圆口铜鱼幼鱼可持续游泳能力及活动代谢研究
2016-02-26涂志英李丽萍王从锋黄应平
涂志英,李丽萍,袁 喜,王从锋,黄应平
(三峡库区生态环境教育部工程研究中心(三峡大学),湖北宜昌 443002)
圆口铜鱼幼鱼可持续游泳能力及活动代谢研究
涂志英,李丽萍,袁喜,王从锋,黄应平
(三峡库区生态环境教育部工程研究中心(三峡大学),湖北宜昌443002)
摘要:以圆口铜鱼(Coreiusguichenoti)幼鱼为研究对象,通过自制的鱼类游泳实验装置,测定了4个温度(10,15,20和25 ℃)下圆口铜鱼幼鱼游泳速度达到临界游速过程中及运动疲劳后耗氧率的变化情况;在自然水温条件下,测定了5个不同流速下的可持续游泳时间,并通过摄像记录分析了不同游泳速度下的游泳行为。结果显示:圆口铜鱼幼鱼在运动疲劳前耗氧率随流速的增加显著上升(P<0.05),在临界游速时达到峰值;运动疲劳后耗氧率逐渐下降,40~50 min内耗氧率恢复至低流速时的水平。自然水温(18.0±1.5)℃ 条件下幼鱼的可持续游泳时间随流速增加逐渐减小(P<0.01),在1.31倍临界游速的固定流速下,平均可持续游泳时间达29 min,体现了较强的游泳耐力及无氧代谢能力。录像分析表明,摆尾频率(TBF)与游速的关系呈线性正相关(P<0.001),且随着温度升高,TBF随流速增加的趋势越明显。
关键词:圆口铜鱼(Coreiusguichenoti);可持续游泳能力;活动代谢;鱼道
我国在大江大河上修建了许多大坝和堤堰等,虽然其在防洪、灌溉、发电等方面发挥了重要作用,但对鱼类而言,大坝阻碍了部分鱼类洄游到水库上游进行产卵和回归大海,对鱼类的生长繁殖产生了严重的影响,其中以对洄游、半洄游性鱼类的影响最大。大坝造成的洄游通道阻隔、水流速度降低及水温变化等因素的组合影响是鱼类数量下降的主要原因[1-3]。

1材料与方法
1.1实验材料
圆口铜鱼幼鱼为2012年4月上旬购自四川合江县长江船体养殖网箱,其鱼苗每年由长江捕获,实验用鱼为养殖半年左右的幼鱼,体长15.5~19.2 cm,体重49.8~84.9 g。实验鱼在体积2 m3、置于露天环境中的自制水槽中暂养一周。暂养槽水源来自长江,日换水量约为20%,水温昼夜变化15~18 ℃。暂养期间于每日夜间投放养殖饲料一次并连续充氧,保持水槽内溶解氧大于8.0 mg/L。
1.2实验方法
基于长江合江段的年均水温(7~25 ℃),设定了4个实验温度(10、15、20、25 ℃)。圆口铜鱼暂养1周后,按不同驯化温度分为4组,每组10条鱼,然后在设定温度下禁食并适应2 d。同时将游泳实验装置中的水温恒定在实验所需温度下。待鱼驯化结束,将其转移至自行设计的12 L游泳实验装置中。该装置为一个横截面为正方形的循环式水槽,由电机带动桨叶旋转控制水流速,水在整个水槽中循环流动。工作区(45 cm×10 cm×10 cm)上游有一个整流装置,下游侧为不锈钢网,鱼在工作区中顶流游动。循环式水槽又被置于一个50 L(84 cm×39 cm×15 cm)的矩形水槽中,工作区溶氧不足时,可以通过潜水泵将密封循环水槽内的水与矩形槽中的水进行交换,保证工作区中溶解氧的浓度。水槽上方安装一摄像头,对鱼的游泳行为进行监测,实现临界游泳速度(Ucrit)和游泳行为的同步测定。水流速度使用挪威产Vectrino三维点式流速仪测定,水温由加热棒控制,水中氧气浓度(DO)用溶氧仪(HACH,HQ10)测定,使用摄像头(25帧/s)对实验鱼的游泳行为进行录像,以分析其游泳行为。
1.2.1临界游泳速度及耗氧率
将一尾驯化后的鱼放入实验装置中,在1.0 bl/s的流速下适应1 h,消除转移过程对鱼体的胁迫,然后每30 min提高一次流速,流速增量为0.5 bl/s。每个30 min期间,中间的20 min将装置密封,测量三次溶解氧的浓度(DO),间隔时间为10 min。开始的5 min及后期的5 min用于内外水的交换及混合,保证工作区中DO值不低于8.0 mg/L。当鱼疲劳时,将流速调至1.0 bl/s,并继续进行DO的测定,连续测定1 h。鱼疲劳的判定:鱼停靠在下游网上时,轻拍下游壁面20 s,鱼仍不重新游动,视为疲劳。实验结束后取出实验鱼并测量体重及常规形态学参数。相对临界游泳速度(Ucrit,bl/s)按以下公式计算:
(1)
其中U为实验鱼能完成间隔时间(△t)的最大水流速度,t为在最高流速下游泳的时间(min),△t为改变流速的时间间隔(30 min),△U为速度增量(1.0 bl/s)。
由于鱼类进行有氧代谢时,放出的热量与消耗的氧成正比,所以本研究通过测定耗氧率(MO2,mg O2kg-1h-1)来间接计算鱼类在单位时间内的全部能量消耗,也称为代谢率。耗氧率的计算公式如下:
(2)
V为游泳装置体积(L),d(DO)/dt为某流速下DO值随时间变化的斜率(mgO2L-1h-1),M为鱼的湿重(kg)。
鱼体的截面积会引发阻挡效应,但当实验鱼的最大横截面积小于10%的游泳区截面积时,测定的临界游泳速度可以不用校正[8]。本研究的实验鱼鱼体最大截面积均小于10%的游泳区截面积。空白溶氧值的变化经测定小于1%,可以忽略。
1.2.2可持续游泳时间
根据20 ℃条件下临界游泳速度的研究结果,设定了6.5、7.0、7.5、8、8.5 bl/s 五个流速,每一流速条件设8个平行。将单尾实验鱼置于游泳实验装置中,在自然水温条件下适应1 h,适应期间流速为1.0 bl/s,然后每隔5 min增加一次流速,流速增量为0.5 bl/s,增加至设定流速后开始计时,观察鱼的游泳行为,直至实验鱼疲劳无法继续游动,结束实验并记录游泳时间。
1.2.3游泳行为的测定
在测量临界游速的过程中,同时用摄像头(25 frames/s)记录鱼在流速增加过程中的游泳行为变化。通过视频慢放记录鱼在某流速下1 min内的摆尾次数(TBF)。
1.3统计方法
实验数据用spss11.5软件进行统计分析,采用单因素方差分析(one-way ANOVA),并利用SNK多重比较法比较各组差异。统计值用平均数±标准差(Mean±SD)描述,差异显著性水平为P<0.05。相关性分析用Pearson相关性分析。
2结果
2.1不同温度下临界游速的变化特征
在不同水温(10、15、20、25 ℃)条件下,测定圆口铜鱼幼鱼的临界游泳速度(Ucrit)发现,实验鱼的临界游速随着温度的升高而增大(表1),将临界游速与水温T之间拟合可得到线性方程:Ucrit=3.86+0.14T(R2=0.99,P<0.001)。
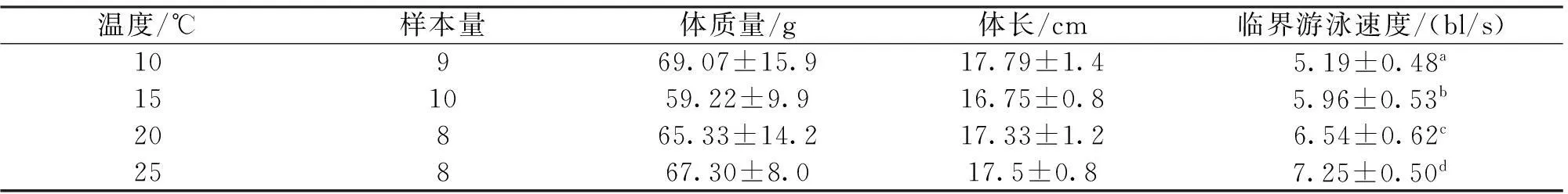
表1 不同水温条件下圆口铜鱼的临界游泳速度
注:数值为平均数±标准偏差,a,b,c,d表示不同水温下的临界游速具有显著差异。
2.2温度及流速对耗氧率的影响
在不同温度条件下的临界游速测定过程中,当流速较低时(2.5~3.0 bl/s),耗氧率(MO2)随流速变化不明显,但随着流速的增加MO2显著增加,直至接近临界游速时达到最大值,且随着温度增加,耗氧率随流速增加的趋势越明显(图1)。4个温度条件下的最大耗氧率分别为(285.6±24.3)、(403.1±20.8)、(453.0±23.7)、(550.3±96.3) mgO2kg-1h-1。耗氧率与水温及游速之间的相关性分析如表2所示,三者具有显著相关性。运动疲劳后的耗氧率称为运动后过量耗氧(EPOC,mg O2kg-1h-1),游泳疲劳后的EPOC值迅速下降,约40~50 min内恢复至低流速下的耗氧率水平。

表2 水温(T)、临界游速(Ucrit)及最大耗氧率
**表示在0.01水平上显著(双尾检验)。

图1 不同温度条件下Ucrit测定过程中及运动疲劳后
2.3不同流速条件下的可持续游泳时间
自然水温条件下(18.0±1.5 ℃),圆口铜鱼幼鱼的可持续游泳时间随流速的增加逐渐减少,但个体差异较大(表3)。在8.5 bl/s(约1.31Ucrit)的固定流速下,平均可持续游泳时间达到29 min。
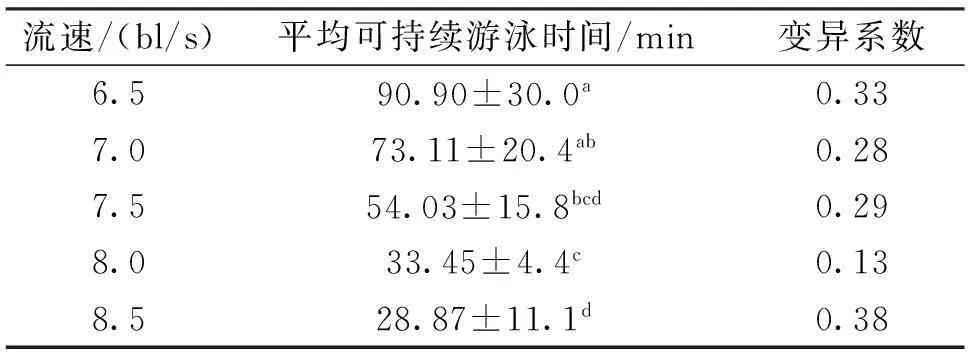
表3 可持续游泳时间变异系数(C.V)
注:数值为平均数±标准偏差,相同字母表示差异不显著。
2.4不同流速下的游泳行为
在1.0 bl/s 的流速下,圆口铜鱼表现出正趋流性,通过尾鳍的偶尔摆动静止于水槽某一位置。当流速增加至3.0 bl/s左右,鱼的尾鳍开始连续摆动且摆尾频率(TBF)随流速增加而逐渐增加。当流速接近临界游速时,尾鳍又开始不连续摆动,出现冲刺-滑行的游动方式,使身体固定在游泳区的上游处;偶尔尾抵下游时,便通过急速摆尾向前冲刺。随着流速继续增加,鱼蜷缩在下游停止游动。视频分析表明,4个温度下的摆尾频率均随流速增加而增大,且温度越高,摆尾频率随流速增加的幅度越大(图2)。低游速(1.5~2.5 bl/s)条件下,不同水温的TBF差别较小,尤其在水温较低时(10~15 ℃),此游速范围内的TBF值几乎重合。

图2 不同温度下摆尾频率(TBF)随流速的变化
3讨论
3.1温度对临界游泳速度的影响
临界游泳速度是评价鱼类最大有氧运动能力的指标之一,也是鱼道内过鱼孔口流速大小设计的主要依据。水温对鱼类有氧游泳行为有较大影响:温度过高,血液运输氧的能力降低,限制了氧气向组织中的传递;温度下降,血红蛋白对氧气的亲和力加强,但肌肉的收缩力却下降。多数研究结果显示鱼类的临界游泳速度(Ucrit)与温度的相互关系呈现先上升再下降的“钟形”变化趋势[9],即温度低于或超过最适宜温度都会导致临界游泳速度的下降。但也有研究表明两者为单调线性上升的趋势[8],与本研究结果一致。原因可能是在20~25 ℃的水温条件下,圆口铜鱼的临界游速还未达到最大值。据文献报道,温水性鱼类最大临界游速大多出现在20 ℃以上。如乌颊鱼(Sparusaurata)在25 ℃时临界游速达到最大[10],黑鲈(Dicentrarchuslabrax)在24.4 ℃时达到最大临界游速[11]。南方大口鲶(Silurusmeridionalis)的最大临界游速对应温度为28.4 ℃[9]。当水温越接近鱼类的最适生长、繁殖等生命活动的温度时,游泳能力随温度的变化幅度越小,越偏离这个温度,变化幅度越大,由此可推断圆口铜鱼幼鱼的运动最适温度为15~25 ℃之间,且在此温度范围内临界游泳速度达到最大。
3.2流速及温度对耗氧率的影响
当流速小于2.5~3.0 bl/s时,耗氧率随流速的变化均不明显(图1),尤其是在10 ℃的条件下。估计是由于低游泳速度下,鱼主要通过胸鳍和背鳍的补偿运动来维持平衡,故能量消耗较小。白鲑(Coregonusalbula)维持代谢率不变的游速为1 bl/s以下[14]。而圆口铜鱼在如此大的游速范围内维持能量代谢基本不变,表明圆口铜鱼在游泳过程中,产生推进力所消耗的能量较低,这是其长期栖息于急流环境而形成的适应性。

表4 四种鲤科鱼类临界游速(Ucrit)及最大耗氧率(MO2,max)的比较
运动疲劳后能量代谢恢复至稳定耗氧率的时间反映了短期内鱼类的再次运动能力,对鱼的生存与繁殖具有重要的意义,快速的恢复有利于鱼类能适时地洄游至目的地。圆口铜鱼在运动疲劳后的耗氧率(EPOC)迅速下降,可能是达到临界游速之前,能量来源主要是有氧代谢,无氧代谢的比例较低[16]。游泳疲劳后约40~50 min内基本可恢复至低游泳速度下的耗氧率水平,恢复时间随温度的升高出现先上升后下降的特点(图1),与南方鲶(Silurusmeridionalis)幼鱼力竭运动后的恢复时间随温度变化规律基本一致[17],表明温度对圆口铜鱼的运动生理代谢具有较显著的影响。不同鱼类力竭运动后代谢恢复时间也不尽相同。大西洋鳕(Gadusmorhua)在临界游泳速度测试力竭后的恢复时间为1.5 h[16],太平洋鲑鱼(Oncorhynchussp.)的恢复时间为45 min[18],成年野生的红鲑(Oncorhynchusnerka)和银鲑(O.kisutch)的恢复时间约为42~78 min,红鲑的恢复时间随温度的升高而加长[19]。目前监测鱼类运动疲劳运动后耗氧率的时间多釆用40~60 min。为了更好地评价和反映运动疲劳后鱼类的生理代谢过程及其代谢率恢复,还应结合常规生理生化指标(乳酸、糖原和葡萄糖)的连续监测。
3.3可持续游泳时间
对于距离较长或者结构较为简单的鱼道,持续游泳时间关系到鱼类可以连续上溯的距离,是鱼道长度以及休息池设计的参考依据之一。随着游泳速度的增加,圆口铜鱼幼鱼的可持续游泳时间逐渐下降,其中在8.5 bl/s(1.31Ucrit)的固定流速下(18.0±1.5 ℃),平均可持续游泳时间约为29 min(表3)。鲜雪梅等[20]测定了青鱼(Mylopharyngodonpiceus)、锦鲫(Carassiusauratus)及南方鲇(Silurusmeridionalis)在1.5Ucrit(25 ℃)条件下的平均可持续游泳时间分别为8.23 、1.07、66.36 s;Zeng等[9]测定了在温度25 ℃,流速1.23Ucrit条件下,南方鲇的可持续游泳时间为220 s。根据圆口铜鱼游泳能力随温度增加而增大(表1),可推测圆口铜鱼在1.5Ucrit(25 ℃)条件下的可持续游泳时间大于青鱼、锦鲫及南方鲇。圆口铜鱼具有较强的游泳耐力应与其细长的身体形态有关:圆口铜鱼、青鱼、锦鲫及南方鲇丰满度分别为0.75、1.64、3.03、0.76[20];此外还与圆口铜鱼具有较长距离的洄游习性有关[5],使其具有较强的游泳耐力及无氧代谢能力。
本研究的实验数据与国外学者得到的可持续游泳时间与游泳速度的数学模型[21]相差甚远。这是由于可持续游泳时间的测试方法不同,如测试前适应时间的长短、如何由适应时的低流速增加到指定流速、疲劳的判断、测试的装置等,导致实验结果差别很大,故得到的经验公式难于普遍适用。
3.4不同流速下的摆尾频率
鱼类游泳的行为特征,如摆尾频率(TBF)及摆尾幅度(TBA),亦可为鱼道设计提供参数[22]。摆尾频率与游泳速度的关系还可用于野外实验研究,通过易于监测的摆尾频率对难于测定的游泳速度[23]及自由游动下的活动代谢率[24]进行估算。有学者得出摆尾频率与游速之间满足幂函数关系[23],而更多的研究则表明两者为线性关系,但这种线性关系存在明显的种间差异。圆口铜鱼幼鱼的摆尾频率与流速变化呈直线关系(图2),在10~25 ℃条件下,直线斜率在0.11~0.42范围内。同为鲤科的台湾突吻鱼(Onychostomabarbatula)TBF与流速关系的斜率为0.59[22],鲤(cyprinuscarpio)为1.31,拟鲤(Rutilusrutilus)为1.30[24],都显著高于圆口铜鱼,说明在相同游泳速度条件下,圆口铜鱼将消耗更少的能量。
实验研究表明,圆口铜鱼是一种游泳能力较强的鱼类,幼鱼的最大绝对临界游速接近8.0 bl/s,且在1.31Ucrit的固定流速下,最大可持续游泳时间可达37 min,这与其游动过程中较高的能量利用效率有关,也与其纺锤形形态、喜急流及长距离洄游的生态特征相适应。
参考文献:
[1]柯福恩,危起伟,罗俊德,等.三峡工程对长江渔业资源的影响与补救措施[J].淡水渔业,1994,24(1):6-9.
[2]张志英,袁野.溪落渡水利工程对长江上游珍稀特有鱼类的影响探讨[J].淡水渔业,2001,31(1):62-63.
[3]吴强,段辛斌,徐树英,等.长江三峡库区蓄水后鱼类资源现状[J].淡水渔业,2007,37(2):70-75.
[4]刘乐和,吴国犀,王志玲.葛洲坝水利枢纽兴建后长江干流铜鱼和圆口铜鱼的繁殖生态[J].水生生物学报,1990,14(3):205-215.
[5]Park Y S,Chang J B,Lek S,et al.Conservation strategies for endemic fish species threatened by the Three Gorges Dam[J].Conserv Biol,2003,17(6):1748-1758.
[6]于丽娟,李秀明,易建华,等.力竭追赶训练对中华倒刺鲃自由基清除能力的影响[J].淡水渔业, 2013,43(3):28-32.
[7]彭姜岚,曹振东,付世建.鲇鱼力竭性运动后的过量耗氧及其体重的影响[J].水生生物学报,2008,32(3):380-386.
[8]Jain K E,Farrell A P.Influence of seasonal temperature on the repeat swimming performance of rainbow troutOncorhynchusmykiss[J].J Exp Biol,2003,206:3569-3579.
[9]Zeng L Q,Cao Z D,Fu S J,et al.Effect of temperature on swimming performance in juvenile southern catfish (Silurusmeridionalis) [J].Comp Biochem Physiol A,2009,153:125-130.
[10]Koumoundouros G,Ashton C,Xenikoudakis G,et al.Ontogenetic differentiation of swimming performance in Gilthead seabream (Sparusaurata,Linnaeus 1758) during metamorphosis[J].J Exp Mar Biol Ecol,2009,370:75-81.
[11]Koumoundouros G,Sfakianakis D G,Divanach P,et al.Effect of temperature on swimming performance of sea bass juveniles[J].J Fish Biol,2002,60:923-932.
[12]Tudorache C,Viaene P,Blust R,et al.A comparison of swimming capacity and energy use in seven European freshwater ?sh species[J].Ecol Freshw Fish,2008,17:284-291.
[13]Behrens J W,Prbel K,Steffensen J F.Swimming energetics of the Barents Sea capelin(Mallotusvillosus)during the spawning migration period[J].J Exp Mar Biol Ecol,2006,331(2):208-216.
[14]Ohlberger J,Staaks G,Hölker F.Effects of temperature,swimming speed and body mass on standard and active metabolic rate in vendace(Coregonusalbula)[J].J Comp Physiol B,2007,177(8):905-916.
[15]Li D,Hu W,Wang Y,et al.Reduced swimming abilities in fast-growing transgenic common carpCyprinuscarpioassociated with their morphological variations[J].J Fish Biol,2009,74(1):186-197.
[16]Reidy S P,Nelson J A,Tang Y,et al.Post-exercise metabolic rate in Atlantic cod and its dependence upon the method of exhaustion[J].J Fish Biol,1995,47(3):377-386.
[17]曾令清.温度对南方鲇幼鱼游泳能力和静止代谢率的影响[D].重庆:重庆师范大学,2008.
[18]Farrell A P,Lee C G,Tierney K,et al.Field-based measurements of oxygen uptake and swimming performance with adult Pacific salmon (Oncorhynchussp.) using a mobile respirometer swim tunnel[J].J Fish Biol,2003,62(1):64-84.
[19]Lee C G,Farrell A P,Lotto A,et al.Excess post-exercise oxygen consumption in adult sockeye (Oncorhynchusnerka) and coho (O.kisutch) salmon following critical speed swimming[J].J Exp Biol,2003,206:3253-3260.
[20]鲜雪梅,曹振东,付世健.4种幼鱼临界游泳速度和运动耐受时间的比较[J].重庆师范大学学报,2010,27(4):16-20.
[21]Castro S T.Optimal swim speeds for traversing velocity barriers:an analysis of volitional high-speed swimming behavior of migratory fishes[J].J Exp Biol,2005,208(3):421-432.
[22]Lin P J,Ni I H,Huang B Q.Evaluation of the swimming ability of wild caughtOnychostomabarbatula(Cyprinidae) and applications to fishway design for rapid streams in Taiwan[J].Raffles B Zool,2008,19:273-284.
[23]Steinhausen M F,Steffensen J F,Andersen N G.Tail beat frequency as a predictor of swimming speed and oxygen consumption of saithe (Pollachiusvirens) and whiting (Merlangiusmerlangus) during forced swimming[J].Mar Biol,2005,148(1):197-204.
[24]Ohlberger J,Staaks G,Hölker F.Estimating the active metabolic rate (AMR) in fish based on tail beat frequency and body mass[J].J Exp Zool,2007,307A(5):296-300.
(责任编辑:邓薇)
Endurance swimming capability and activity metabolism of
Coreius guichenoti juvenile
TU Zhi-ying,LI Li-ping,YUAN Xi,WANG Cong-feng,HUANG Ying-ping
(EngineeringResearchCenterofEco-environmentinThreeGorgesReservoirRegion,
MinistryofEducation,ChinaThreeGorgesUniversity,Yichang443002,Hubei,China)
Abstract:To obtain data that can be applied to the design of an effective fishway forCoreiusguichenoti,and other species in the fish community,a laboratory study of juvenileC.guichenoti’s swimming ability and energetics was conducted in a flume-type respirometer.The oxygen consumption rate (MO2)before and after swimming fatigue were determined during steady swimming to reach the critical swimming speed (Ucrit) at four water temperatures (10 ℃,15 ℃,20 ℃ and 25 ℃).The endurance swimming time at five swimming speeds was determined at natural water temperature.The swimming behavior was recorded with a high speed video camera system during theMO2protocol.Before swimming fatigue,MO2increased with the increase of swimming speed and reached the maximum at theUcrit.Following exhaustion atUcrit,recovery time was 40~50 min.Results show that temperature has a significant effect on swimming performance.The endurance swimming time decreased with the increase of flow velocity at 18.0±1.5 ℃ (P<0.01) but reach 29 min at 1.31Ucrit,showing high swimming endurance and anaerobic metabolic capacity.There was a significant positive linear correlation between tail beat frequency (TBF) and swimming speed (P<0.001) and the increase is more significant as the temperature is higher.
Key words:Coreiusguichenoti;endurance swimming capability;active metabolism;fishway
中图分类号:S917.45
文献标识码:A
文章编号:1000-6907-(2016)01-0033-06
作者简介:第一涂志英(1973-),女,博士,副教授,主要从事水污染及水生态研究。E-mail:zytu1111@163.com通讯作者:黄应平。E-mail:yingpinghuang@126.com
收稿日期:2014-08-24;
修订日期:2015-07-26
资助项目:国家水专项子课题(2012ZX07104-003-04);国家自然科学基金(51309140);三峡大学科研启动基金(0620120138)
