兰寿金鱼与虎头金鱼形态性状差异分析
2016-02-26陈再忠
周 祺,张 帅,谢 松,陈再忠
(上海海洋大学水产与生命学院,上海 201306)
兰寿金鱼与虎头金鱼形态性状差异分析
周祺,张帅,谢松,陈再忠
(上海海洋大学水产与生命学院,上海201306)
摘要:对金鱼(Carassiusauratusauratus)中兰寿金鱼与虎头金鱼两大不同品系的 28个性状进行了形态学测量。通过统计学分析,结果显示:两种样品主要在体高、体宽和头长3项指标上具有差异,其主成分分析特征值均在0.7以上。另外,鱼体外部各性状之间联系十分密切。其中11项形态参数在经聚类分析与近似性矩阵分析后,获得了由相关性距离构成的环状系统。该系统中的一个或多个性状特征的改变都会对其他性状参数造成不同影响,并进一步影响其鱼体外形和器官功能。
关键词:兰寿金鱼(Carassiusauratusauratus);虎头金鱼;形态学
金鱼(Carassiusauratusauratus)是我国传统的观赏鱼养殖品种,其短暂的驯化史中产生了包括背鳍在内14种外部结构变异。对其各个层面的研究自20世纪至今从未停止[1-3]。但因其各个品种间均不存在生殖隔离,后代性状表达也极不稳定,目前对于金鱼品种的分类均是以外部形态特征作为基础,且以陈桢[4]对金鱼外形变异的描述作为依据。在王春元[5]创建金鱼的分类系统中,同样以性状作为分类的标准,该标准与进化系统有着密切的关系,但又有所区别,其揭示了部分金鱼可能的进化规律,但并非完全反映进化系统。这符合数值分类学的基本思想,但缺乏与可量数据间的结合,在金鱼进化或发育方面无法得出更加准确的结论。
作为蛋种金鱼中最具代表性的两个品种,虎头与兰寿,虎头金鱼原产于中国,而兰寿金鱼原产于日本,两者极易混淆,唯一区分方式是兰寿背后部弯曲成弓形,而虎头则保留正常金鱼背部平滑的特征[6]。本研究通过对两者的表型可测性状进行统计学分析,结合金鱼进化与发育从不同角度解析兰寿与虎头差异的原因,以期为今后相关的研究提供基础。
1材料与方法
1.1材料
实验用鱼取自上海浦东观赏鱼中心,其中兰寿、虎头各2组,共122尾(表1),确保其性状发育完整,各性状特征均符合系统分类学中对其的描述(图1)。实验用鱼均为1龄。

表1 虎头与兰寿金鱼样品

图1 实验用鱼
1.2方法
1.2.1形态学测量
选取27个可测量性状,对各测量参数分类编号,其中长度指标通过游标卡尺测量,包括体长、体宽等;重量指标通过电子天平测量;角度测量以量角器完成。每尾鱼的各个形态性状均测3次,最终性状参数取其平均值。所测指标见表2。
由于不同金鱼个体间存在大小差异较大,故针对长度性状观测值进行如下处理:
最终个体性状参数对应=个体性状观测值/个体全长
通过SPSS 18.0 软件的ANOVA模块及T检验模块对所检测的同类金鱼的各指标进行组内、组间显著性差异检测。
进行聚类分析、主成分分析及回归分析。标准化(STD)处理后,对27个性状进行R型聚类分析。再采用主成分分析,最后通过回归分析检测各性状之间关系。
2结果
2.1兰寿金鱼与虎头金鱼差异性状主成分分析
由主成分分析结果(表2)可知,前7个主成分的总贡献率为68.763%。第1主成分的贡献率为23.686%,其中影响最大的是体高、体宽和头长,其特征值均在0.750以上;尾柄与尾鳍夹角、眼下距、吻至腹鳍基部长度、吻至臀鳍基部长度、臀鳍长和头瘤长也具有较大影响,其特征值均接近0.600。第2主成分的贡献率为16.079%,其中影响较大的主要是上叶尾长,达0.741,体长次之,为-0.592。第3主成分的贡献率为9.405%,影响较大的性状是头宽,达0.779。第4-7主成分的贡献率分别为6.122%、5.455%、4.119%、3.897%,对其影响较大的性状依次是下叶尾宽、吻长、上叶尾宽和尾柄高。

表2 金鱼27个性状指标的主成分特征值、贡献率和累计贡献率
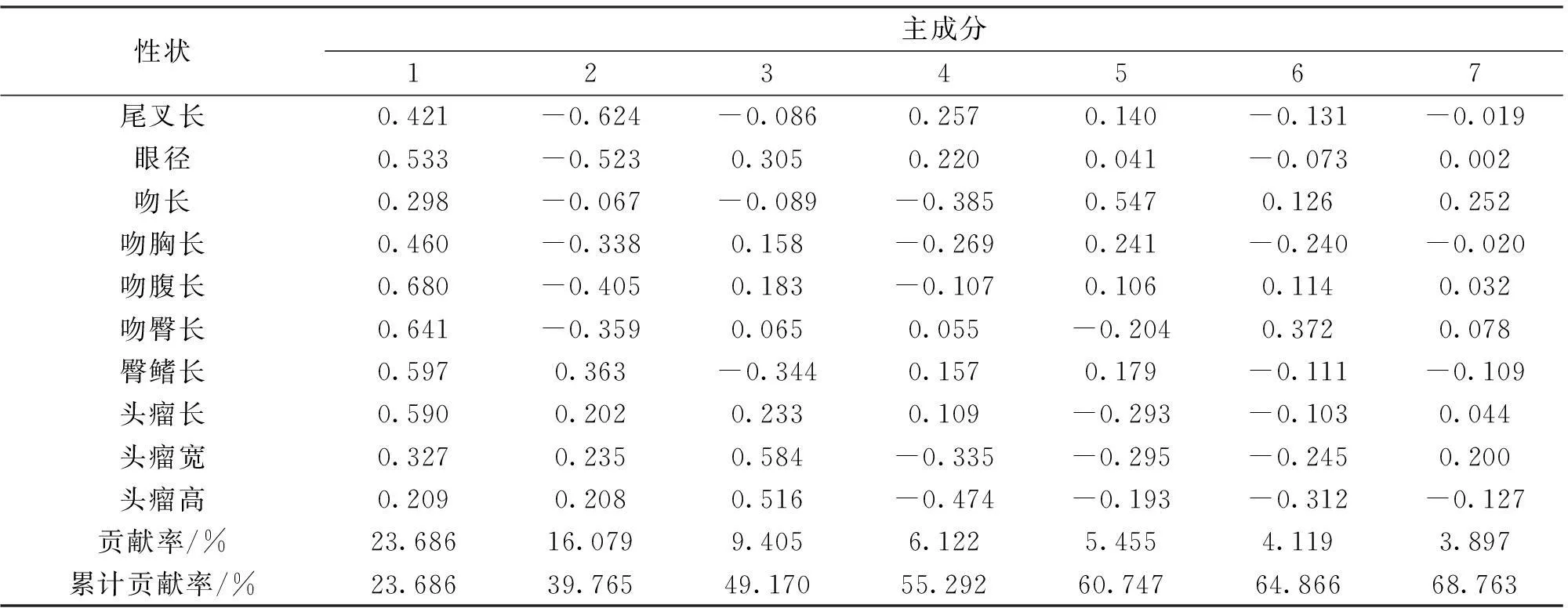
续表2
2.227个性状指标的R型聚类分析
由R型聚类输出的树形图(图2)可知,针对兰寿金鱼与虎头金鱼的27个金鱼性状可分为3个大类,如表3。结合聚类分析及其近似性矩阵,可以发现其多对性状参数间均存在相关性(r>0.5),且各类性状参数之间相关性联系非常紧密(如图3)。

图2 R型聚类输出
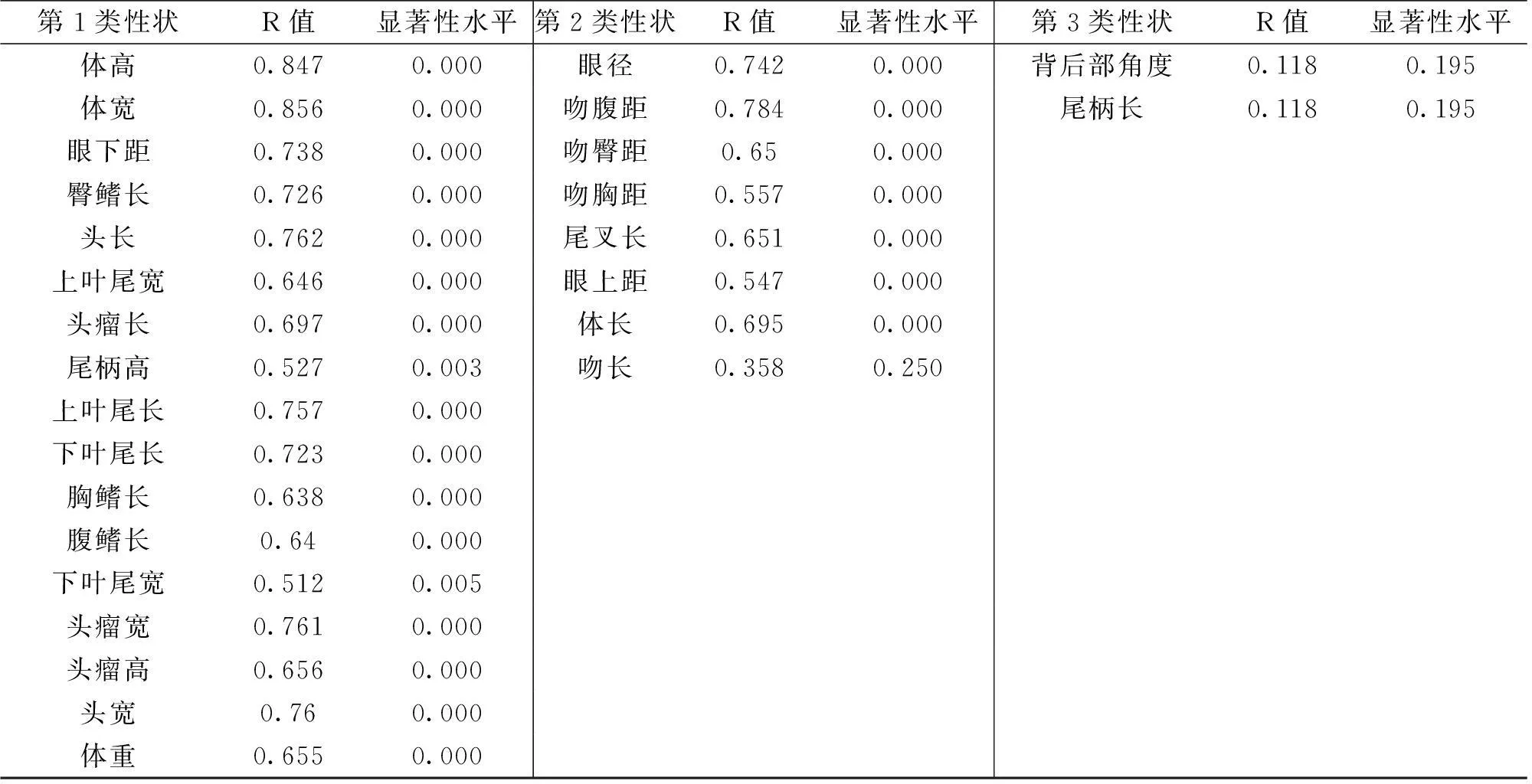
第1类性状R值显著性水平第2类性状R值显著性水平第3类性状R值显著性水平体高0.8470.000眼径0.7420.000背后部角度0.1180.195体宽0.8560.000吻腹距0.7840.000尾柄长0.1180.195眼下距0.7380.000吻臀距0.650.000臀鳍长0.7260.000吻胸距0.5570.000头长0.7620.000尾叉长0.6510.000上叶尾宽0.6460.000眼上距0.5470.000头瘤长0.6970.000体长0.6950.000尾柄高0.5270.003吻长0.3580.250上叶尾长0.7570.000下叶尾长0.7230.000胸鳍长0.6380.000腹鳍长0.640.000下叶尾宽0.5120.005头瘤宽0.7610.000头瘤高0.6560.000头宽0.760.000体重0.6550.000

图3 组间近似性距离r>0.5的金鱼性状图
由此图可以发现,虎头金鱼与兰寿金鱼的之间的差异并非单一存在的,是连续而具有整体性的。
2.3性状参数间回归分析
经过聚类分析,每类中参数过多,通过回归线性结果筛选可以发现,第1类中可复相关系数R值最高者依次为体宽、体高,均达到0.8以上,且在ANOVA中回归模型的概率为0,小于显著性水平0.05或0.01,表明其线性模型是成立的,也就意味着其参数可以作为该分类组中的代表;在第2类中,以吻腹距(R=0.784)最为适合作为代表;第3类中仅有两个参数指标,经显著性检测其线性模型不成立。
3讨论
虎头金鱼和兰寿金鱼作为金鱼中的代表品种,极易被人混淆。通过研究发现,两者不仅在背部与尾鳍夹角角度上具有显著性差异,在多个其他性状特征上亦存在区别之处。同一时间,整个鱼体一个或多个性状特征的改变都会对其他性状参数造成不同影响,进一步影响其鱼体外形和器官功能。这与两种金鱼进化过程中受到的人工选择及环境影响密不可分。
金鱼本由中国大陆野生金鲫演化而来,其外部结构变异中背鳍的消失是重要的一环。本次对无背鳍金鱼的研究发现,27个性状参数聚类所得的三组性状中,体高、吻腹距和尾柄尾鳍间夹角可分别作为各组代表性性状参数,可将其作为识别虎头金鱼与兰寿金鱼的主要特征。同时各个性状参数间的相关性也反映出,两者在外部特征中存在差异但不影响整体性,这显然源于长期的人工选择与鱼体自身功能适应。金鱼属于奇鳍/对鳍推进模式(MPF:Median and/ or paired fin)[7],结合其11个性状所构成的环状系统,我们可以假设兰寿金鱼是在虎头金鱼的基础上进一步变异而来,其原本平坦的背部发生拱起,导致体长变短,为了适应这一改变,鱼体其他部分做出回应。其中尾鳍是金鱼在水中前进时转向与动力的主要来源[8],因此为了改善背部弯曲所加剧的鱼体不稳定性,尾鳍上扬,尾柄与尾鳍夹角变小,尾鳍上尾叶长增长,增加动力与转向稳定性。同时背后部的拱起,造成内脏质量重心后移,鱼体背腹重心得以进一步协调,也造成了体宽增大;头部吻部区域作为鱼体游泳状态下水平方向,其垂直位置的改变也是为了更好的适应前进过程中质量重心改变所带来的流体动力学障碍[9]。
在自然状态下,物种新性状的形成以及机体各器官之间互相适应是一个漫长的过程[10]。但金鱼作为短期内出现大量性状变异的物种,其原因包括以下方面。
首先,金鱼最早经海外贸易之路传至日本,从而产生了一定程度的地理隔离。虽然有研究从分子水平证明,兰寿金鱼祖先仍为中国鲫鱼种群中的一支或几支[11-12]]。但结合适应辐射现象[13-14],我们可以推测,虎头金鱼与兰寿金鱼在进化过程中受到的环境影响、本地鲫属影响均会存在一定差异。使其后代个体在性状表达上出现更多不稳定性,表现出了更多的个体间差异性。
另外,研究显示,金鱼的染色体倍性并不稳定,部分基因或基因簇较相似物种出现冗余拷贝现象[15]。根据脊椎动物多倍体祖先二倍体化(2R)学说,这显示其有可能并未完成基因组加倍事件[16]。这为金鱼提供了比一般鱼类更加充分的突变机会。最后,金鱼作为观赏性品种而存在,在剧烈的人工选择作用下,其奠基者效应与瓶颈效应加剧,大大缩短了其性状发生突变到稳定的时间。
参考文献:
[1]傅毅远.关于我国金鱼品种演化及系统分类的初步意见[J] .淡水渔业,1981,11(6):15-18.
[2]牟希东,白俊杰,汪学杰,等.金鱼核糖体转录间隔区(ITS-1) 的克隆和序列分析[J].淡水渔业,2008.38(1):74-76.
[3]曹海鹏,刘丽玲,李怡,等.金鱼致病菌的分离鉴定与药敏特性[J].淡水渔业,2011,41(6):46-49.
[4]陈桢,金鱼的变异与天演[M].北京:科学出版社.1925.
[5]王春元,我国金鱼品种的分类与命名[J].淡水渔业,1984,(4):30-33.
[6]Matsubara S.Goldfish and Their Culture in Japan:CarassiusAuratus[M].1910:US Government Printing Office.
[7]沈林成,王光明.仿鱼长鳍波动推进器研究的进展与分析[J].国防科技大学学报,2005,27(4):96-100.
[8]李丹,林小涛,朱志明.不同流速下杂交鲟幼鱼游泳状态与活动代谢研究[J].水生生物学报,2011, 35(4):578-585.
[9]周澍欣,胡庆松.鲤鱼三维建模及流场受力数值计算[J].上海海洋大学学报,2013,22(6):855-861.
[10]段昌群,王焕,姜汉侨.污染条件下生物多样性丧失的生态遗传学机制[J].生态科学进展,2004,1:267-290.
[11]胡彩霞,陈再忠,王成辉,等.金鱼的微卫星标记开发[J].江苏农业科学,2012,40(8):22-25.
[12]王姝妍.古环境变化和人工选择对鲫属鱼类遗传结构的影响[D].中国科学技术大学,2013.
[13]Rainey P B,Travisano M.Adaptive radiation in a heterogeneous environment[J].Nature,1998,394(6688):69-72.
[14]Schluter D,Price T,Moores A,et al.,Likelihood of ancestor states in adaptive radiation[J].Evolution,1997,S1(6):1699-1711.
[15]Luo,J,Stadler P F,He S P,et al.,PCR survey of Hox genes in the goldfishCarassiusauratusauratus[J].J Exp Zoo Part B:Mole Develop Evol,2007,308(3):250-258.
[16]Meyer A,Peer Y V D.From 2R to 3R:evidence for a fish‐specific genome duplication (FSGD)[J].Bioessays,2005,27(9):937-945.
(责任编辑:邓薇)
Analysis on the morphology difference of Ranchu goldfish and
tiger head goldfish
ZHOU Qi,ZHANG Shuai,XIE Song,CHEN Zai-zhong
(CollegeofFisheriesandLifeSciences,ShanghaiOceanUniversity,Shanghai201306,China)
Abstract:The experiment focused on 28 different characteristics of Ranchu goldfish and tiger head goldfish.From the result,two kinds of samples had different characteristics in body height,body width and head length.the value of principal component analysis were more than 0.7.In addition,we get a cyclic system composed by 11 morphological parameters after the R cluster and regression equation analysis.One or more characters in the system could affect the other characters,and further affect the shape and function of the fish.
Key words:Ranchu goldfishCarassiusauratusauratus;tiger head goldfish;morphology
中图分类号:S932.4
文献标识码:A
文章编号:1000-6907-(2016)01-0046-05
作者简介:第一周祺(1989-),男,硕士研究生,专业方向为水产养殖。E-mail:zhouqi95@hotmail.com通讯作者:陈再忠。E-mail:chenzz@shou.edu.cn
收稿日期:2015-03-26;
修订日期:2015-10-22
资助项目:沪农科种字(2012)第9号;上海海洋大学水产动物遗传育种协同创新中心(ZF1206)
