岛礁水域海藻场食物网基准生物的选择
2016-03-04周曦杰陈亮然章守宇汪振华
陈 玲,王 凯,周曦杰,赵 旭,陈亮然,章守宇,汪振华
(上海海洋大学海洋科学学院,上海 201306)
岛礁水域海藻场食物网基准生物的选择
陈 玲,王 凯,周曦杰,赵 旭,陈亮然,章守宇,汪振华
(上海海洋大学海洋科学学院,上海 201306)
岛礁海域海藻场是近岸浅海重要的湿地生境之一,其所在水域的食物网往往较开阔水域复杂,而利用稳定同位素方法来揭示其独特的物质传递和能量流动规律是当前国内外研究的趋势。在利用该技术之前,往往需选择恰当的基准生物以更准确地阐释海藻场食物网各阶层的营养关系。为此,于2014年春季(5月)和秋季(10月)在浙江省枸杞岛海藻场选取4种大型底栖动物优势种[蝾螺(Turbo petholatus)、角蝾螺(Turbocornutus)、条纹隔贻贝(Septifer virgatus)和带偏顶蛤(Modiolus comptus)]为实验对象,利用多元方差分析检验其δ13C和δ15N值的种内(间)时空差异,并分析造成该差异的原因,评估其作为基准生物的可行性。结果显示,4种底栖动物的δ13C和δ15N值受其食性和栖息地环境的影响,呈现不同程度的时空差异,发现带偏顶蛤和角蝾螺的δ13C和δ15N值相对更为稳定,因此可分别作为浮游和底层营养关系的基准生物。本文结果可为我国在东海岛礁水域开展相关研究提供基础数据和重要参考。
稳定同位素;海藻场;基准生物;大型底栖动物
岛礁水域海藻场生态系统食物网的物质和能量流动是特殊生境研究的一个关键领域,对于解释其结构和功能具有重要意义[1]。稳定同位素技术是研究水生生态系统食物网的有力手段[1-3],碳、氮稳定同位素比值(δ13C、δ15N)可以提供食物网中长时间尺度的摄食和能量流动信息[4]。基准生物可反映食物网生物最初物质来源的稳定同位素特征[5],因此在构建食物网前,需选择合适的基准生物,以准确分析其它生物的营养层次(trophic level)[5-6]和所处的食物链,从而确定生物间的营养关系[6]。王海霞等[7]在研究大连黑石礁海域时,选择初级生产者中的孔石莼(Uiva pertusa)作为基准生物估算其它海洋生物的营养层次。MATTHEWS等[8]分析发现浮游动物Daphnia食性单一,且营养层次较低(在浮游食物链中营养层次为2),因此以其作为基准生物分析了高营养层次生物间的营养关系。但近年来大量学者认为浮游动、植物或水生植物的δ13C和δ15N值受其生长环境理化因素的影响较大,已不适合作为基准生物[6,9]。双壳类、螺类等大型底栖动物生活史周期长、活动范围小且代谢率低,与其它活动能力强的消费者相比,能更好地反映栖息地的稳定同位素特征[1,6,10]。近年来双壳类已开始作为基准生物用于海洋生态系统食物网的构建[11-12],但不同海域的底栖动物种类及其δ13C和δ15N值存在差异[13-14],且在海藻场、红树林、海草场等特殊海洋生态系统中往往同时存在浮游植物链和底栖食物链,食物网结构复杂[3,6,],因此需根据不同海域实际情况进行选择。
枸杞岛位于浙江省舟山群岛东北部,其岛礁水域海藻场是典型的近岸海域子生态系统之一,也是鱼类、节肢动物、软体动物、棘皮动物等生物资源的重要栖息场所[15]。准确地选择基准生物对利用稳定同位素技术构建海藻场食物网,进而分析海藻场水生生物资源的养护机制,充实海藻场生态系统的理论有重要意义。本文选取了枸杞岛海藻场4种大型底栖动物优势种作为研究对象,通过测定其δ13C和δ15N值,分析了种内(间)时空差异和造成该差异的原因,并在此基础上选出最适基准生物,以深入研究枸杞岛海藻场食物网结构。
1 材料与方法
1.1 样品采集和处理
本研究样品采集于2014年春季(5月)和秋季(10月),基于前期对枸杞岛海藻场底栖动物群落研究结果,选择了4种大型底栖动物优势种[16-17],包 括 2种 牧 食 性 螺 类 [蝾 螺(Turbo petholatus)、角蝾螺(Turbo cornutus)]和2种滤食性双壳类[条纹隔贻贝(Septifer virgatus)、带偏顶蛤(Modiolus comptus)]。共设4个具有代表性的采样站点:北部的后头湾(St1)、西北部的黄石洞(St2)、西南部的龙泉(St3)和东南部的石浦(St4)(图1,表1)。如今自然水域或多或少都暴露在人类活动下,本文选取站点相对处于较低的干扰强度。St1海藻茂密,种类繁多,但离贻贝养殖区较近;St2靠近长江径流入海口,秋冬季受海流影响有少量陆源营养物质输入;St3春季潮流湍急且风浪较大,海水透明度低,因此春季未在St3采样;St4外侧有小范围贻贝养殖区。通过SCUBA水肺潜水采集生物样本,冷冻保存运回实验室。为减少不同生活史阶段个体稳定同位素比值差异对本研究结果的干扰,样品解冻后,根据个体大小将同种底栖动物分组,以各组测定结果的平均值进行分析。螺类取腹足肌肉[18],双壳类取闭壳肌肉[19],去离子水清洗后将同一组样品置于-55℃真空条件下冷冻干燥24 h,使用混合型球磨仪(Retsch MM440)粉碎样品。
1.2 稳定同位素测试
各组样品分别包被后送入元素分析仪-稳定同位素质谱仪联机(Flash EA 1112 HT-Delta V Advantages,Thermo公司)测定。样品在元素分析仪中高温燃烧后生成CO2和N2,质谱仪通过检测CO2的13C/12C以及14N的15N/14N,并与国际标准物(PDB)对比后计算出样品的δ13C值(δ13C的分析精度<±0.2‰),与国际标准物(纯净空气中的N2)对比后计算出样品的δ15N值(δ15N的分析精度<±0.3‰)。所有样品在国家海洋局第三海洋研究所同位素实验室测定。
δ13C和δ15N值以下面的公式进行计算:

式中:X指13C或15N,R表示13C/12C或15N/14N的值[6]。

图1 研究海域地理位置和采样站点Fig.1 Location of Gouqi Island and sampling stations
1.3 数据分析
运用多元方差分析方法检验大型底栖动物δ13C和δ15N值的时空差异性(α=0.05)。基于数据分析结果,选择同位素比值时空变化小的大型底栖动物作为基准生物。
2 结果与分析
2.1 稳定同位素测定值
春季海藻场大型底栖动物δ13C和δ15N值测定结果见表2所示,对比发现相同站点的螺类δ13C和δ15N值无种间差异(P>0.05),但条纹隔贻贝的 δ13C和 δ15N值显著高于带偏顶蛤(P<0.05),且相同站点的双壳类δ15N值与螺类无差异(P>0.05,图2-b),而δ13C值显著高于该站点的螺类(P<0.05)(图2-a)。
对比秋季相同站点的螺、双壳类δ13C和δ15N值后发现其种间无差异(P>0.1,表3),δ13C值显著高于该站点的螺类(图2-A),但双壳类δ15N值与螺类也无差异(图2-B)。
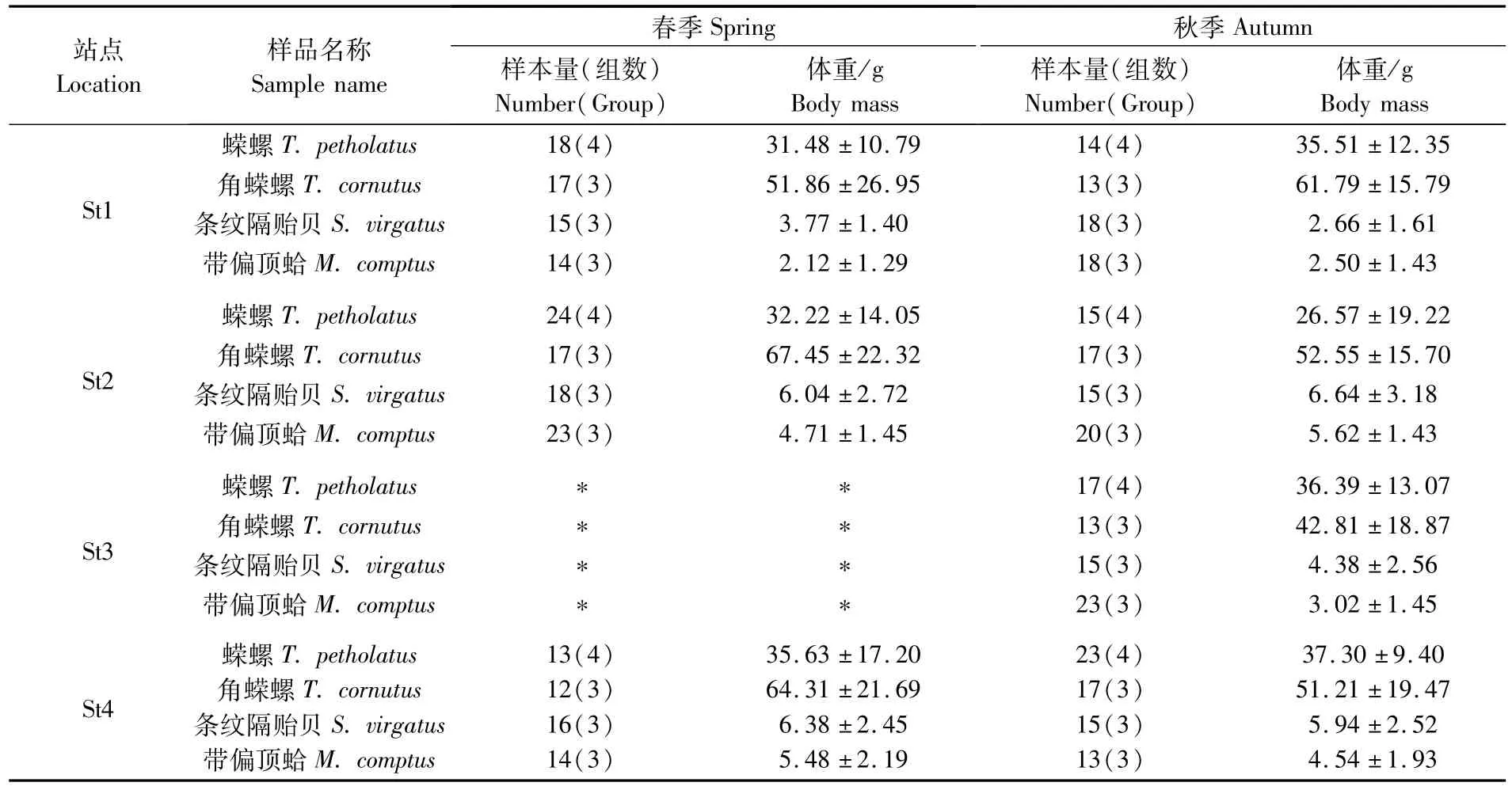
表1 海藻场大型底栖动物生物学参数Tab.1 Biological parameters of themacrobenthos collected from seaweed bed
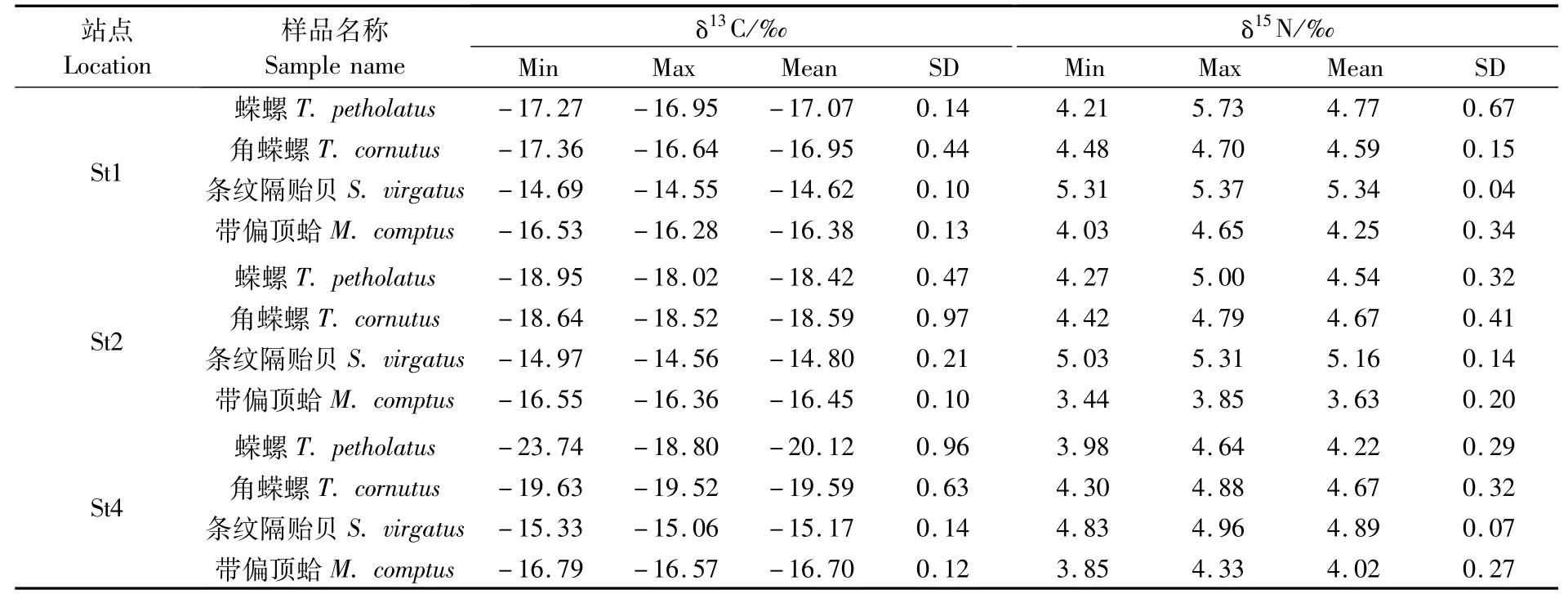
表2 春季海藻场大型底栖动物碳、氮稳定同位素的值Tab.2 Stable carbon and nitrogen isotopic ratios of themacrobenthos collected from seaweed bed in the spring

表3 秋季海藻场大型底栖动物碳、氮稳定同位素的值Tab.3 Stable carbon and nitrogen isotopic ratios of themacrobenthos collected from seaweed bed in the Autumn

图2 大型底栖动物在不同采样点的碳、氮稳定同位素值的方差分析(误差线表示标准偏差;春季:a和b;秋季:A和B)Fig.2 Stable carbon(above)and nitrogen(below)isotope signatures of themacrobenthos species between sam pling sites(Error bars represent the standard deviations.Spring:a and b;Autumn:A and B)

表4 春季海藻场大型底栖动物δ13C和δ15N的Turkey post-hoc分析结果Tab.4 Tukey post-hoc com parisons among sites forδ13C andδ15N of all species from seaweed field in the sp ring
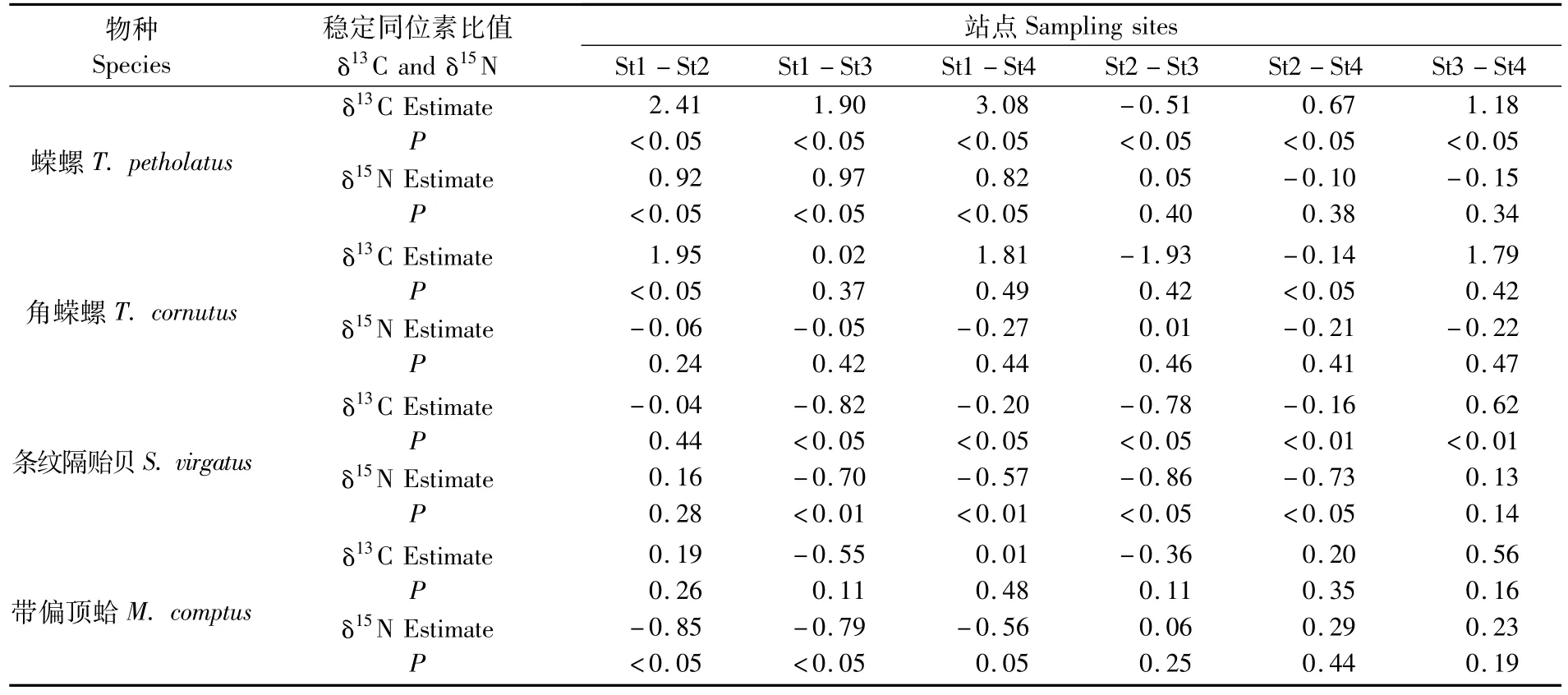
表5 秋季海藻场大型底栖动物δ13C和δ15N的Turkey post-hoc分析结果Tab.5 Tukey post-hoc com parisons among sites forδ13C andδ15N of all species from seaweed field in the Autumn
2.2 稳定同位素比值空间差异
从表4、5可以看出春、秋两季各站点间的δ13C值均存在显著差异(P<0.05);春季各站点间δ15N值无显著差异(表3),而秋季St1的δ15N值显著高于其它站点(P<0.05,表5)。春、秋两季各站点间角蝾螺的δ13C值存在一定差异,但δ15N值在各站点间均无显著差异(P>0.05)。
春季St4的条纹隔贻贝δ13C和δ15N值显著低于St1和St2(P<0.05,表4),带偏顶蛤的δ13C值在各站点间无差异,St2的δ15N值显著低于St1和St4(P<0.05)。从表5可以看出秋季St3条纹隔贻贝的δ13C值显著高于其它站点(P<0.05),且St3与St4的δ15N值较高(P<0.05);秋季带偏顶蛤的δ13C值在各站点间无差异(P>0.05),除St1的δ15N值较低外,其它站点间无显著差异(P>0.05)。
2.3 δ13C和δ15N值的季节差异
不同季节螺、双壳类季节差异如表6所示。
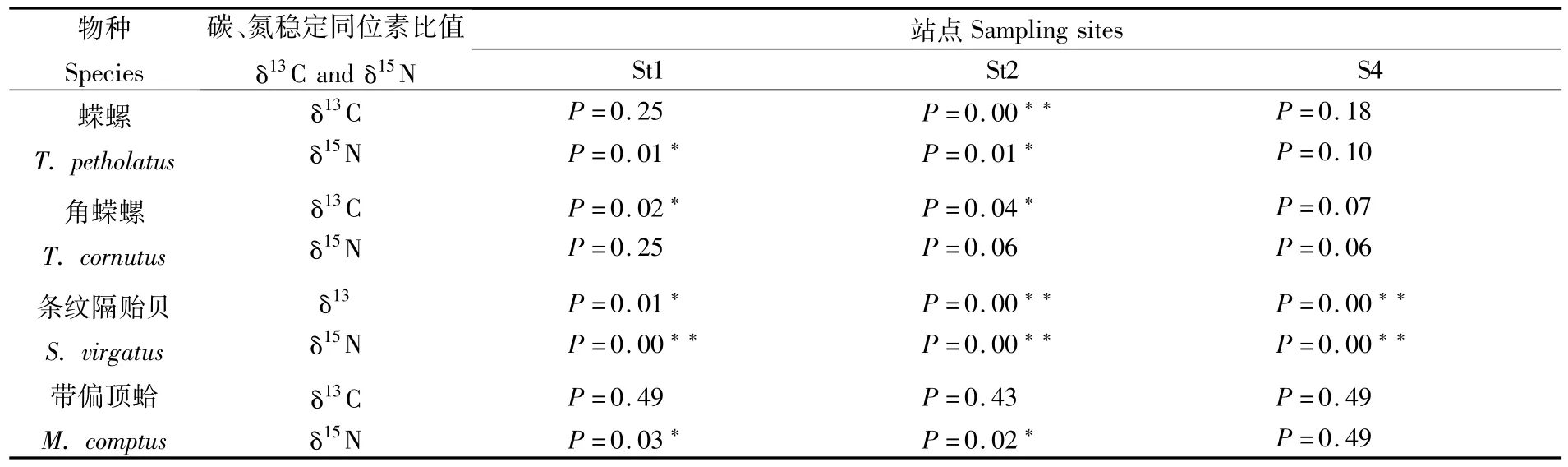
表6 海藻场大型底栖动物δ13C和δ15N值的季节差异Tab.6 Tukey post-hoc com parisons between seasons forδ13C andδ15N of all species from seaweed field
3 讨论
大型底栖动物生命周期较长,其组织周转时间长于浮游动、植物,且具有常年存在、食性简单的特点,与浮游生物相比,其稳定同位素比值受季节影响较小,因此其更适合作为基准生物[20]。FUKUMORI等[13]研究发现马氏珠母贝(Pinctadafucata martensii)肌肉组织周转时间约为120~180 d,认为可将其作为近海海洋生态系统食物网的基准生物。生物的δ13C值在各营养层次间变化较小,可被用于追溯C源,区分生物所处的食物链,而δ15N可分析生物的营养层次。岛礁水域海藻场同时存在浮游食物链与底栖食物链,单一的基准生物无法解释不同食物链生物间的营养关系,针对这一情况,已有学者尝试选择多个基准生物来分析食物网结构。李云凯等[10]以铜锈环棱螺(Bellamya aeruginosa)和河蚬(Corbicula fluminea)分别作为东太湖水生食物网底层和浮游能量传递途径的基准生物,计算了各鱼、虾类的营养层次。
本研究发现春、秋两季每个站点双壳类的δ13C值都显著高于该站点的螺类。生物机体的δ13C值会受其食性的影响[6],研究发现双壳类和螺类的食性不同[16-17,21],双壳类主要被动滤食水中的浮游性藻类、碎屑和浮游动物等,而螺类以啃食海藻为主,因此δ13C值的差异可能是由双壳类和螺类的食性不同造成的。DOI等[22]对日本北上市河口生态系统的大型底栖动物碳、氮稳定同位素研究发现双壳类δ13C值低于腹足类δ13C值,这与本研究不符,推测这可能是因为淡水中的螺类主要以沉积物有机质(SOM)为食[10-22],而海藻场中的角蝾螺和蝾螺以大型海藻为食[16]。δ15N值可用于确定生物的营养层次[6],春秋两季,相同站点螺类和双壳类的δ15N值均无差异,表明本研究选取的螺类与双壳类的营养层次较为接近,且作为营养层次较低的初级消费者均具有作为基准生物的可能性。
3.1 时空差异
双壳类的食物种类包括浮游植物、微型底栖生物、碎屑、沉积有机质等[13],作为滤食性的底栖动物,相同站点的双壳类其食物来源是类似的,但研究发现条纹隔贻贝和带偏顶蛤的空间差异程度不同,推测这与两种双壳类对食物的利用方式差异有关。尽管未对本研究中的双壳类进行喂养实验,但已有研究发现在相同摄食条件下,不同双壳类在摄入POM时存在选择性[23],且双壳类的稳定同位素比值与其机体周转速率也有一定关系[24]。海藻种类及其生物量发生季节性变化会受到海水温度的影响[25],秋季水温降低,暖水性海藻凋谢死亡后形成碎屑从而影响水体中POM的组成,这可能是造成条纹隔贻贝春、秋两季δ13C和δ15N值差异的原因。而带偏顶蛤未表现出季节性差异,推测这与其个体大小有关。与条纹隔贻贝相比,本研究中的带偏顶蛤个体偏小,其所能滤食的POM粒径也较小,而大型海藻死亡后产生的碎屑粒径较大,因此带偏顶蛤未受到其影响。对比各站点后发现角蝾螺的空间差异程度较低。研究发现枸杞岛海藻场角蝾螺能够摄食海带、铜藻、粗枝软骨藻等11种大型藻类[26],蝾螺所摄食的大型海藻种类虽未见报道,但已有研究证明螺类啃食海藻的过程存在摄食选择性[26],因此可能是角蝾螺和蝾螺对食物的选择性差异造成了其稳定同位素比值的空间差异。枸杞岛海藻场初级生产力复杂,春、秋两季大型海藻生物量和群落结构存在差异[25-27],且St2靠近长江径流入海口,秋季受海流影响有大量陆源营养物质输入,食物来源的季节变化和陆源营养物质的影响,可能引起了螺类δ13C和δ15N值的季节性变化。
3.2 基准生物
准确的基准生物对确定海洋生物营养层次十分关键[5-6]。早期学者研究近海海洋生态系统时仅选择一种双壳类作为基准生物,未考虑其是否适用于不同食物链,而研究表明不同食物链的基准生物的δ13C值存在差异[6,10,28]。滤食性的双壳类属于浮游食物链,而螺类属于底栖食物链[6,10],这与本研究中双壳类和螺类的δ13C值存在显著差异的结果相一致。为如实反映岛礁海域海藻场食物网结构,本研究建议分别选择一种双壳类和螺类作为两条食物链的基准生物。分析发现,不同双壳类的δ13C和δ15N值的空间差异程度不同。带偏顶蛤在枸杞岛海藻场各站点均分布广泛,且在春、秋两季其δ13C和δ15N值相对稳定。所以选择带偏顶蛤作为浮游能量传递途径的基准生物。
同一站点蝾螺和角蝾螺的δ13C和δ15N值无显著差异,与蝾螺相比,春、秋两季角蝾螺的δ13C和δ15N值在各站点间的差异均较小,且在春、秋两季间其δ13C和δ15N值相对稳定,表明角蝾螺更适合作为底层能量传递的基准生物。
4 小结
本文通过测定螺类、双壳类的δ13C和δ15N值,对比了其种内(间)时空差异,并分析了造成该差异的原因。结果表明,不同螺类、双壳类δ13C、δ15N值的时空差异程度不同,其δ13C和δ15N值与自身摄食选择性和栖息地环境有关,并建议选择带偏顶蛤和角蝾螺分别作为枸杞岛海藻场浮游和底栖食物链的基准生物。
本研究从时间和空间角度综合选择了δ13C和δ15N值相对稳定的底栖动物作为基准生物,为进一步利用稳定同位素技术构建岛礁水域海藻场食物网提供了基础数据。今后将分析本研究结果在其它岛礁海域海藻场的适用性,并采集基准生物的主要食物(大型海藻、浮游植物等),通过受控实验研究,深入分析不同初级生产者对基准生物δ13C、δ15N值的影响。
[1] MONCREIFFCA,SULLIVANM J.Trophic importance of epiphytic algae in subtropical seagrass beds:Evidence from multiple stable isotope analyses[J].Marine Ecology Progress Series,2001,215(1):93-106.
[2] 纪炜炜,李圣法,陈雪忠,等.基于稳定同位素方法的东海北部及其邻近水域主要游泳动物营养结构变化[J].海洋渔业,2015,37(6):494-500.
JIW W,LI S F,CHEN X Z,et al.Variation in trophic structure of nekton organisms from the northern East China Sea and adjacent waters based on stable isotope values[J].Marine Fisheries,2015,37(6):494-500.
[3] FREDRIKSEN S.Food web studies in a Norwegian kelp forest based on stable isotope(δ13C andδ15N)analysis[J].Marine Ecology Progress Series,2003,260(1):71-81.
[4] CABANA G,RASMUSSEN JB.Modelling food chain structure and contaminant bioaccumulation using stable nitrogen isotopes[J].Nature(London),1994,372(6503):255-257.
[5] 颜云榕,张武科,卢伙胜,等.应用碳、氮稳定同位素研究北部湾带鱼(Trichiurus lepturus)食性及营养级[J].海洋与湖沼,2012,43(1):192-200.
YAN Y R,ZHANG W K,LU H S,et al.Using stable isotopes to analyze feeding habits and trophic position of hairtail(Trichiurus lepturus)from the Bei Bu Gulf,South China Sea[J].Oceanologia Et Limnologia Sinica,2012,43(1):192-200.
[6] POST D M.Using stable isotopes to estimate trophic position:Models,methods,and assumptions[J].Ecology,2002,83(3):703-718.
[7] 王海霞,刘 瑀,鲍惠铭,等.黑石礁海域生物碳、氮稳定同位素组成的研究[J].海洋湖沼通报,2012(4):113-120.
WANG H X,LIU Y,BAO H M,et al.Study on marine biota using stable carbon and nitrogen isotopes in Hei Shi Jiao,Dalian[J].Transactions of Oceanology and Limnology,2012(4):113-120.
[8] MATTEWS B,MAZUMDER A.Compositional andinterlake variability of zooplankton affect baseline stable isotope signatures[J].Limnology and Oceanography,2003,48(5):1977-1987.
[9] 徐 军.氮稳定同位素基准的可变性及对营养级评价的影响[J].湖泊科学,2010,22(1):8-20.
XU J.Variability of stable nitrogen isotopic baselines and its consequence for trophic modeling[J].Journal of Lake Sciences,2010,22(1):8-20.
[10] 李云凯,贡 艺.基于碳、氮稳定同位素技术的东太湖水生食物网结构[J].生态学杂志,2014,33(6):1534-1538.
LIY K,GONG Y.Food web structure of the East Taihu by analysis of stable carbon and nitrogen isotopes[J].Chinese Journal of Ecology,2014,33(6):1534-1538.
[11] 杨国欢,侯秀琼,孙省利,等.流沙湾食物网结构的初探—基于稳定同位素方法的分析结果[J].水生生物学报,2013,37(1):150-156.
YANG G H,HOU X Q,SUN S L,et al.Construction food webmodel of Liu Sha Bay-using stable isotope analysis results[J].Acta Hydrobiologica Sinica,2013,37(1):150-156.
[12] 蔡德陵,李红燕,唐启升,等.黄东海生态系统食物网连续营养谱的建立:来自碳氮稳定同位素方法的结果[J].中国科学(C辑),2005,35(2):123-130.
CAID L,LIH Y,TANG Q S,et al.Establishment of trophic continuum in the food web of the Yellow Sea and East Sea ecosystem:insight from carbon and nitrogen stable isotopes[J].Science in China Ser.C Life Sciences,2005,35(2):123-130.
[13] FUKUMORIK,OIM,DOIH,et al.Bivalve tissue as a carbon and nitrogen isotope baseline indicator in coastal ecosystems[J].Estuarine Coastal and Shelf Science,2008,79(1):45-50.
[14] 余 婕,刘 敏,侯立军,等.崇明东滩大型底栖动物食源的稳定同位素示踪[J].自然资源学报,2008,23(2):319-326.
YU J,LIU M,HOU L J,et al.Food sources of macrofaunal in East Chong Ming saltmarsh as traced by stable isotopes[J].Journal of Natural Resources,2008,23(2):319-326.
[15] 蒋日进,章守宇,王 凯,等.枸杞岛近岸海域食物网的稳定同位素分析[J].生态学杂志,2014,33(4):930-938.
JIANG R J,ZHANG S Y,WANG K,et al.Stable isotope analysis of the offshore food web of Gouqi Island[J].Chinese Journal of Ecology,2014,33(4):930-938.
[16] 周曦杰.枸杞岛典型生境螺贝类代表种—角蝾螺、紫贻贝摄食生态初步研究[D]:上海:上海海洋大学,2014.
ZHOU X J.Preliminary study on feeding behaviour of the representative species in typical habitats(seaweed bed and mussel farm)in Gouqi island:the gastropod,Turbocornutus Solander;the mussel,Mytilus edulis[D].Shanghai:Shanghai Ocean University,2014.
[17] 吴祖立,章守宇,陈 彦,等.枸杞岛海藻场大型底栖无脊椎动物摄食类群研究[J].水产学报,2015,39(3):381-391.
WU Z L,ZHANG SY,CHEN Y,et al.Analysis of functional feeding groups of macroinvertebrates communities in themacroalgae beds of Gouqi island,Zhejiang Province[J].Journal of Fisheries of China,2015,39(3):381-391.
[18] MITCHELL M J,MILLS E L,IDRISI N,et al.Stable isotopes of nitrogen and carbon in an aquatic food web recently invaded by Dreissena polymorpha(Pallas)[J].Canadian Journal of Fisheries and Aquatic Sciences,1996,53(6):1445-1450.
[19] POST D M,LAYMAN C A,ARRINGTON D A,etal.Getting to the fat of thematter:models,methods and assumptions for dealing with lipids in stable isotope analyses[J].Oecologia,2007,152(1):179-189.
[20] GUSTAFSON L,SHOWERSW,KWAK T,et al.Temporal and spatial variability in stable isotope compositions of a freshwatermussel:implications for biomonitoring and ecological studies[J].Oecologia,2007,152(1):140-150.
[21] 余 婕.河口潮滩湿地有机质来源、组成与食物链传递研究[D].上海:华东师范大学,2008.
YU J.Sources,composition and food chain transfer of organicmatter in the estuarine tidal ecosystem[D].Shanghai:East China Normal University,2008.
[22] DOIH,MATSUMASA M,TOYA T,et al.Spatial shifts in food sources for macrozoobenthos in an estuarine ecosystem:Carbon and nitrogen stable isotope analyses[J].Estuarine Coastal and Shelf Science,2005,64(2-3):316-322.
[23] MACHAS R,SANTOS R,PETERSON B.Tracing the flow of organic matter from primary producers to filter feeders in Ria Formosa lagoon,southern Portugal[J].Estuaries,2003,26(4A):846-856.
[24] JENNINGSS,WARR K J.Environmental correlatesof large-scale spatial variation in theδ15N ofmarine animals[J].Marine Biology,2003,142(6):1131-1140.
[25] 曾宴平,马家海,陈斌斌,等.浙江省枸杞岛潮间带大型底栖海藻群落的研究[J].浙江农业学报,2013,25(5):1096-1102.
ZENG Y P,MA JH,CHEN B B,et al.Survey on the community of benthic macro-algae in Gouqi island of Zhejiang Province[J].Acta Agriculturae Zhejiang Gensis,2013,25(5):1096-1102.
[26] 周曦杰,章守宇,王 旭,等.枸杞岛海藻场角蝾螺夏季摄食选择性及其生态学意义[J].水产学报,2015,39(4):511-519.
ZHOU X J,ZHANG S Y,WANG X,et al.The feeding behaviour and ecological function during summer of one herbivore on seaweed bed in Gouqi island:the gastropod,Turbo cornutus Solander[J].Journal of Fisheries of China,2015,39(4):511-519.
[27] 陈明超,章守宇.枸杞岛海域浮游植物群落结构分析[J].上海海洋大学学报,2011,20(4):607-612.
CHEN M C,ZHANG SY.The community structure of phytoplankton in Gouqi island[J].Journal of Shanghai Ocean University,2011,20(4):607-612.
[28] 彭士明,施兆鸿,尹 飞,等.利用碳氮稳定同位素技术分析东海银鲳食性[J].生态学杂志,2011,30(7):1565-1569.
PENG SM,SHIZH,YIN F,etal.Feeding habits of silver pomfret(Pampus argenteus)in East China Sea based on stable isotope techniques[J].Chinese Journal of Ecology,2011,30(7):1565-1569.
Investigation on food web’s isotopic baseline in typical reef ecosystem-seaweed bed
CHEN Ling,WANG Kai,ZHOU Xi-jie,ZHAO Xu,CHEN Liang-ran,ZHANG Shou-yu,WANG Zhen-hua
(College of Marine Sciences of Shanghai Ocean University,Shanghai201306,China)
Kelp bed is a typical rocky inshore ecosystem whose trophic structure ismuch more complicated than the offshore habitats.Pelagic food chain and benthic food chain exist simultaneously,with phytoplankton and macrophytes as their primary production.Recently,more and more stable isotope analysis has been employed to investigate inter and intraspecific relationships between species in the kelp ecosystems.The accurate identification on isotopic baseline species of the food web is of importance in estimating the trophic positions of consumers and evaluating the trophic structure of kelp ecosystem.Aquatic biota samples were collected off the Gouqi Island rocky kelp bed in Jan.and Oct.2014.Based on the survey results of benthic species in 2012,we chose four primary consumer species with large abundance and wide distribution.Two snail species(Turbo petholatus,Turbo cornutus)and two mussel species(Septifer virgatus and Modiolus comptus)were collected in kelp ecosystem,the foot tissues of snail and mussels were used for the stable isotope analysis,respectively.δ13C andδ15N of all species from seaweed field between sampling sites(seasons)were explored to select the isotopic baseline species by multivariate analysis of variance(MANOVA).In this study,snails andmussels exhibited significantly different stable carbon but similar stable nitrogen signatures,suggesting that they had similar trophic position but selectively ingested and assimilated different suspended particulatemixture from littoral and pelagic food chain,respectively.The results showed thatδ13C andδ15N ofmacro-benthoswere affected both by their diets and habitatenvironment.Compared withT.petholatus,we found thatT.cornutusreflected the isotopic signature of the base of the littoral food web.The variation scales inδ15N andδ13C of differentmussels specieswere different.In this sense,using amore stable long-lived primary consumermussel specieswith large abundance to present the pelagic baseline species could facilitate estimating the trophic position of higher trophic level consumers.Our results can provide basic data and important references for the research on other waters of islands in the East China Sea.
stable isotope;seaweed;baseline;macrobenthos
S 931
A
1004-2490(2016)04-0364-10
2015-06-11
国家自然科学基金(NFSC41176110,NFSC41406153)
陈 玲(1990-),女,硕士研究生,研究方向为食物网科学、稳定同位素生态学。E-mail:cl780576862@163.com
王 凯,讲师。E-mail:kwang@shou.edu.cn
