双台子河口刀鲚个体生殖力
2016-02-07霍堂斌郑文军李耕赵荣伟代尚英那文学
霍堂斌,郑文军,李耕,赵荣伟,代尚英,那文学
(1.中国水产科学研究院黑龙江水产研究所,农业部黑龙江流域渔业资源与环境重点野外科学观测试验站,黑龙江 哈尔滨150070;2.中国水产科学研究院营口增殖实验站,辽宁 营口 115004;3.哈尔滨市水产技术推广站,黑龙江 哈尔滨 150076;4.东北农业大学,黑龙江 哈尔滨 150040)
双台子河口刀鲚个体生殖力
霍堂斌1,郑文军2,李耕2,赵荣伟3,代尚英4,那文学4
(1.中国水产科学研究院黑龙江水产研究所,农业部黑龙江流域渔业资源与环境重点野外科学观测试验站,黑龙江 哈尔滨150070;2.中国水产科学研究院营口增殖实验站,辽宁 营口 115004;3.哈尔滨市水产技术推广站,黑龙江 哈尔滨 150076;4.东北农业大学,黑龙江 哈尔滨 150040)
于2015年5~6月在辽河双台子河口水域采捕176尾性腺发育到Ⅳ期的刀鲚Coilia nasus雌鱼,测量其体长(L)、体质量(W)、净体质量(Wo)、性腺重(W1)和卵径等生物学指标。取左侧鳃盖骨鉴定年龄,用重量法计数个体绝对生殖力(F),计算体长相对生殖力(FL)和体重相对生殖力(Fw),用5种数学模型及多元逐步回归方程拟合个体生殖力与生物学指标的关系。结果显示:样本由4+~6+龄组成,个体绝对生殖力(F)17 968~116 935粒,平均44 995粒;体长相对生殖力(FL)66~297粒/mm,平均为145粒/mm;体质量相对生殖力(Fw)192~710粒/g,平均为362粒/g。F和FL与体长(L)、体质量(W)、净体质量(Wo)、性腺重(W1)、成熟系数(GSI)和年龄呈极显著正相关,与肥满度(K)相关关系不显著。Fw仅与W1、GSI极显著正相关,和其他指标的相关性不显著。多元逐步回归显示,F与W和W1密切相关,相关式为:F=-15623.878+1158.377Wl+324.104W(n=176,R=0.851;DW=1.890,P<0.01);FL与W1和K密切相关,相关式为:FL=-68.850+3.868Wl+302 687.076K(n=176,R=0.728;DW=1.743,P<0.01);Fw均与L和GSI密切相关,相关式为:Fw=207.215-5.17L+7.5918GSI(n=176,R=0.646;DW=1.849,P<0.01)。第Ⅳ~Ⅴ期刀鲚卵巢中,卵径变化范围在0.55~1.05mm之间,平均0.81mm。依据卵径频率分布推测,刀鲚属单峰一次性产卵鱼类。
刀鲚;个体生殖力;双台子河口
刀鲚Coilia nasus隶属于鲱形目Clupeiformes鳀科Engraulidae鲚属Coilia,俗称刀鱼、凤尾鱼,是辽河名贵的洄游性鱼类,在我国的东海、黄海、渤海和通海的江河均有分布[1-5]。在每年繁殖季节,成熟刀鲚集群分批上溯进入通海的江河。随着环境变化、人类干扰等诸多因素影响,刀鲚资源量逐渐下降,目前已处于濒危状态,1998年列入《中国濒危动物红皮书·鱼类》[6]。目前,对长江刀鲚的资源动态[7-9]、种群结构[10-13]、生物学特征[13-16]和遗传进化分析[17-21]等进行了大量研究,而关于刀鲚繁殖生物学,仅见于陈文银[15]对长江刀鲚性腺发育的组织学研究;管卫兵[16]也对2006~2008年刀鲚溯河洄游期间刀鲚的生殖特征的研究;而对辽河双台子河口刀鲚个体繁殖力的研究尚未见报道。本研究通过观察刀鲚个体生殖力,探讨其与各生物学指标的相关关系,旨在为双台子河口刀鲚资源合理利用、种质资源保护和人工增殖提供参考。
1 材料与方法
1.1 双台子河口的自然概况
双台子河河口区位于121°30'~122°00'E,40° 50'~41°20'N,辽宁省盘锦市境内、辽东湾北部辽河下游冲积平原,地势低洼平坦,海拔高度为1.3~4.0m,坡降为1/20 000~1/25 000,多苇塘泡沼和潮间带滩涂[22]。水深多小于10m,水下地形是辽河等河口三角洲的水下延伸部分,地势自岸向海缓倾,水深逐渐增大。在河口附近海域,水道交错,浅滩广布,地形复杂多变[23]。本区属暖温带大陆性半湿润半干旱季风气侯。多年平均气温8.4℃,7月份最高气温为24.6℃,1、2月份极端最低值为-23.6~-29.3℃之间。年平均降水量为623.2mm,多年平均蒸发量为1 568.6mm[24]。
1.2 样本采集及处理
2015年4~7月在双台子河口(西河沿村段至河口水域)(图1),采用拉网、单层挂网(网长100m,网高1.5m,网目2cm、4cm、6cm)等方式捕获刀鲚579尾,选取5~6月采捕的性腺发育达Ⅳ期的176尾雌刀鲚用于生殖力研究。逐尾测量体长(L,mm)、体质量(W,g)、净体质量(Wo,g)、卵巢质量(W1,g)等生物学指标。用数显游标卡尺测量长度,精确至0.1mm,用电子天平测量重量,精确至0.1g。采用国标GB12763.6-91判定性腺分期。采用重量计数法统计个体绝对生殖力,即称重后,随机从卵巢前、中、后部称取约0.1g样品,用4%甲醛溶液固定,计数所有沉积卵黄的卵粒数[25]。采用左侧主鳃盖骨多条呈明暗(宽窄)相间排列的环纹来鉴定刀鲚的年龄。
1.3 数据分析
按下式计算相关指标:
个体绝对生殖力(粒/尾)F=(样品卵粒数/样品重)×W1

图1 双台子河口刀鲚采样点分布示意图Fig.1 Sampling sites of tapertail anchovy Coilia nasus in Shuangtaizi River esturary
体长相对生殖力(粒/mm)FL=F/L
体质量相对生殖力(粒/g)FW=F/W0
成熟系数(%)GSI=(W1/Wo)×100
肥满度(g/mm3)K=(W/L3)×100
式中:L为体长(mm),W为体质量(g),Wo为净体质量(g),W1为卵巢质量(g)。其他使用的符号及单位:年龄(A,龄),标本数(n,尾)
利用Excel 2010和SPSS 19.0进行数据统计和相关分析。利用线性函数、幂函数、指数函数、对数函数和二次函数方程等5种函数模型拟合个体生殖力与各生物学指标的关系,相关系数(R2)最大者作为选取型式,建立个体生殖力与生物学指标间的多元逐步回归方程。
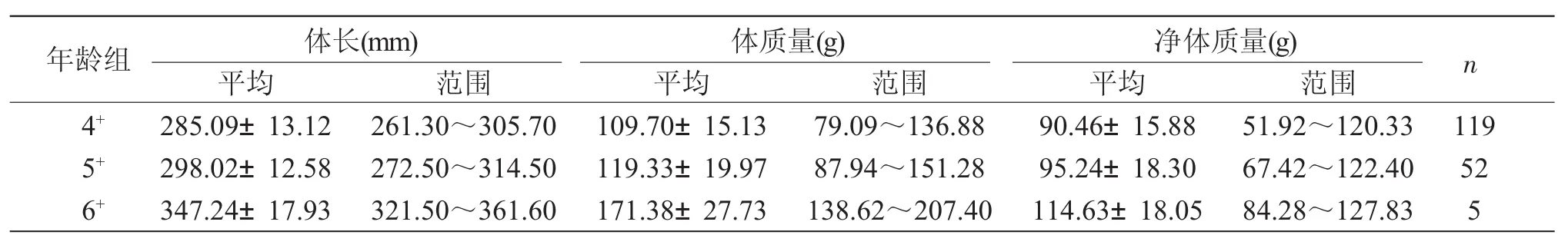
表1 刀鲚的相关生物学指标Tab.1 The related biological character indices in tapertail anchony Coilia nasus

表2 刀鲚的个体生殖力Tab.2 The individual fecundity(Mean±SD)in tapertail anchony Coilia nasus

图2 刀鲚生殖力分布Fig.2 The frequency of individual fecundity in tapertail anchony Coilia nasus
2 结果和分析
2.1 生物学指标
刀鲚的体长范围 261.30~361.60mm,平均294.73mm;体质量范围 79.09~207.40g,平均118.39g;净体质量范围51.92~127.83g,平均94.14g。
如表1所示,刀鲚繁殖群体由4+~6+龄3个年龄组成,4+龄组、5+龄组和6+龄组分别占67.61%、29.55%和2.84%,4+龄组最多。体长、净体质量和卵巢质量随年龄的增长而增大,同一年龄组个体间的体长、净体质量和卵巢重也存在差异。
2.2 个体生殖力
表2表明,刀鲚个体绝对生殖力(F)17 968~116 935粒,平均44 995粒;体长相对生殖力(FL)66~297粒/mm,平均为145粒/mm;体质量相对生殖力(Fw)192~710粒/g,平均为362粒/g;成熟系数(GSI)2.51%~51.07%,平均为20.43%。随着年龄的增长,F增大,而FL和FW在5+龄后有所下降。
由图2可知,F变幅在20 000~50 000粒之间的样本,占总数的72.6%;FL主要变幅在80~160粒/mm之间的,占样本总数68.6%;Fw主要变幅在200~350粒/g之间的,占样本总数72.0%。
2.3 个体生殖力与生物学指标的关系
2.3.1 个体生殖力与生物学指标的相关分析
由表3可知,F、FL与L、W、Wo、W1、GSI和A之间的相关系数均达到极显著水平(P<0.01),但F、FL与K之间的相关关系不显著。FW与W1和GSI之间的相关系数达到极显著水平(P<0.01),与L、W、Wo、K和A之间的相关关系不显著(P>0.05)。
2.3.2 个体生殖力与单一生物学指标的关系
从表4可见,F、FL与K,FW与L、W、Wo、K、A所拟合的5个方程均达不到显著水平。
F与L、GSI、A的拟合方程均以二次函数为最佳(表4),F与W、Wo的拟合方程以幂函数为最佳,F与W1的拟合方程则均以指数函数为最佳。FL与L、W、Wo的最佳拟合方程为幂函数,FL与W1、GSI、A;FW与W1、GSI的最佳拟合方程为二次函数。
2.3.3 个体生殖力与各项生物学指标的多元回归分析
将刀鲚的F、FL和FW分别与L、W、Wo、W1、GSI、K和A共7个生物学指标进行逐步多元回归分析,得到以下3个回归方程(R为相关系数,DW为残差独立性检验):
F=-15623.878+1158.377Wl+324.104W(n=176,R=0.851;DW=1.890,P<0.01);
FL=-68.850+3.868Wl+302687.076K(n=176,R=0. 728;DW=1.743,P<0.01);
FW=207.215-5.17L+7.5918GSI(n=176,R=0.646;DW=1.849,P<0.01)。
多元逐步回归显示,F与W和W1密切相关,FL与W1和K密切相关,FW与L和GSI密切相关。
2.4 卵径频率分布与产卵类型
刀鲚卵巢发育到第Ⅳ~Ⅴ期时,卵径变化在0.55~1.05mm之间,平均0.81mm。由图3可知,刀鲚的卵径频率分布图只有一个波峰,集中在0.75~0.95mm之间,所占比例为89.50%,由此推断刀鲚为一次性产卵鱼类。

表3 刀鲚个体生殖力与生物学指标的相关系数Tab.3 The correlation coefficients between individual fecundity and biological indicator indices in tapertail anchony Coilia nasus

表4 刀鲚个体生殖力与各生物学指标的最佳拟合方程Tab.4 The best fitting equations for individual fecundity and biological character indices in tapertail anchony Coilia nasus
3 讨论
3.1 刀鲚个体生殖力与繁殖策略
鱼类繁殖习性是对外界环境适应的一种进化过程,个体生殖力是种群生殖力的重要指标,受遗传特性与环境因子的共同影响[26,27]。本研究中刀鲚个体绝对生殖力17 968~116 935粒,平均44 995粒,低于蔡德霖[28]报道的黄河下游刀鲚、曹富康[29]报道的曹娥江刀鲚的个体绝对生殖力,略高于张敏莹[8]报道的长江下游刀鲚的研究结果。近年来,由于双台子河口水质污染改善不明显、上游给水量和输沙量减少及人为因素影响增大等原因,双台子河口刀鲚资源量大幅下降,为了种群的延续,被迫改变繁殖策略,增加产卵数量来抵御环境胁迫,保证种群的延续,个体绝对生殖力的上限也相应增大。
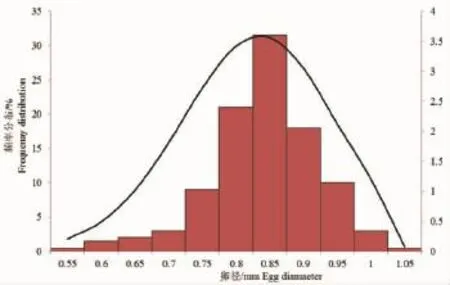
图3 刀鲚卵径的频率分布Fig.3 The frequency of egg diameter in tapertail anchony Coilia nasus
3.2 刀鲚个体生殖力的体征表达
鱼类个体生殖力不仅与本质特性、营养状况、外界环境条件有关,还与其生物学指标显著相关[30,31]。不同鱼类的个体生殖力与生物学指标之间的相关关系也有所不同。刀鲚F、FL与L、W、Wo、W1、GSI和A之间均呈极显著正相关(P<0.01),FW与仅与W1、GSI呈极显著正相关(P<0.01),F、FL、FW与K相关关系不显著(P>0.05)。这一结果与凤鲚Coilia mystus[32]、黄尾鲴Xenocypris davidi)[33]、黑斑原鮡Glyptosternum maculatum[34]的生殖力研究结果一致。
西藏高原鳅 Triplophysa tibetana[35]、黑斑狗鱼Esox reichrti[36]、广东北江马口鱼Opsariichthys bidens[37]、湖北道观河水库青梢鲌Culter dabryi[38]、东海银鲳Stromateoides argenteus[39]F与L呈二次函数关系;鸭绿江水系Hemiculter leucisculus[40]、徐家河水库翘嘴鲌Culter alburnus[41]、北江侧条光唇鱼Acrossocheilus parallens[42]、沱江宽体沙鳅 Botia reevesae[43]、湖北道观河水库青梢鲌、梁子湖乌鳢Channa argus[44]、切尾拟鲿Pseudobagrus truncatus、凤鲚[32]F与W、Wo均呈幂函数关系;黑斑原鮡[34]、凤鲚、切尾拟鲿F与W1、GSI均呈幂函数关系;鸭绿江水系黑斑狗鱼、广东北江马口鱼F与GSI呈二次函数关系。本研究中F与W、Wo均呈幂函数相关,F与L、GSI、A及FW与W1、GSI均呈二次函数相关,F与W1及FL与L、W、Wo均呈指数函数相关。
本研究中,单项回归分析结果与多元逐步回归分析的结果不同,如单项回归分析中K与F、FL、FW无显著相关性,而在多元逐步回归分析中,K与FL呈显著相关。多元逐步回归显示,F与W和W1密切相关,FL与W1和K密切相关,FW与L和GSI密切相关。可能是由于各项生物指标之间的关系较为复杂,单一指标可能与个体生殖力不相关,但在多元逐步回归分析中可能由于其他指标的关系而呈现出一定的相关性。
3.3 刀鲚产卵环境条件及产卵类型
洄游型刀鲚受精卵具有油球,漂浮于水体上层孵化。产卵场一般水流较急,无污染,水深为3~5m,pH为7,水温为15~27.5℃[8,45]。
在鱼类繁殖生物学中,常用卵径频率分布来判断鱼类的产卵类型。刀鲚的卵径频率分布图上只有一个波峰,所以认为刀鲚为一次性产卵鱼类,这与同属的凤鲚[32]和短颌鲚[46]的研究结果一致,同样与人工养殖条件下刀鲚繁殖力的研究结果相一致[47]。
[1]袁传宓,林金榜,秦安舲,等.关于我国鲚属鱼类分类的历史和现状—兼谈改造旧鱼类分类学的几点体会[J].南京大学学报:自然科学版,1976(2):1-12.
[2]袁传宓,秦安舲,刘仁华,等.关于长江中下游及东南沿海各省的鲚属鱼类种下分类的探讨[J].南京大学学报:自然科学版,1980(3):67-77.
[3]袁传宓,秦安舲.我国近海鲚鱼生态习性及其产量变动状况[J].海洋科学,1984(5):35-37.
[4]刘蝉馨,秦克静.辽宁动物志·鱼类[D].沈阳:辽宁科学技术出版杜,1987.
[5]张世义.中国动物志(硬骨鱼纲):鲟形目,海鲢目,鲱形目,鼠鳝目[D].北京:科学出版社,2001:148-156.
[6]乐佩琦,陈宜瑜.中国濒危动物红皮书·鱼类[D].北京:科学出版社,1998.
[7]长江流域刀鲚资源调查协作组.长江流域刀鲚资源调查报告[R].1977:1-179.
[8]张敏莹,徐东坡,刘凯,等.长江下游刀鲚生物学及最大持续产量研究[J].长江流域资源与环境,2005,14(6): 694-698.
[9]毛成责,矫新明,钟俊生,等.长江口刀鲚资源现状及保护研究进展[J].淮海工学院学报:自然科学版,2015,24(3):78-83.
[10]袁传宓,林金榜,刘仁华,等.刀鲚的年龄和生长[J].水生生物学集刊,1978,6(3):285-298.
[11]万全,赖年悦,李飞,等.安徽无为长江段刀鲚生殖洄游群体年龄结构的变化分析[J].水生态学杂志,2009,2(4):60-65.
[12]郑飞,郭弘艺,唐文乔,等.溯河洄游的长江刀鲚种群的年龄结构及其生长特征[J].动物学杂志,2012,47(5): 24-31.
[13]董文霞,唐文乔,王磊.长江刀鲚繁殖群体的生长特性[J].上海海洋大学学报,2014,23(5):669-674.
[14]何为,李家乐,江芝娟.长江刀鲚性腺的细胞学观察[J].上海水产大学学报,2006,15(3):292-296.
[15]陈文银,李家乐,练青平.长江刀鲚性腺发育的组织学研究[J].水产学报,2006,30(6):773-777.
[16]管卫兵,陈辉辉,丁华腾,等.长江口刀鲚洄游群体生殖特征和条件状况研究[J].海洋渔业,2010,32(1):73-81.
[17]刘文斌.中国鲚属4种鱼的生化和形态比较及其系统发育的研究[J].海洋与湖沼,1995,26(5):558-565.
[18]马春艳.鳗科鱼类分子系统进化及凤鲚刀鲚遗传多样性研究[D].上海:华东师范大学,2010.
[19]周晓犊.中国鲚属鱼类物种有效性及刀鲚种群遗传结构研究[D].上海:上海海洋大学,2011.
[20]杨巧莉.中国鲚属鱼类进化关系及刀鲚、凤鲚的分子系统地理学研究[D].青岛:中国海洋大学,2012.
[21]陈芳.刀鲚和凤鲚遗传多样性的微卫星分析[D].上海:上海海洋大学,2012.
[22]刘爱江,吴建政,姜胜辉等.双台子河口区悬沙分布和运移特征[J].海洋地质动态,2009,25(8):13-16.
[23]朱龙海.双台子河口潮流沉积体系研究[D].青岛:中国海洋大学,2004.
[24]杨俊鹏.双台子河口潮滩土壤元素地球化学特征及其生态效应[D].长春:吉林大学,2007.
[25]凌建忠,严利平,林龙山,等.东海带鱼繁殖力及其资源的合理利用[J].中国水产科学,2005,12(6):726-730.
[26]Alvarez-Lajonchere L.The fecundity of mullet(Pisces, Mugilidae)from Cuban waters[J].Journal of Fish Biology,1982,21(6):607-613.
[27]苏锦祥.鱼类学与海水鱼类养殖[M].北京:中国农业出版社.1995.
[28]蔡德霖,汲岱昌,周才武.黄河下游刀鲚的生态和渔业概况[J].山东大学学报:自然科学版,1980(2):97-107.
[29]曹富康.曹娥江刀鲚生物学[J].动物学杂志,1985(6): 20-25.
[30]殷名称.鱼类生态学[D].北京:中国农业出版社,1995.
[31]Anibeze CI P and InyangN M.Oocyte structure,fecundityand sex ratio of Heterobranchus longifilis Valenciennes 1840 in IdodoRiver basin(Nigeria)with comments on the breeding biology[J].Journal of Aquatic Sciences,2000, 15(1):59-62.
[32]毕雪娟,张涛,冯广朋,等.长江口凤鲚个体生殖力的研究[J].海洋渔业,2015,37(3):223-232.
[33]蓝昭军,李强,陈龙秀,等.北江黄尾鲴个体生殖力的研究[J].华南师范大学学报:自然科学版,2008(4):107-113.
[34]李红敬,刘鸿艳,樊启学,等.黑斑原鮡个体生殖力研究[J].应用与环境生物报,2008,14(4):499-502.
[35]刘鸿艳,谢从新,郑跃平,等.西藏高原鳅个体繁殖力的研究[J].淡水渔业,2009,39(4):12-16.
[36]霍堂斌,袁美云,阿达可白克·可尔江,等.白斑狗鱼与黑斑狗鱼繁殖力的比较研究[J].大连海洋大学学报, 2010,25(6):511-517.
[37]李强,蓝昭军,赵俊,等.广东北江马口鱼个体生殖力研究[J].四川动物,2010,29(3):440-445.
[38]王银东,熊邦喜,马徐发.湖北道观河水库青梢鲌(Culter dabryi)的个体生殖力[J].海洋与湖沼,2007,38(2): 180-186.
[39]倪海儿,龚启祥.东海银鲳个体生殖力的研究[J].浙江水产学院学报,1995,14(2):118-122.
[41]覃亮,熊邦喜,吕光俊.徐家河水库翘嘴鲌的个体生殖力[J].应用生态学报,2009,20(8):1952-1957.
[42]蓝昭军,赵俊,李强.北江侧条光唇鱼的个体生殖力[J].华南师范大学学报:自然科学版,2010(1):92-97.
[43]黄燕,岳兴建,王芳,等.沱江宽体沙鳅个体生殖力的研究[J].四川动物,2011,30(6):916-920.
[44]马陶武,谢从新.梁子湖乌鳢个体生殖力研究[J].水利渔业,2000,20(5):1-3.
[45]陈栋良.长江刀鲚的天然繁殖与胚胎发育观察[J].水产科技情报,1992,19(2):49-51.
[46]刘引兰.鄱阳湖短颌鲚的鉴定及其生物学特性[D].南昌:南昌大学,2008.
[47]徐钢春,万金娟,顾若波,等.池塘养殖刀鲚卵巢发育的形态及组织学研究[J].中国水产科学,2011,18(3):537-546.
Individual Fecundity of Tapertail Anchovy Coilia nasus from Shuangtaizi Estuary
HUO Tang-bin1,ZHENG Wen-jun2,LI Geng2,ZHAO Rong-wei3,DAI Shang-ying4,NA Wen-xue4
(1.Key Field Scientific Observation Station of Fisheries Resources and Environment in Heilongjiang River Valley,Ministry of Agriculture,Heilongjiang River Fisheries Research Institute,Chinese Academy of Fishery Sciences,Harbin 150070,China; 2.Yingkou Enhancement and Experiment Station,Chinese Academy of Fishery Sciences,Yingkou 115004,China; 3.Harbin Fisheries Technology Extension Center,Harbin 150076,China; 4.Northeast Agricultural University,Harbin 150040,China)
Biological parameters including body length(L),body weight(W),net body weight(W0)and ovary weight(W1)were measured 176 individuals of females tapertail anchovy Coilia nasus with stageⅣovary collected from the Shuangtaizi estuary in Liao River from May to June in 2015.The left opercula bone was used to estimate age,egg diameter and account were measured by a caliper,and the individual absolute fecundity(F),relative fecundity to body length(FL)and relative fecundity to body weight(FW)were calculated by gravimetric method to evaluate the relationship between individual fecundity and the biological indicators by five kinds of mathematical model and stepwise regression.The results showed that the individual absolute fecundity(F)was varied from 17 968 eggs to 116 935 with an average of 44 995 eggs,the individual relative fecundity per millimeter(FL)ranged from 66 to 297 eggs with an average of 145 eggs per millimeter and the individual relative fecundity per gram(FW)from 192 to 710 eggs with an average of 362 eggs per gram age in all samples of the fish from 4+to 6+years old.The relationship between individual absolute fecundity(F)and relative fecundity to body length(FL)was significantly positive to body length(L),body weight(W),net body weight(W0),ovary weight(W1),Gonado-Somatic Index(GSI),age,excepting for condition factor(K).While the relationship between FWwas only significantly positive correlated with W1and GSI,but not correlated with other morphometrical indicators.The stepwise multi-regressive analysis revealed that the relationship between F and W,and W1was significantly correlated,with the regression equation of F=-15623.878+1158.377Wl+324.104W (n=176,R=0.851;DW=1.890,P<0.01).The significant regression equation between FLwith W1and K,was expressed as FL=-68.850+3.868Wl+302687.076K(n=176,R=0.728;DW=1.743,P<0.01);the significant regression equation between FW,L and GSI was described as FW=207.215-5.17L+7.5918GSI(n=176,R=0.646;DW=1.849,P<0.01).There was an egg with diameter from 0.55 mm to 1.05 mm with an average of 0.81 mm in the stageⅣ~Ⅴovarres,the profile of egg diameter indicating that tapertail anchovy spawns once a year.
Coilia nasus;individual fecundity;Shuangtaizi estuary
Q492
A
1005-3832(2016)06-0031-06
2016-06-08
中国水产科学研究院基本科研业务费(2014C02XK01);大辽河口鲚属鱼类早期资源调查研究(2014C02XK01).
霍堂斌(1980-),男,博士,助理研究员,从事鱼类生态学研究.E-mail:tbhuo@163.com
