大口黑鲈GHRH基因启动子区域序列分析及其活性检测
2016-03-04马冬梅韩林强白俊杰
马冬梅,韩林强,白俊杰
(1.中国水产科学研究院珠江水产研究所,农业部热带亚热带鱼类选育与养殖重点开放实验室,广州 510380;2.淡水水产健康养殖湖北省协同创新中心,武汉 430070)
大口黑鲈GHRH基因启动子区域序列分析及其活性检测
马冬梅,韩林强,白俊杰
(1.中国水产科学研究院珠江水产研究所,农业部热带亚热带鱼类选育与养殖重点开放实验室,广州 510380;2.淡水水产健康养殖湖北省协同创新中心,武汉 430070)
生长激素释放激素(growth hormone releasing hormone,GHRH)是下丘脑弓状核合成和分泌的小分子多肽,其主要功能是调节垂体细胞合成和释放生长激素。为研究大口黑鲈(Micropterus salmoides)GHRH基因5’侧翼启动子区域的活性和该区域中潜在的转录因子对GHRH基因表达的调控作用,对该基因5’端启动子区域约1400 bp长度的片段进行序列分析,预测顺式作用元件,获得了Oct-1、SP1、NF-1、C/EBPalp和C/EBP等多个潜在的调控GHRH基因表达的调节因子结合位点序列。在包括外显子1和内含子1的GHRH基因5’侧翼区两侧加入两个限制性酶切位点Xho I和Bam H I,对其进行改造,并将该片段插入红色荧光蛋白报告基因载体pDsRed2-1,构建了重组表达质粒pGHRH1-RFP。同时,用不含有外显子1和内含子1的GHRH基因5’侧翼区构建重组表达质粒pGHRH-RFP。将质粒pGHRH1-RFP和pGHRH-RFP转染鲤(Cyprinus carpio)上皮细胞(epithelioma papillosum cyprinid,EPC)。经过48 h的培养,在pGHRH1-RFP转染的部分细胞中检测到红色荧光蛋白表达。又将pGHRH1-RFP或pGHRH-RFP质粒注射到斑马鱼(Danio rerio)一细胞或二细胞期的胚胎中,注射了pGHRH1-RFP的胚胎在受精后48 h约有22.5%能检测到有红色荧光蛋白表达,受精后72 h约有29%的仔鱼检测到红色荧光蛋白表达。实验结果表明,目前分离到的GHRH基因5’侧翼序列具有启动基因表达的活性,且该基因的内含子1和外显子1是启动子的活性所必需的。另外,pGHRH1-RFP质粒注射的斑马鱼胚胎只能在胚胎和仔鱼的脊椎和肌肉中检测到RFP的表达,而在脑中没有检测到表达。推测扩增到的大口黑鲈GHRH启动子序列1407 bp(-1043 bp~362 bp)只是起到了驱动RFP脊椎和骨骼肌表达的作用,而不包括驱动在脑组织中特异性表达的启动子,本研究为GHRH基因功能的深入分析奠定了基础。
生长激素释放激素;启动子活性;大口黑鲈
生长激素释放激素(growth hormone releasing hormone,GHRH)是一种小分子多肽,为PACAP/胰高血糖素(pituitary adenylate cyclase-activating polypeptide,PACAP/glucagon)超家族成员,主要由丘脑下部的弓状核合成和分泌[1-2],其主要的功能是通过与GHRH受体结合刺激垂体合成和分泌生长激素(growth hormone,GH)[3]。GHRH在胚胎期对调节生长激素细胞增殖分化、脑垂体形成等有重要作用[4];外源适度增加动物体内GHRH的含量能够加快动物的生长[5];人类GHRH受体基因的突变或缺失会导致侏儒症、巨人症等疾病[6],但人类GHRH基因本身的突变或缺失引起的疾病还未见报道[7]。GHRH的精确表达以及对GH时空表达的准确调节,对于动物器官的形成和发育来说是必不可少的,该基因启动子区域顺式调控元件的突变极可能会影响到启动子的活性,从而影响到基因的正确表达,因此分析GHRH基因启动子的序列和活性是十分有意义的一项工作,可为分析启动子序列突变对其活性的影响奠定基础。
在鱼类启动子的研究中,管家基因β-actin的启动子研究得最为深入[8-9],并在转基因鱼研究中得到了较为广泛应用[10-11]。而热激蛋白70(HSP 70)[12]、肌肉生长抑制素(myostatin)[13]、促甲状腺激素(TSHβ)[14]、胰岛素样生长因子-II(IGF-II)[15]等多种基因的启动子也得到了研究,为深入研究基因功能、基因表达调控和基因之间相互作用奠定了基础。目前,GHRH基因启动子在人(Homo sapiens)、小鼠(Musmusculus)和大鼠(Rattus norregicus)中都有研究[16-19],且Gsh-1基因对大鼠GHRH表达的调控功能得到了深入的分析[19]。但鱼类GHRH启动子研究较少,对大口黑鲈(Micropterus salmoides)GHRH基因的研究发现,其5’侧翼序列中存在一个66 bp的插入/缺失位点(c.-923_-858del),该插入/缺失位点为隐性致死位点[20]。
为进一步探讨大口黑鲈GHRH基因启动子区域突变对其功能的影响,本研究分析了大口黑鲈GHRH基因5’侧翼约1400 bp序列潜在的顺式作用调控元件,并构建了重组红色荧光蛋白质粒,在鲤(Cyprinus carpio)上皮细胞(EPC)和斑马鱼(Danio rerio)体内初步分析了该片段的启动子活性,以为深入分析GHRH基因功能奠定基础。
1 材料和方法
1.1 实验鱼和细胞系
用于实验的AB品系斑马鱼由本实验室繁育、饲养。选择6月龄斑马鱼成鱼雌鱼5 ind、雄鱼5 ind,平均体质量1.2 g,平均全长4.5 cm,雌雄分别饲养,繁殖斑马鱼时一雌一雄配对,收集受精卵用于显微注射。实验用鲤上皮细胞系(EPC)由中国水产科学研究院珠江水产研究所实验室保存。
1.2 启动子区域的转录元件分析
用Transcription Element Search System软件(http://www.cbil.upenn.edu/cgi-bin/tess/tess)预测大口黑鲈GHRH基因5’侧翼区[20]1451 bp片段上顺式作用元件,所有参数设置均使用默认值,核心序列矩阵相似度与序列矩阵相似度比值均大于0.8。
1.3 表达质粒的构建
根据大口黑鲈GHRH基因的5’侧翼序列[20]设计特异引物,转录启始位点定义为 +1,GHRH1-pF:5’-CCGCTCGAGCGGGCTGGTCTGTT AAATACAAGGT-3’和GHRH1-pR:5’-CGGGATC CAGCTAGTCGTGGAGAAGAATGGACAG-3’(-1043 bp~362 bp,扩增产物包括内含子1和外显子1);GHRH-pR:5’-CGGGATCCTTCACTCTCAT CTCTCATCCTC-3’(-1043 bp~50 bp,扩增产物不包括完整的内含子1和外显子1),在扩增引物两端引入XhoI和BamH I酶切位点,以大口黑鲈基因组为模板进行PCR扩增。扩增产物经限制性内切酶Xho I和Bam H I(Fermentas公司)酶切纯化后,插入红色荧光蛋白报告基因载体pDsRed2-1(Clontech公司),转化大肠杆菌E.coliDH5α,测序验证重组质粒。得到的重组质粒命名为pGHRH1-RFP和pGHRH-RFP。
1.4 细胞培养和转染
用质粒DNA提取试剂盒(北京天根公司)提取并纯化质粒pGHRH1-RFP和pGHRH-RFP,溶解于H2O中至终浓度为50μg·mL-1。将冻存的鲤上皮瘤细胞(epithelioma papulosum cyprini,EPC)复苏后,在CO2培养箱中用含有10%小牛血清(Gibco公司)的M199培养基37℃培养至单层,然后将EPC细胞接种到24孔板内,每孔约1 ×105cell细胞,37℃培养22~24 h后用于转染。当24孔板的细胞密度达到90%时进行转染,分别将800 ng的pGHRH1-RFP和pGHRH-RFP质粒与2μL脂质体2000转染试剂(Invitrogen公司)混合,加入Opti-MEM培养基至100μL用于每一孔的转染,同时以800 ng不含GHRH基因启动子的pDsRed2-1质粒作为对照。转染方法参照转染试剂说明书。培养48 h,在Nikon荧光显微镜下观察红色荧光蛋白在EPC细胞中的表达。
1.5 显微注射
将重组质粒注射到斑马鱼受精卵内瞬时表达进行启动子活性分析。在斑马鱼受精卵一细胞或二细胞时分别注射约2 nL pGHRH1-RFP或pGHRH-RFP质粒DNA溶液至细胞质,同时以注射空载体pDsRed2-1为对照。胚胎在曝气水中培养,用MS222麻醉后,在Zeiss荧光显微镜下观察红色荧光蛋白在斑马鱼体内的表达。
2 结果与分析
2.1 大口黑鲈GHRH基因5’侧翼区域转录因子作用位点预测
用Transcription Element Search System软件预测分析了GHRH基因从-1803 bp到362 bp(以转录起始位点记为+1)潜在启动子区域上的顺式作用元件,预测结果见图1。在GHRH基因5’侧翼及内含子1和外显子1区域内存在启动子转录元件TATA框和GATA框各1个,八聚体转录因子1(Oct-1)结合位点6个,核转录因子SP1结合位点4个,肝细胞核因子HNF-1结合位点2个,核转录因子NF-1结合位点4个,Homeobox结合位点3个,脂肪形成转录因子C/EBPalp结合位点13个,增强子结合蛋白C/EBP结合位点8个,垂体特异性转录因子Pit-1a和Pit-1结合位点3个,同源转录因子Gsh-1、上游激活因子USF、激活蛋白AP-1转录因子、生物钟基因REV-ErbA、环磷腺苷效应元件结合蛋白CREB、小眼畸形相关转录因子MITF、抑制糖皮质激素受体GR、原癌基因c-Jun、转录因子PU.1和转录因子Oct-2.1结合位点各1个。
2.2 pGHRH1-RFP质粒在真核细胞中的表达
利用脂质体2000转染试剂将重组质粒pGHRH1-RFP、pGHRH-RFP和pDsRed2-1(阴性对照)转染EPC细胞。转染24 h后,荧光显微镜下镜检观察,可以看到经pGHRH1-RFP转染的细胞孔中,部分细胞发出明显的红色荧光,而质粒pGHRH-RFP转染的细胞和阴性对照组没有表达红色荧光的细胞,结果见图2。
2.3 pGHRH1-RFP质粒在斑马鱼体内的表达
将pGHRH1-RFP质粒注射到斑马鱼的受精卵中,48 h后在荧光显微镜下观察,约200 cell胚胎中有45个表达红色荧光;72h斑马鱼胚胎出膜后在荧光显微镜下观察,56 ind活仔鱼中有16 ind表达红色荧光,约占存活个体的29%。红色荧光蛋白主要表达在斑马鱼的肌肉和脊椎骨中(图3),而脑中的表达不显明。用pGHRH-RFP和pDsRed2-1质粒注射的斑马鱼没有观察到有红色荧光蛋白的表达。
3 讨论
本研究经软件预测大口黑鲈GHRH基因5’侧翼约1400 bp区域序列中存在多个顺式调控元件,这些顺式调控元件中包括多个Oct-1、SP1、C/EBPalp和C/EBP结合位点,推测GHRH受到多种因子的精密调控,Oct-1、SP1、C/EBPalp和C/EBP可能在大口黑鲈GHRH基因的时空表达活性调节中起到重要作用。另外,顺式作用元件中还包括调节因子CREB与Gsh-1的结合位点各1个,小鼠的GHRH启动子的研究表明,CREB与Gsh-1的共表达对于GHRH启动子的活性具有重要的调节作用[19]。
完整的内含子1和外显子1序列是GHRH启动子活性所必需的,用不完整的内含子1和外显子1序列构建的重组质粒,经细胞转染和显微注射两种方法,都检测不到RFP的表达,说明内含子1和外显子1区域中含有GHRH启动子活性所必须的重要调节因子结合位点,具体的调节机制还有待进一步研究。人类的金属蛋白酶1基因组织抑制剂基因(TIMP-1)的内含子1和外显子1序列对于该基因在成纤维细胞中的转录活性也是必须的[21]。
利用红色荧光蛋白报告载体pDsRed2-1,以包括外显子1和内含子1在内的1407 bp的GHRH基因5’侧翼区序列为启动子序列,构建GHRH启动子红色荧光蛋白表达质粒,注射斑马鱼受精卵后,检测到红色荧光蛋白主要在其胚胎中部分脊椎骨和骨骼肌有表达,而没有检测到在脑组织中的明显表达。迄今为止,尚未见鱼类GHRH早期基因表达的研究报道,可见GHRH早期基因表达的复杂性,而本研究为鱼类GHRH早期基因表达的研究提供了一定的数据与参考。目前,有研究者用RT-PCT的方法,分别在斑点叉尾鮰(Ictalurus punctatus)的骨骼肌中检测到了GHRH-LP基因的表达[22]及在中国林蛙的脊椎中检测到GHRH-LP明显的表达[23]。而关于GHRH基因的进化研究认为,GHRH基因、GHRH-LP基因以及PHI-VIP基因是由一个共同的祖先基因进化而来的[24]。推测GHRH和GHRH-LP这一类同源基因在进化过程中仍保留了启动子功能区域的相似性,时空表达上也具有相似的特征。

图1 大口黑鲈GHRH基因5’侧翼片段转录因子结合位点分析Fig.1 Analysis of transcription factor binding sites in 5’flanking region of GHRH gene from Micropterus salmoides
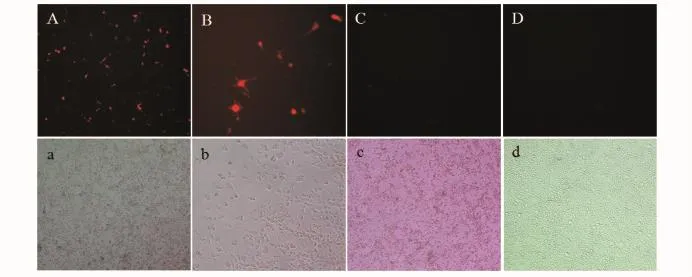
图2 pGHRH1-RFP质粒在EPC细胞中的表达(24 h)Fig.2 Expression of pGHRH-RFP in the EPC cells(24 h)

图3 pGHRH1-RFP质粒在斑马鱼胚胎和仔鱼中的表达Fig.3 Expression of pGHRH1-RFP in the embryos and larvae of zebrafish Danio rerio
对大口黑鲈GHRH基因mRNA的组织分布研究只在延脑和前脑组织检测到了GHRH的表达,而在肌肉中未见表达[25];对金鱼(Carassiusauratus)的研究中也只在大脑(brain)内检测到了GHRH基因的表达[24]。本研究在斑马鱼的脑中没有检测到有规律的明显的表达,推测本研究扩增到的大口黑鲈GHRH启动子序列1407 bp(-1043 bp~362 bp)只是起到了驱动RFP脊椎和骨骼肌表达的作用,而不包括驱动在脑组织中特异性表达的启动子。对大鼠GHRH启动子的研究表明,GHRH mRNA在精巢、胎盘和下丘脑中具有不同的剪切方式,精巢和下丘脑的转录起始位点相距约10.7 kb,说明该基因不同组织特异调控区域分布于GHRH基因5’侧翼序列很大范围内的不同位置[18]。
本研究没有采用双荧光素酶方法检测启动子活性的方法,而是采用了构建红色荧光蛋白报告重组质粒细胞转染和显微注射斑马鱼受精卵的方法,在转染EPC细胞体外直接观察的检测方法更为简便、直接,而显微注射斑马鱼胚胎的体内检测方法则可以为研究基因的表达和功能提供更为详尽的依据。
[1] NAM B H,MOON JY,KIM Y O,etal.Molecular and functional analyses of growth hormone-releasing hormone(GHRH)from olive flounder(Paralichthysolivaceus)[J].Comparative Biochemistry and Physiology Part B Biochemistry and Molecular Biology,2011,159(2):84-91.
[2] MAYO K E,MILLER T,DEALMEIDA V,et al.Regulation of the pituitary somatotroph cell by GHRH and its receptor[J].Recent Progress in Hormone Research,2000(55):237-266.
[3] MAYO K E,CERELLIG M,LEBO R V,et al.Gene encoding human growth hormone-releasing factor precursor:structure,sequence,and chromosomal assignment[J].Proceedings of the National Academy of Sciences USA,1985,82(1):63-67.
[4] BILLESTRUP N,SWANSON LW,VALEW.Growth hormone releasing factor stimulates proliferation of somatostrophsin vitro[J].Proceedings of the National Academy of Sciences USA,1986,83(18):6854-6857.
[5] BAUMANN G.Mutations in the growth hormone releasing hormone receptor:a new form of dwarfism in humans[J].Growth Hormone&IGF Research,1999,9(SUPPL.B):24-30.
[6] DESAIM P,UPADHYE P S,KAMIJO T,et al.Growth hormone releasing hormone receptor(GHRH-r)gene mutation in Indian children with familial isolated growth hormone deficiency:a study from western India[J].Journal of Pediatric Endocrinology&Metabolism,2005,18(10):955-973.
[7] ZHANG Y,ZHU Y J,LIZ,etal.Injection of porcine growth hormone releasing hormone gene plasmid in skeletalmuscle increase piglets’growth and whole body protein turnover[J].Livestock Science,2008,115(2-3):279-286.
[8] FREDERICKSON R M,MICHEAU M R,IWAMOTO A,etal.5'flanking and first intron sequences of the human beta-actin gene required for efficient promoter activity[J].Nucleic Acids Research,1989,17(1):253-270.
[9] LIU Z J,MOAV B,FARASA J,et al.Functional analysis of elements affecting expression of the betaactin gene of carp[J].Molecular and Cellular Biology,1990,10(7):3432-3440.
[10] FENG J,LIG,LIU X,et al.Functional analysis of the promoter region of amphioxusβ-actin gene:a useful tool for driving gene expressionin vivo[J].Molecular Biology Reports,2014,41(10):6817-6826.
[11] JIAN Q,CHEN M,BAI J J,et al.Generation and characterization of a stable red fluorescent transgenic Tanichthys albonubes line[J].African Journal of Biotechnology,2012,11(30):7756-7765.
[12] QI J,LIU X D,LIU J X,et al.Molecular characterization of heat shock protein 70(HSP 70)promoter in Japanese flounder(Paralichthys olivaceus),and the association of Pohsp70 SNPs with heat-resistant trait[J].Fish&Shellfish Immunology,2014,39(2):503-511.
[13] 孙科军,刘希良,王开卓,等.斑鳜myostatin基因及其启动子的克隆与序列[J].基因组学与应用生物学,2012,31(2):133-140.
SUN K J,LIU X L,WANG K Z,etal.Cloning and sequence analysis of the myostatin gene and its promoter in golden mandarin fish(Siniperca scherzeri)[J].Genomics and Applied Biology,2012,31(2):133-140.
[14] WANG Y,SUN ZH,ZHOU L,et al.Grouper tshβ promoter-driven transgenic zebrafish marks proximal kidney tubule development[J].Plos One,2014,9(6):1-10.
[15] TSE M C,CHAN K M,CHENG C H.Cloning,characterization and promoter analysis of the common carpIGF-IIgene[J].Gene,2008,412(1-2):26-38.
[16] SOLLOSO A,BARREIRO L,SEOANE R,et al.GHRH proliferative action on somatotrophs is celltype specific and dependent on Pit-1/GHF-1 expression[J].Journal Cell Physiol,2008,215(1):140-50.
[17] NOGUÉSN,DELRÍO JA,PÉREZ-RIBA M,etal.Placenta-specific expression of the rat growth hormone-releasing hormone gene promoter in transgenic mice[J].Endocrinology,1997,138(8):3222-3227.
[18] SRIVASTAVA CH,MONTSB S,ROTHROCK JK,et al.Presence of a spermatogenic-specific promoter in the rat growth hormone-releasing hormone gene[J].Endocrinology,1995,136(4):1502-1508.
[19] MUTSUGA N,IWASAKIY,MORISHITA M,etal.Homeobox protein Gsh-1-dependent regulation of the rat GHRH gene promoter[J].Molecular Endocrinology,2001(12):2149-2156.
[20] MA D M,HAN L Q,BAI J J,et al.A 66-bp deletion in growth hormone releasing hormone gene 5’-flanking region with largemouth bass recessive embryonic lethal[J].Animal Genetics,2014,45(3):421-426.
[21] CLARK IM,ROWAN A D,EDWARDSDR,etal.Transcriptional activity of the human tissue inhibitor ofmetalloproteinases1(TIMP-1)gene in fibroblasts involves elements in the promoter,exon 1 and intron 1[J].Biochemical Journal,1997,324(Pt2):611-617.
[22] SMALL B C,NONNEMAN D.Sequence and expression of a cDNA encoding both pituitary adenylate cyclase activating polypeptide and growth hormone-releasing hormone-like peptide in channel catfish(Ictalurus punctatus)[J].General and Comparative Endocrinology,2001,122(3):354-363.
[23] ALEXANDRE D,VAUDRY H,JÉGOU S,et al.Structure and distribution of the mRNAs encoding pituitary adenylate cyclase-activating polypeptide and growth hormone-releasing hormone-like peptide in the frog,Rana ridibunda[J].Journal of Comparative Neurology,2000,421(2):234-246.
[24] LEE L T O,SIU F K Y,TAM J K V,et al.Discovery of growth hormone releasing hormones and receptors in nonmammalian vertebrates[J].Proceedings of the National Academy of Sciences USA,2007,104(7):2133-2138.
[25] 韩林强,白俊杰,李胜杰.大口黑鲈GHRH-LP和GHRH基因序列同源性、基因结构和时序表达研究[J].水生生物学报,2011,35(3):473-481.
HAN L Q,BAI J J,LI S J,Comparison of gene structure,sequence homology and expression pattern of largemouth bass GHRH-LP and GHRH[J].Acta Hydrobiologica Sinica,2011,35(3):473-481.
Sequence and activity analysis of GHRH promoter region from M icropterus salmoides
MA Dong-mei,HAN Lin-qian,BAIJun-jie
(1.Key Laboratory of Tropical&Subtropical Fishery Resource Application&Cultivation,Ministry of Agriculture,Pearl River Fisheries Research Institute,Chinese Academy of Fishery Sciences,Guangzhou510380,China;2.Freshwater Aquaculture Collaborative Innovation Center of Hubei Province,Wuhan430070,China)
Growth hormone releasing hormone(GHRH)is a smallmolecular weight peptide released from the arcuate nucleus of hypothalamus.It mainly plays the role to regulate the synthesis and release of growth hormone(GH)from the anterior pituitary somatotrophs.Non-pituitary GHRH has awide spectrum of activity,includingmodulating cell proliferation,especially in malignant tissues,regulating differentiation of some cell types,and promoting healing of skin wounds.In order to analyze the activity of GHRH 5’flanking region and themechanism of the potential transcription factors regulating GHRH gene expression in largemouth bass(Micropterus salmoides),the cis-acting elementswere predicted by the transcription element search system for the 1400 bp-length fragment in 5’flanking region.Many sites related to regulating the GHRH expression were identified,includingmultiple of Oct-1,SP1,NF-1,C/EBPalp and C/EBP binding sites,which indicated that these sites probably played the key roles to regulate the spatiotemporal expression of GHRH.The GHRH 5’flanking region fragmentwith integrate exon 1 and intron 1 wasmodified by adding to two restrict enzyme sites,XhoI andBamH I,and was inserted into the red fluorescent protein(RFP)reporter gene vector pDsRed2-1.Then the recombined plasmid pGHRH1-RFP was constructed.Meanwhile,the recombinant plasmid pGHRH-RFP was constructed using the fragment without exon 1 and intron 1.The plasmids pGHRH1-RFP and pGHRH-RFPwere transfected epithelioma papillosum cyprini(EPC)cells.After 48 h of culture,the expression of the RFP was detected in EPC cells with pGHRH1-RFP,but not in the cells as negative controls or the cells transfected by pGHRH-RFP.At the same time,the plasmid pGHRH1-RFP or pGHRH-RFPwas alsomicroinjected in zebrafish embryos at the one cell or two cell stages.Embryos without microinjection served as negative controls.A total of 22.5%(n=45/200)embryos injected with pGHRH1-RFP could be detected RFP 48hpf(hours post fertilization)and 29%(n=16/56)fries could be detected RFP 72hpf.And the RFP was not detected in the zebrafish embryos and larvae microinjected with plasmid pGHRH-RFP and negative controls.The results showed that the isolated GHRH 5’flanking region had the activity of starting GHRH gene expression.And there are indispensable cis-elements in exon 1 and intron 1 of GHRH gene for promoter activity.Additionally,it is interesting that the RFP is found in spine and muscle,not in brain.In rat,GHRH gene is spliced into alternative upstream promoters in brain,gonads and placenta.It suggests that the cis-acting elements responsible for GHRH spatially specific expression in brain of largemouth bass have not been discovered yet.The current study can lay foundations for analysis of the functions of GHRH gene.
growth hormone releasing hormone(GHRH);promoter activity;largemouth bass(Micropterus salmoides)
Q 954
A
1004-2490(2016)04-0383-08
2015-08-27
国家自然科学基金项目(31001107);948计划重点项目(2011-G12);广东省省级科技计划项目(2015A020209035);国家科技支撑计划(2012BAD26B03)
马冬梅(1978-),女,辽宁抚顺人,博士,主要研究方向为鱼类遗传育种。Tel:020-81616127,E-mail:madongmei2003@163.com
白俊杰,研究员。Tel:020-81616129,E-mail:jjbai@163.net
