉,周子维,王秋铭,范文静,孙云
(福建农林大学园艺学院,福建福州350002)
摘要:通过分析武夷岩茶水仙和肉桂加工过程中茁-葡萄糖苷酶活性以及香气成分,探讨茁-葡萄糖苷酶对水仙和肉桂加工过程香气形成影响的差异性。结果表明,水仙香气含量和茁-葡萄糖苷酶活性低于肉桂,但加工过程香气和茁-葡萄糖苷酶活性变化率高于肉桂;水仙茁-葡萄糖苷酶活性与香气、橙花叔醇、香叶醇均呈高度指数负相关,相关系数分别为0.931、0.973、0.999,与芳樟醇、雪松醇分别呈高度、较高指数正相关,相关系数分别为0.947、0.872;肉桂茁-葡萄糖苷酶活性与香气、橙花叔醇、香叶醇、芳樟醇氧化物I呈较高指数负相关,相关系数分别为0.897、0.888、0.869、0.882。
关键词:茁-葡萄糖苷酶;香气;水仙;肉桂
乌龙茶是中国特有茶类,外形紧结壮实,色泽油亮,具有天然花果香气,其香气成分含量高,同时香气是影响乌龙茶品质的重要因子。
茶叶香气形成于三个部分[1],即鲜叶中固有的游离态香气、鲜叶在加工过程中由酶促作用产生的香气以及加工过程中经热物理化学变化所形成的香气。香气的形成主要有四种途径,其中最受关注的途径是糖苷类前体物质水解生成相应的醇类物质。在茶叶醇类糖苷水解途径[2]中,起重要作用的主要是茁-樱草糖苷酶、茁-葡萄糖苷酶以及茁-半乳糖苷酶[3,4],其中茁-葡萄糖苷酶和茁-樱草糖苷酶对乌龙茶花果香气的形成起关键作用。陈艺元[5]的研究表明晒青过程茁-葡萄糖苷酶活性升高,做青过程酶活性降低有利于乌龙茶香气总量、醇类香气增加。张娴静[6]研究铁观音加工过程茁-葡萄糖苷酶活性变化与香气形成的关系结果表明,晒青过程酶活性提高,香气总量、醇系香气、橙花叔醇含量也相应提高。
武夷岩茶在乌龙茶中的声名远扬。武夷山茶树品种丰硕,名丛浩繁,但武夷山主栽品种是水仙和肉桂等[7]。肉桂香高,水仙品质特征优良稳定,因而深受消费者喜爱[8]。
本试验通过对水仙和肉桂的茁-葡萄糖苷酶活性、香气组分和含量进行测定,分析水仙和肉桂香气以及茁-葡萄糖苷酶对香气形成的影响差异性等问题,通过从加工技术等环节入手,使武夷岩茶独特香气优势更加突显、品质稳定中有所突破。
1 材料与方法
1.1材料
本试验于2015年4-5月在武夷山市凯捷岩茶厂进行,以采自武夷山市上梅乡的春季水仙、肉桂品种为供试材料,鲜叶采摘标准为开面三四叶。
1.2茁-葡萄糖苷酶活性测定方法
1.2.1主要仪器设备
仪器设备:FA1104电子天平、台式高速冷冻离心机、722型可见光分光光度计、恒温水浴锅、电热鼓风干燥箱、微波炉、YDS-10B液氮罐(四川亚西低温设备有限公司)、IMS-70全自动雪花制冰机、超低温冰箱、超纯水器。
1.2.2原料加工方法
武夷岩茶制作工艺流程:鲜叶寅萎凋寅做青寅杀青寅揉捻寅干燥。
1.2.3取样方法
取样标准:选取原料相对一致的材料取第二叶5g样品,采用“液氮固样法”进行固样,试验样品编号及名称如表1。
1.2.4茁-葡萄糖苷酶活性测定
参照吴乔[9]的试验方法。
1.3香气测定方法
参照刘琳燕[10]的试验方法。
2 结果与分析
2.1水仙加工过程香气成分与茁-葡萄糖苷酶活性变化
2.1.1水仙加工过程香气成分分析
水仙加工过程香气总离子流图(以XY-S和GZH-S为例)如图1所示。

图1 水仙在制品香气总离子流图

表1 试验样品编号与名称
水仙加工过程中,香气成分总量呈上升趋势,如图2所示。其中1YQ-S至GZH-S的香气成分总峰面积值分别为XY-S的1. 5、1.94、4.69、5.75倍,1YQ-S至SQQ-S的涨幅最为明显,GZH-S的香气成分总峰面积达到最高值。
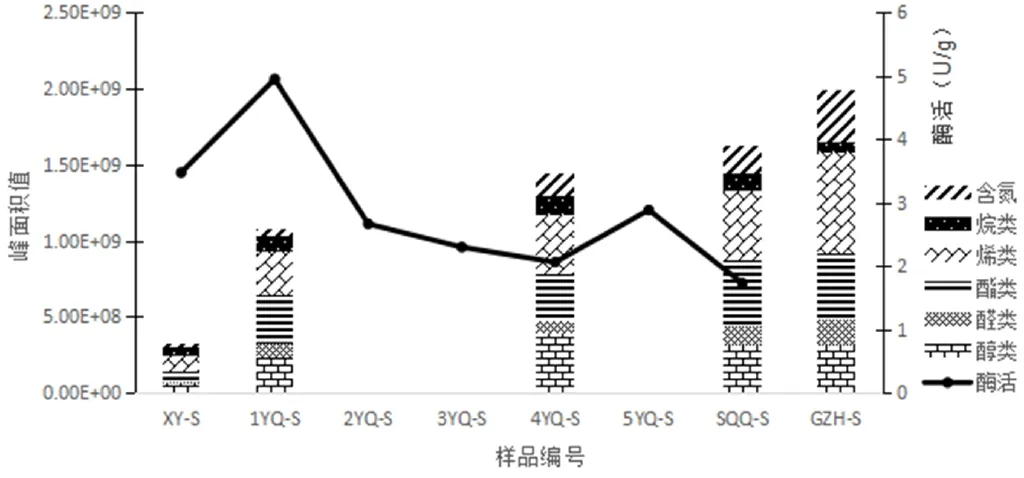
图2 水仙加工过程茁-葡萄糖苷酶活性、香气峰面积总值及香气组分变化规律注:图中2YQ-S、3YQ-S、5YQ-S未测定峰面积值
水仙各工序香气组分峰面积值的变化规律:醛类呈缓慢上升趋势,烯类和含氮化合物呈显著上升趋势,三类化合物峰面积值均在GZH-S达到最高值,分别为1.65E+08、6.59E+08、3.46E+08;酯类峰面积值在SQQ-S达到最高值4.40E+08后下降;醇类和烷类峰面积值变化平缓,在4YQ-S达到最高值(3.87E+08、1.26E+08)后下降。
2.1.2水仙加工过程茁-葡萄糖苷酶活性变化
水仙加工过程茁-葡萄糖苷酶活性总体呈“双峰”型变化趋势,如图2所示。在萎凋过后1YQ-S达到第一个峰值4.95U/g,摇青过程中持续下降,至4YQ-S达到2.06U/g后上升,5YQ-S达到第二个峰值2.88U/g,SQQ-S达到最低值1.73U/g。萎凋过程酶活性水平均值为4.21U/g,做青过程酶活性水平均值为2.37U/g。整个加工过程出现两个峰值,但总体酶活性呈现下降趋势。
2.1.3茁-葡萄糖苷酶活性与醇系香气相关性分析
茶叶加工过程中,鲜叶中存在大量的以单萜烯醇[11]、芳香族醇[12]、脂肪族醇[13]等为配基的糖苷类前体物质。其中参与香气形成的主要是茁-葡萄糖苷、茁-樱草糖苷和少许其他类型二糖苷[14-16]。这些物质经过糖苷酶作用,水解释放出挥发性配基,如芳樟醇、香叶醇、橙花叔醇等,这是茶叶花果香气的重要来源[17]。
晒青过程随着茁-葡萄糖苷酶活性的升高香气总量显著增加;1YQ-S 至SQQ-S整个做青过程香气总量伴随苷酶活性的下降和增加,呈现此消彼长的趋势。分析酶活性与香气峰面积值线性相关性发现两者呈高度指数负相关,其线性相关公式为Y= 2E+09e-0.1006x(R2=0.931)。

图3 水仙加工过程茁-葡萄糖苷酶活性与主要醇系香气成分峰面积值相关性分析
茁-葡萄糖苷酶活性与主要醇系香气成分峰面积值的关系如图3所示,在晒青阶段和做青1YQ-S至4YQ-S阶段,橙花叔醇、香叶醇、芳樟醇、雪松醇含量均与酶活性有相同的变化趋势,做青后期4YQ-S至SQQ-S阶段,伴随茁-葡萄糖苷酶活性下降,芳樟醇含量增加,而橙花叔醇、香叶醇、雪松醇均呈现上升趋势。
分析酶活性与醇系香气峰面积总值及主要醇系香气成分峰面积值相关性发现,1YQ-S至SQQ-S阶段酶活性分别与橙花叔醇、香叶醇峰面积值呈高度指数负相关,其线性相关公式分别为Y=1E+09e-1.1511x(R2= 0.973)、Y=6E+08e-1.1496x(R2=0.999),与芳樟醇、雪松醇峰面积值分别呈高度和较高指数正相关,公式为Y=1E+07e0.5494x(R2=0.946)、Y=1E+06e0.8851x(R2=0.872),而与醇系香气相关性较低。
2.2肉桂加工过程香气成分与茁-葡萄糖苷酶活性变化
2.2.1肉桂加工过程香气成分分析
肉桂加工过程香气总离子流图(以XY-R和GZH-R为例)如图4所示。

图4 肉桂在制品香气总离子流图

图5 肉桂加工过程茁-葡萄糖苷酶活性、香气峰面积总值及香气组分变化规律注:图中2YQ-R、3YQ-R、5YQ-S、7YQ-R未测定峰面积值
肉桂加工过程香气成分总量呈上升趋势,如图5所示。其中1YQ-R 至GZH-R的香气成分总峰面积值分别为XY-R的1.41、2.6、2.97、4.59、3.35倍,4YQ-R至SQQ-R的涨幅最为明显,SQQ-R的香气成分总峰面积达到最高值后下降。
肉桂各工序香气组分峰面积值的变化规律:醇类、酯类和烯类均为显著上升后下降趋势,醛类和含氮化合物则先缓慢上升后下降,五类化合物峰面积值均在SQQ-R达到最高值,分别为7.88E+08、5.51E+08、6.59E+08、2.10E+08、2.45E+08;烷类峰面积值变化平缓,4YQ-R达到最高值1.22E+08。
2.2.2肉桂加工过程茁-葡萄糖苷酶活性变化
肉桂加工过程茁-葡萄糖苷酶活性总体呈现波动下降趋势,如图5所示。在萎凋过后1YQ-R达到最高值5.25U/g,摇青过程中波动下降,至4YQ-R和7YQ-R达到做青过程两个峰值,分别是4.66U/g、3.93U/g,至SQQ-R达到最低值2.59U/g。萎凋过程酶活性水平均值为4.7U/g,做青过程酶活性水平均值为4.15U/g。整个加工过程出现三个峰值,总体酶活性呈现下降趋势。
2.2.3茁-葡萄糖苷酶活性与醇系香气相关性分析
晒青阶段香气总量随着酶活性的升高而增加;做青阶段1YQ-R至4YQ-R和6YQ-R至SQQ-R阶段香气总量随着酶活性的升高而下降,4YQ-R至6YQ-R阶段香气总量随着酶活性升高而递增。分析酶活性与香气峰面积值相关性发现两者呈较高指数负相关,其线性相关公式为Y=8E+09e-0.39783x(R2=0.897)。
茁-葡萄糖苷酶活性与主要醇系香气成分峰面积值的关系如图6所示,在晒青阶段,随着酶活性升高,橙花叔醇、香叶醇含量增加,而芳樟醇及其氧化物呈现下降趋势,做青前期1YQ-R至4YQ-R,随着酶活性下降,橙花叔醇、香叶醇、芳樟醇及其氧化物含量均呈现上升趋势,做青中期4YQ-R至6YQ-R,随着酶活性下降,橙花叔醇、香叶醇含量增加,而芳樟醇及其氧化物呈现下降趋势,做青后期6YQ-R至SQQ-R,橙花叔醇、香叶醇、芳樟醇及其氧化物含量均呈现显著上升趋势。

图6 肉桂加工过程茁-葡萄糖苷酶活性与主要醇系香气成分峰面积值相关性分析注:图中SQQ-R橙花叔醇含量为原始含量的1/5;SQQ-R香叶醇含量为原始含量的1/2。
分析酶活性与醇系香气及主要醇系香气成分峰面积值相关性发现,酶活性与橙花叔醇、香叶醇、芳樟醇氧化物I峰面积值均呈较高指数负相关,其线性相关公式分别为Y=1E+10e-0.8293x(R2=0.888)、Y=4E+09e-0.8753x(R2=0.869)、Y=2E+09e-1.0959x(R2=0.882),而与醇系香气及芳樟醇无明显相关性。
2.3水仙和肉桂加工过程茁-葡萄糖苷酶对香气形成的影响对比分析
2.3.1水仙和肉桂加工过程茁-葡萄糖苷酶活性变化对比分析
由图2和图5分析,在相同加工工艺下,水仙茁-葡萄糖苷酶活性变化范围为4.95U/g-1.73U/g,肉桂为5.25U/g-2.59U/g,是水仙的1.06-2.26倍。水仙和肉桂酶活性变化幅度不同,肉桂酶活性变化幅度为2.66U/g,水仙为3.22U/g,是肉桂的1.21倍。
水仙和肉桂酶活性变化趋势有一定的差异性,晒青阶段水仙和肉桂酶活性均呈显著上升趋势,水仙变化率大于肉桂;水仙做青阶段1YQ-S 至5YQ-S呈“凹”型变化趋势,肉桂做青阶段1YQ-R至4YQ-R与4YQR至7YQ-R呈“双凹”型变化趋势,变化率小于水仙,水仙和肉桂做青后期5YQ-S(7YQ-R)至SQQ-S酶活性均下降,肉桂变化率小于水仙,两者整个加工过程中酶活性均呈现下降趋势。
2.3.2水仙和肉桂加工过程香气变化对比分析
XY至SQQ水仙和肉桂香气总量均呈现上升趋势,水仙SQQ-S至GZH-S的香气总量依旧上升,而肉桂香气总量明显下降。5个阶段肉桂香气总量分别是水仙的1.59、0.67、1.12、1.51、0.9倍;且肉桂变化率小于水仙。
对比水仙和肉桂醇系香气的变化趋势发现,XY至4YQ-S(6YQ-R)阶段水仙和肉桂醇系香气均为上升趋势,4YQ-S(6YQ-R)至SQQ水仙醇系香气下降而肉桂含量增加,SQQ至GZH两者醇系香气含量均下降。5个阶段肉桂醇系香气含量分别是水仙的2.48、0.98、1.42、2.49、2.45倍;且肉桂变化率小于水仙。
2.3.3水仙和肉桂加工过程茁-葡萄糖苷酶对香气形成的影响对比分析
水仙茁-葡萄糖苷酶活性和香气总量均低于肉桂,说明酶活性对香气的形成总体呈正相关;对各阶段进行分析得出,水仙和肉桂茁-葡萄糖苷酶活性与香气总量总体呈现“正相关寅负相关”。
分析酶活性与醇系香气发现,水仙酶活性与醇系香气总体呈现“正相关寅负相关寅正相关”的关系,肉桂酶活性对醇系香气及香气总量的影响基本一致,在加工中各阶段的相关性也基本相同,均呈现“正相关寅负相关”的变化趋势。
3 讨论
3.1水仙和肉桂茁-葡萄糖苷酶活性变化差异
乌龙茶在做青过程中,摇青使叶细胞破损,液泡中糖苷类前体外溢和胞质中的茁-葡萄糖苷酶充分接触,加快键合态香气的释放速度[18]。而摊放过程,茶叶细胞内多种酶系代谢加强,进行充分水解作用,则酶活性增强,因做青工艺与程度差异,茶树叶片失水率不同,细胞破损率不同,导致酶活性差异。
本研究结果表明,加工过程中茁-葡萄糖苷酶活性总体水平表现为肉桂跃水仙,肉桂酶活性是水仙的1.06-2.26倍;两者变化趋势差异不明显,酶活性变化幅度表现为水仙跃肉桂,且水仙和肉桂酶活性总体均呈下降趋势。
张正竹等[19]研究表明加工初期,茶叶组织逐渐失水,细胞呼吸增强,茁-葡萄糖苷酶活性升高,刘立华等[20]认为揉捻使得茶叶细胞破碎,引起多酚类物质大量外溢,它与蛋白酶作用使之沉淀,从而降低茁-葡萄糖苷酶活性。因此本研究中水仙和肉桂酶活性均在1YQ达到最高值,而在做青结束时酶活性有所下降。
3.2水仙和肉桂茁-葡萄糖苷酶活性变化与香气形成的关系
本研究表明,水仙晒青阶段随着酶活性升高,香气总量、醇系香气及主要醇系香气成分含量均增加;做青前期香气总量、醇系香气含量增加而主要醇系香气成分含量减少,做青后期香气总量及橙花叔醇、香叶醇、雪松醇含量增加而醇系香气及芳樟醇含量减少。
肉桂晒青阶段随着酶活性升高,香气总量、醇系香气及橙花叔醇、香叶醇含量增加而芳樟醇及其氧化物含量减少;做青前期香气总量、醇系香气及主要醇系香气成分含量均增加,做青中期香气总量、醇系香气及橙花叔醇、香叶醇含量增加而芳樟醇及其氧化物含量减少,做青后期香气总量、醇系香气及主要醇系香气成分含量均增加。
由于晒青过程随着茶叶含水量逐渐降低,呼吸作用增强且酶活性明显增强,酶水解糖苷类香气前体释放香气,故晒青阶段水仙和肉桂的香气均增加,而做青阶段酶活性受水分、膜透性以及抗性机制的影响呈现动态变化,故加工过程中水仙和肉桂香气成分也波动变化。
酶活性与香气成指数负相关的主要原因是茁-葡萄糖苷酶活性的高低引起了香气含量变化,由于活性启动机制先于香气含量的增加,酶活性预先增强导致水解作用加剧,使香气含量增加;加工过程中香气形成为累积效果,而酶活性为实时动态变化,随着加工的延续,香气含量增加而细胞活力下降,酶活性降低符合生理变化过程。香气形成机理较为复杂,加工工艺繁复引起细胞内、外部环境不稳定,因此酶活性与香气变化相关性并不总是一致和统一。
参考文献
[1]姚月明,陈永霖.武夷肉桂名枞的生化特性[J].茶叶科学,1989,9 (2):151-154.
[2]宛晓春.茶叶生物化学[M].北京:中国农业出版社,2003.8:119-127.
[3]丁勇.茶叶中糖苷酶类的研究进展[J].中国茶叶加工,2001,(4):34-36.
[4] YANG ZY,BALDERMANN S,WATANABE N.
Recent studies of the volatile compounds in tea [J].Food Research International,2013,53(2):585-599.
[5]陈艺元.永福高山乌龙茶品质形成机理研究[D].福州:福建农林大学,硕士论文,2014.
[6]张娴静.不同工艺铁观音香气形成的生化及分子生物学机制研究[D].福州:福建农林大学,硕士论文,2012.
[7]张文锦,王峰,翁伯琦.武夷山市茶业现状与发展对策[J].福建农业学报,2012,27( 11):1264 -1268.
[8]修明.武夷岩茶品质特征及审评方法讨[J].中国茶叶加工.2004,(1):40.
[9]吴乔.乌龙茶做青对p-葡萄糖苷酶活性及酶学性质的影响[D].福州:福建农林大学,硕士论文,2013.
[10]刘琳燕,周子维,邓慧莉,等.不同年份白茶的香气成分[J].福建农林大学学报,2015,(1):28.
[11] KOBAYASHI A,KUBOTA K,JOKI Y,et al.(Z)-3-Hexenyl-茁-D-glucopyranoside in fresh tea leaves as a precursor of green odor [J].Bioscience,biotechnology,and biochemistry,1994,58(3):687-690.
[12] YANO M,JOKI Y,MUTOH H,et al.Benzyl glucoside from tea leaves [J].Agric.Biol.Chem.,1991,55(4):1205-1806.
[13] KOBAYASHI A,KUBOTA K,JOKI Y,et al.(Z)-3-Hexenyl-茁-D-glucopyranoside in fresh tea leaves as a precursor of green odor [J]. Bioscience,biotechnology,and biochemistry,1994,58(3):592-593.
[14] GUO W,SAKATA K,WATANABE N,et al.Geranyl-6-O-茁-dxylopyl-茁-d-glucopyranoside isolated as all aroma precursor from tea leaves for oolong tea[J].Phytochemistry,1993,33(6):1373-1375.
[15] GUO W,HOSOI R,SAKATA K,et al.(S)-linalyl,2-phenylethyl,and benzyl disaccharde glycosides isolated as aroma precursors from oolong tea leaves[J].Bioscience,biotechnology,and biochemistry,1994,58(8):l532.
[16] MOON JH,WATANABE N,IJIRNA Y,et al.Cis -and trans -lingalool 3,7-oxides and methyl salicylate glycosides and (Z)-3-hexenyl茁-D-glucepyraneside as aroma precursors from tea leaves for oolong tea[J].Bioscience,biotechnology,and biochemistry,1996,60(11):1815-1819.
[17] SAKATA K,GUO W,MOON J -H.Tea Chemistry Part II:With special reference to tea aroma precursors [M].New Delhi:Aravali Books Internationa,1999:692-704.
[18]骆耀平,董尚胜,童启庆,等.7个茶树品种新梢生育过程中葡萄糖苷酶活性变化[J].茶叶学,1997,17(增):104-107.
[19]张正竹,宛晓春,施兆鹏,等.鲜茶叶摊放过程中呼吸速率、葡萄糖苷酶活性、游离态香气和糖苷类香气前体含量的变化[J].植物生理学通讯,2003,39(2):134-136.
[20]刘莉华,宛晓春,文勇,等.祁门红茶加工过程中葡萄糖苷酶活性变化研究[J].安徽农业大学学报,2003,30(4):386-389.
通讯作者:孙云(1964-),女,教授,博士,博士生导师,研究方向:茶叶加工与生物技术,E-mail:sunyun1125@126.com
作者简介:邓慧莉(1991-),女,硕士研究生,研究方向:茶叶加工与加工工程。
基金项目:福建省现代农业(茶叶)产业技术体系项目(2060302闽财指[2014] 1032号)
